
不同储存年限老芒麦种子种带真菌检测及致病性测定
2015-02-25 08:34陈焘南志标
草业学报 2015年2期
陈焘,南志标
(草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020)
不同储存年限老芒麦种子种带真菌检测及致病性测定
陈焘,南志标*
(草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020)
摘要:对来自青海的5个不同收获年份的老芒麦种样进行了系统的种带真菌研究;测定了12种分离率大于1%的种带真菌对老芒麦种子萌发和幼苗生长的影响。结果表明,种样发芽率为56%~80%,S2发芽率最高,达到80%,显著高于S1和S5(P<0.05);种样带菌率为24%~38%,随储藏时间延长呈下降趋势,S5带菌率最高,达到38%,显著高于其他种样(P<0.05);共鉴定出老芒麦种带真菌15属17种,真菌分离率为0.25%~8.75%,其中青霉和曲霉是老芒麦最常见的种带真菌,在5个种样上均被分离得到;燕麦镰孢、串珠镰孢、镰孢菌1、离蠕孢和德氏霉5种真菌是老芒麦最主要的致病真菌,均显著地降低了老芒麦种子的萌发、抑制了幼苗的生长、降低了幼苗的生物量(P<0.05);细交链孢对种子的萌发没有显著抑制作用(P>0.05),但是显著地降低了幼苗的长度和干物质产量(P<0.05)。皮思霉、离蠕孢、曲霉3种真菌显著延长了老芒麦种子平均萌发时间,而燕麦镰孢则显著地缩短了种子平均发芽时间(P<0.05)。
关键词:老芒麦;发芽率;种带真菌;致病性
老芒麦(Elymussibiricus)是禾本科(Gramineae)小麦族(Triticeae)披碱草属(Elymus)多年生疏丛型中旱生植物,广泛分布于北半球温带地区,主要集中在东欧、西伯利亚、中国、蒙古、朝鲜、日本及印度等;在我国主要分布在东北、华北、西北和青藏高原等地。老芒麦具有抗寒性强、适应性广、品质好、草籽产量高等优点,是我国青藏高原地区广泛栽培的优良牧草,常用于牦牛和藏羊等家畜的补饲饲料,在草地畜牧业中发挥着重要作用,同时对退化草地治理、三江源区生态环境保护等具有重要意义。
种子作为高等植物的繁殖器官,能够携带大量危害种苗或植株的病原,既是病害的载体,同时也是受害者。而在所有病原中,真菌是对种子质量影响最为严重的一类。种带真菌既可通过混杂于种子中间或粘附于种子表面,也可侵入种子组织内部,导致病害在时间上进行延续(从一个生长季到下一个生长季)和空间上进行扩展(从此地到彼地)。种带病原真菌不仅对种子的萌发、幼苗的健康生长产生负面影响,而且能在贮藏期间降低种子的品质[10]。因此,对种子进行病原真菌及其携带量检测是种子健康检验的重要组成部分[11]。
目前,国内外学者对多种植物种子进行了种带真菌研究[12-14]。例如,Wiewióra和Prończuk[15]对多年生黑麦草(Loliumperenne)进行了种带真菌检测,分离出细交链孢(Alternariaalternata)、镰刀菌属(Fusariumsp.)、根腐离蠕孢(Bipolarissorokuniana)、德氏霉属(Drechsleraspp.)、黑色附球菌(Epicoccumnigrum)、立枯丝核菌(Rhizoctoniasolani)、枝孢属(Cladosporiumsp.)等多种病原真菌。又如,李春杰和南志标[13]对来自新疆等11个省区的38个苜蓿(Medicago)种样进行了种带真菌检测,共鉴定出36属40种真菌,其中青霉(Pecinilliumsp.)、细交链孢、粉红单端孢(Trichotheciumroseum)、黑曲霉(Aspergillusniger)、多主枝孢(Cladosporiumherbarum)和黄曲霉(Aspergillusflavus)为苜蓿常见的种带真菌, 且对种子或幼苗造成不同程度的危害。然而,关于老芒麦种带真菌的研究几乎属于空白,仅见袁庆华等[16]对产自甘肃甘南的老芒麦种子进行了初步检测。
近年来,随着退耕还草、草原改良和人工草地建设等工程的实施,老芒麦的种植面积不断扩大,种子需求量迅速增加[17]。因此,本研究主要通过对我国青藏高原地区的5个不同收获年份的老芒麦种子进行种带真菌检测及分离,明确不同储存年限的老芒麦种带真菌区系,测定种带真菌对种子萌发以及幼苗生长的影响,确定老芒麦主要致病真菌,从而为老芒麦种传病害的防治提供理论依据。
1材料与方法
1.1 研究材料
供试老芒麦种子分别于2006,2007,2008,2009,2010年收获于青海,4℃保存于农业部牧草与草坪草种检中心种子库(兰州)。种样编号依次为S1,S2,S3,S4,S5。试验于2010年11月至2011年5月进行。
1.2 研究方法
1.2.1种子发芽率的测定采用培养皿纸上法(TP),每种样随机取400粒净种子,均匀摆放于铺有两层滤纸直径120 mm培养皿中,每皿100粒,4次重复,25℃恒温培养15 d,每天加水保湿。参照《国际种子检验规程》[18],第5天初次统计发芽种子数,第15天发芽结束,计算种子发芽率。
1.2.2种带真菌分离与鉴定参照李春杰和南志标[13]的方法,每种样随机取老芒麦种子200粒,经75%酒精浸泡1 min,用1% NaClO消毒2 min,用无菌蒸馏水充分洗涤5次,放置于无菌滤纸上进行干燥,然后用经火焰消毒的镊子随机选取10粒,等距离放置于ABPDA培养基上(内加100 mg/L链霉素和100 mg/L青霉素的马铃薯葡萄糖琼脂培养基),4皿为一重复,4次重复,置于20℃恒温培养箱中培养。第3天开始观察菌落生长情况,及时挑取长出菌落至PDA培养基上纯化培养,纯化好的菌株置试管于4℃冰箱中保存,备用。待不再有新的菌落出现时统计带菌种子数和带某种真菌种子数,参照徐秀兰等[19]的方法,计算种子带菌率和真菌分离率:
种子带菌率(%)=(带菌种子数/检测种子总数)×100
某菌分离率(%)=(带某类菌种子数/检测种子总数)×100
参考相关著作[20-21],借助显微镜观测孢子形态、孢子着生方式、测定孢子大小、孢子梗长度等形态学指标进行鉴定。
1.2.3种带真菌致病性测定选取分离率大于1%的12种真菌分离物用于试验。供试真菌接种于PDA培养基上在黑暗培养箱中培养2周后,打开皿盖加入10 mL无菌水,用灭菌的解剖刀将孢子轻轻刮下,充分分散后配成孢子悬浮液,调整孢子浓度至1×106/mL,备用。
参照杨颜霞等[22]的方法,随机选取400粒种子,在无菌条件下用1% NaClO消毒10 min,无菌蒸馏水充分润洗5次,置于经灭菌的滤纸上干燥,用备好的孢子悬浮液2 mL接种24 h,对照用2 mL无菌蒸馏水代替。将接种过真菌孢子悬浮液的种子等距离放置于铺有两层无菌滤纸的灭菌培养皿(直径12 cm)中,每皿100粒,重复4次,于25℃恒温的光照培养箱中,12 h光照/12 h黑暗培养。每天记录发芽种子数,15 d后按《国际种子检验规程》统计种子最终发芽数,分别计算种子最终发芽率(FGP)、发芽势(GP)、平均发芽时间(MGT)。另外,每皿随机取15株种苗测定芽长和根长,然后置于80℃烘箱中24 h,测定干重。
1.3 统计分析
用Microsoft Excel录入数据并制表。采用SPSS 15.0进行方差分析和多重比较(Duncan法)。发芽率及发芽势数据分析前反正弦转换。
2结果与分析
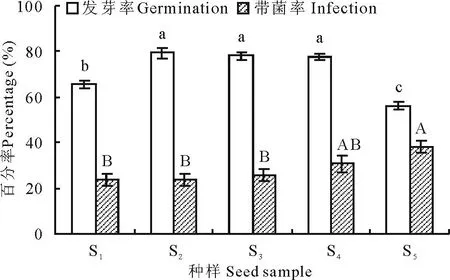
图1 不同收获年份种样发芽率和带菌率Fig.1 Percentage of seed germination and seed-borne fungus infection among five seed samples of E. sibiricus 不同大写和小写字母分别表示不同种样发芽率和带菌率差异显著(P<0.05,Duncan)。Different capital and small letters mean the significant difference of seed germination percentage and seed infection rate among five samples respectively at P<0.05 (Duncan).
2.1 种样发芽率与带菌率
从图1看出,不同收获年份的老芒麦发芽率不同。总体来看,种样发芽率介于56%~80%,其中种样S2发芽率最高,达到80%,显著高于S1和S5(P<0.05);种样S2、S3与S4之间发芽率差异不显著(P>0.05)。老芒麦种样带菌率随着储藏时间的增加呈下降趋势,带菌率介于24%~38%。其中,种样S5带菌率最高,达到38%,显著高于其他种样(P<0.05);种样S1、S2与S3之间带菌率差异不显著(P>0.05)。
2.2 种带真菌种类及分离率
经分离培养,共鉴定出老芒麦种带真菌15属17种(表1),另有3种真菌分离物未能得到鉴定。各种真菌的平均分离率介于0.25%~8.75%,平均为1.61%。其中,分离率大于1%的有12种,按分离率由高到低依次为青霉(8.75%)、曲霉(4.50%)、细交链孢(3.75%)、丛梗孢(2.75%)、串珠镰孢(2.63%)、德氏霉(2.00%)、镰孢菌1(1.38%)、皮思霉(1.25%)、单格孢(1.25%)、燕麦镰孢(1.13%)、离蠕孢(1.13%)、链二孢(1.13%)。在5个种样中均分离到的真菌有2种,分别是青霉和曲霉(表1)。
2.3 种带真菌对老芒麦种子发芽率的影响
与对照相比,供试的12种种带真菌对老芒麦种子的发芽率产生了不同程度的影响(图2)。根据其对种子发芽率的影响程度,可归为3类: 1)燕麦镰孢、镰孢菌1和串珠镰孢3种镰孢菌极显著地降低了种子的发芽率,分别达到14.5%,26.5%和31.3%(P<0.01);2)离蠕孢、德氏霉、单格孢、链二孢、皮思霉5种真菌较强地抑制了老芒麦种子的萌发(P<0.05);3)曲霉、细交链孢、青霉、丛梗孢4种真菌对种子的发芽率没有明显作用(P>0.05)。
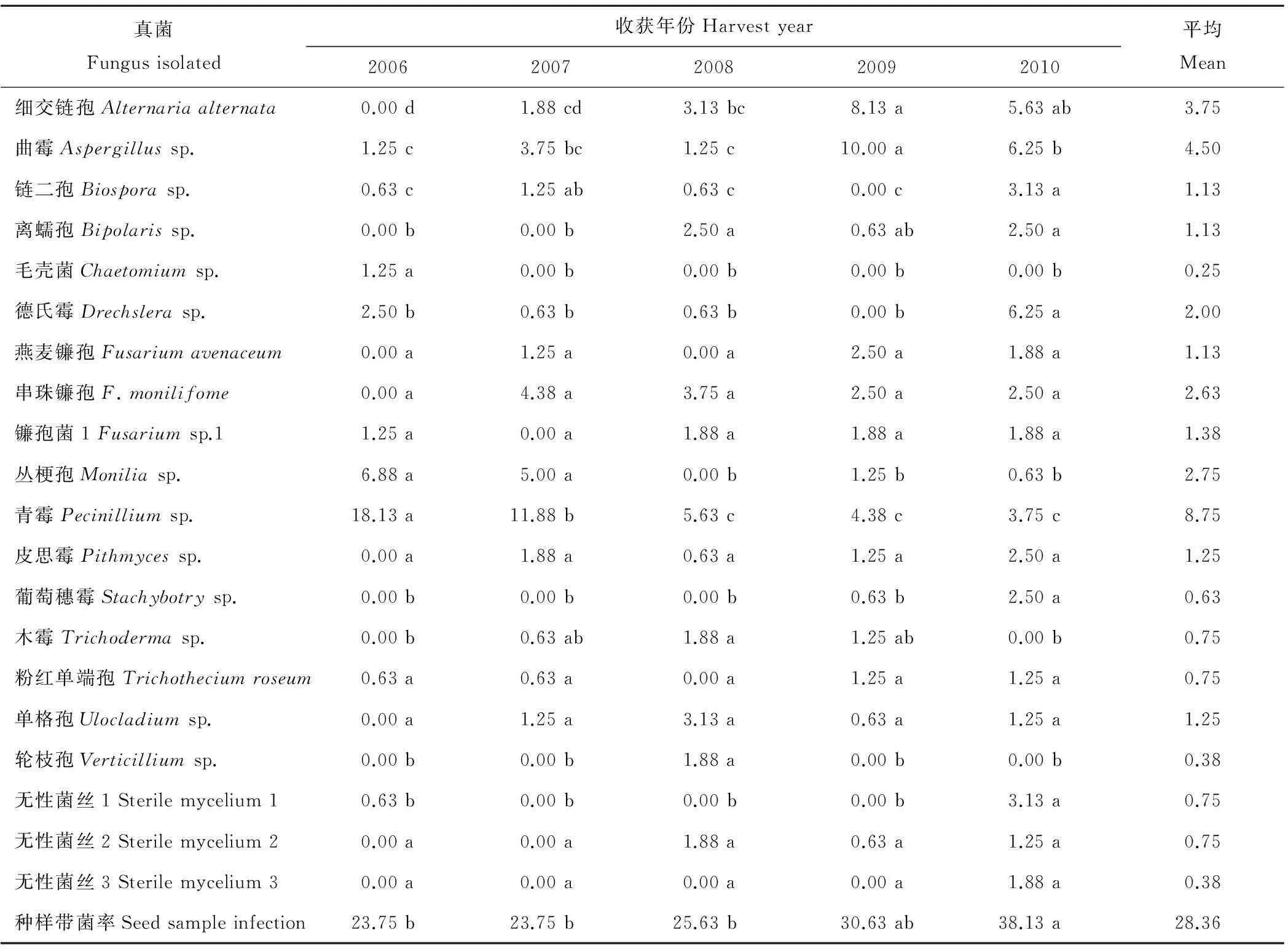
表1 老芒麦种样种带真菌种类及分离率
注:同行数据后不同小写字母表示差异显著(P<0.05,Duncan)。
Note:Different small letters indicate the significant difference among five samples atP<0.05 (Duncan).
2.4 种带真菌对老芒麦种子发芽势的影响
发芽势是反映种子发芽整齐度的指标,其数值越大,表示种子萌发越集中,反之则越分散。从结果可以看出(图3),不同种带真菌对老芒麦种子的发芽势影响不同。燕麦镰孢、串珠镰孢和镰孢菌1等3种真菌极显著地降低了种子的发芽势(P<0.01);离蠕孢、德氏霉、单格孢、链二孢、皮思霉、青霉、细交链孢、丛梗孢8种真菌显著地降低了老芒麦种子的发芽势(P<0.05);曲霉对种子发芽势没有显著抑制作用(P>0.05)。
2.5 种带真菌对老芒麦种子平均发芽时间的影响
活力较高的种子具有发芽整齐、出苗快的特点,反之则差异性明显。从结果可以看出(图4),不同真菌对老芒麦种子的平均发芽时间影响不同。皮思霉、离蠕孢、曲霉3种真菌显著延长了老芒麦种子平均萌发时间(P<0.05);相反,燕麦镰孢显著地缩短了种子平均发芽时间(P<0.05)。其他真菌对老芒麦种子的平均发芽时间没有显著影响(P>0.05)。
2.6 种带真菌对老芒麦幼苗生长的影响
从图5看出,接种真菌对老芒麦幼苗的生长产生了不同程度的影响。与对照相比,供试的12种种带真菌均显著地降低了老芒麦芽长(P<0.05)。3种镰孢菌(燕麦镰孢、串珠镰孢和尖镰孢)对苗长的抑制作用最为明显,接种后老芒麦苗长分别只有0.51,0.62和1.14 cm;其次细交链孢、离蠕孢、丛梗孢、链二孢和德氏霉5种真菌较强地抑制了幼苗的生长;曲霉、青霉、皮思霉、单格孢对幼苗生长的影响较弱。除曲霉对老芒麦根长无显著影响外,其他真菌均显著地抑制了老芒麦幼根的生长(P<0.05)。燕麦镰孢和串珠镰孢对根长的抑制作用最为显著,接种后根长分别为0.53和0.57 cm;尖镰孢、细交链孢、离蠕孢、链二孢、单格孢对老芒麦幼苗的根长也有较强的抑制作用;德氏霉、青霉、丛梗孢、皮思霉4种真菌对老芒麦根长的影响较弱。
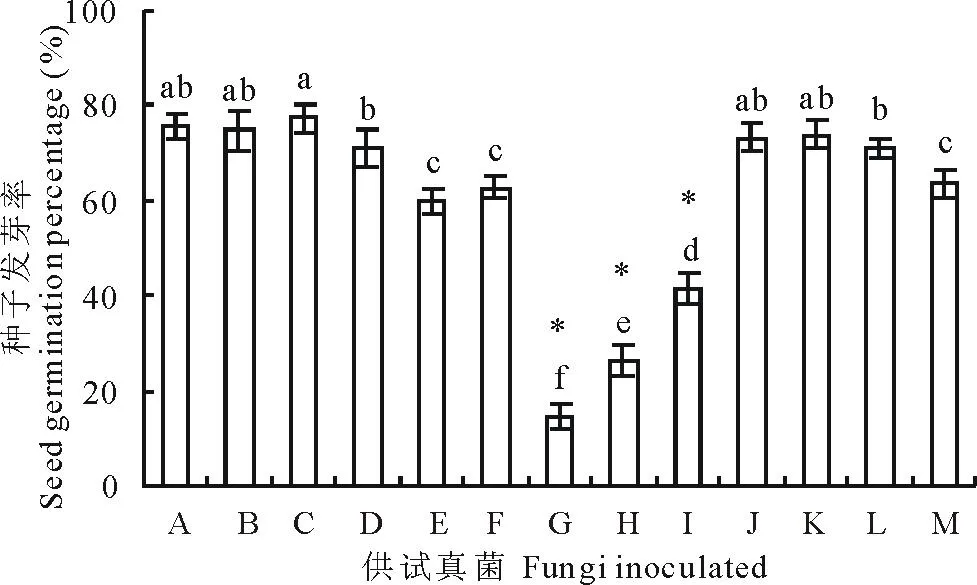
图2 真菌对老芒麦种子发芽率的影响Fig.2 Percentage of seed germination of E. sibiricus after inoculation with various seed-borne fungi A:对照Control; B:细交链孢Alternaria alternata; C:曲霉Aspergillus sp.; D:链二孢Biospora sp.; E:离蠕孢Bipolaris sp.; F:德氏霉Drechslera sp.; G:燕麦镰孢Fusarium avenaceum; H:串珠镰孢F. monilifome; I:镰孢菌1 Fusarium sp.1; G:丛梗孢Monilia sp.; K:青霉Pecinillium sp.; L:皮思霉Pithmyces sp.; M:单格孢Ulocladium sp.;不同小写字母表示在P<0.05水平差异显著,*表示在P<0.01水平差异显著Different letters and * indicate significant difference at 0.05 level and 0.01 level with Duncan’s multiple range tests, respectively.下同The same below.
2.7 种带真菌对老芒麦幼苗干重的影响
由图6可以看出,除曲霉外,其他供试种带真菌均不同程度地降低了老芒麦幼苗的干重。其中,燕麦镰孢、串珠镰孢、镰孢菌1、细交链孢极显著地降低了幼苗的干重(P<0.01);链二孢、离蠕孢、德氏霉、丛梗孢、青霉、皮思霉、单格孢7种真菌显著降低了老芒麦幼苗的干重(P<0.05)。
3讨论
种子休眠是导致种子发芽率降低的重要原因,随着储存时间的增加,种子的休眠会逐渐被解除[23]。本试验所试的5种老芒麦种样中,种样S5储藏时间最短, 发芽率最低,显著低于其他种样,可能是由于种子收获时间不长,大多数种子还处于种子休眠状态所致[24-25]。另外,在自然状态下,随着储藏年限的增加,种子活力不断下降,发芽率也会逐渐下降[26]。在本试验中,种样 S2、S3和S4发芽率差异不显著,表明随着储藏年限增加种子发芽率并未显著降低,可能是由于老芒麦种子在低温条件下能持久保持活力,种子的发芽率还未呈现明显的下降趋势[27]。与之相比,种样带菌率随着储藏年限呈现下降的趋势,这与李春杰等[10]在紫花苜蓿的研究结果类似。究其原因,一方面可能是由于刚成熟的种子携带着大量的真菌(既通过母体植株垂直传播而来,也通过周围环境水平传播获得)。随着储藏年限的增加,种子种带真菌区系不断发生变化,一些不能适应储藏环境的种带真菌逐渐被淘汰。另一方面,Meyer等[28]研究表明真菌侵染速率与种子的萌发速率呈负相关。刚收获的种子由于大部分处于休眠状态不能萌发,给真菌的大量繁殖创造了条件,因此更容易被分离。而随着储藏时间的延长,老芒麦种子休眠不断被解除,种子通过快速萌发可以规避真菌的侵染[29]。
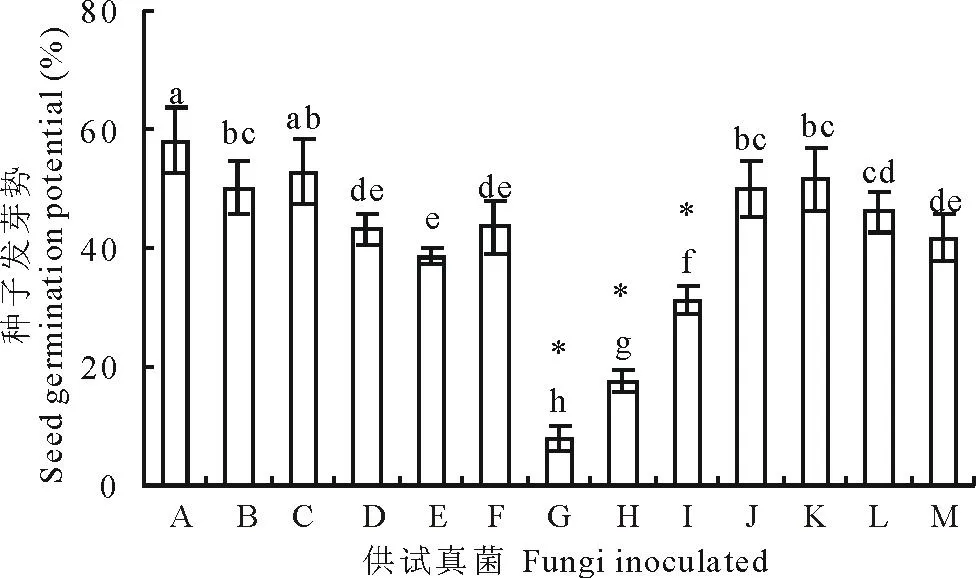
图3 真菌对老芒麦种子发芽势的影响Fig.3 Seed germination potential of E. sibiricus after inoculation with various seed-borne fungi
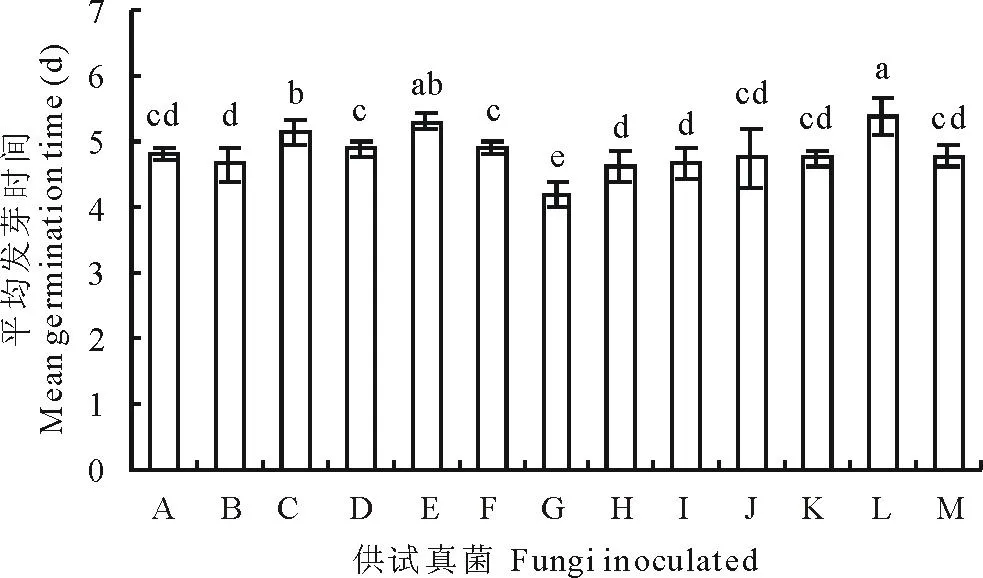
图4 真菌对老芒麦种子平均发芽时间的影响Fig.4 Seed mean germination time of E. sibiricus after inoculation with various seed-borne fungi
通过分离培养,本试验从5个老芒麦种样中共检测出15属17种种带真菌,另有3种白色丝状真菌未产孢,未能鉴定(表1)。其中青霉和曲霉两种真菌是老芒麦种子最常见的种带真菌,在5个种样中均被分离得到,这与李春杰和南志标[13]在苜蓿种子上的研究结果相类似。在对分离率高于1%的12种种带真菌进行致病性研究后发现,燕麦镰孢、串珠镰孢、镰孢菌1、离蠕孢和德氏霉5种真菌是老芒麦最主要的致病真菌,均显著地降低了老芒麦种子的萌发、抑制了幼苗的生长、降低了幼苗的生物量。细交链孢尽管对种子的萌发没有显著抑制作用(图2),但是对幼苗的生长产生了较强的抑制作用,显著降低了老芒麦苗长和根长,以及干物质产量(图5和图6),这与杨颜霞等[22]在结缕草(Zoysiajaponica)种子上研究结果相一致。
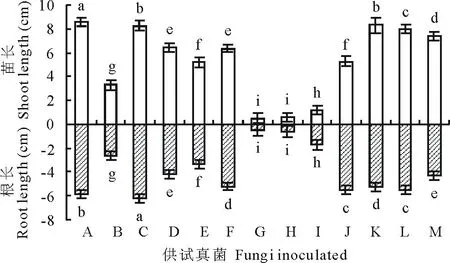
图5 真菌对老芒麦幼苗生长的影响Fig.5 Shoot and root length of E. sibiricus after inoculation with various seed-borne fungi
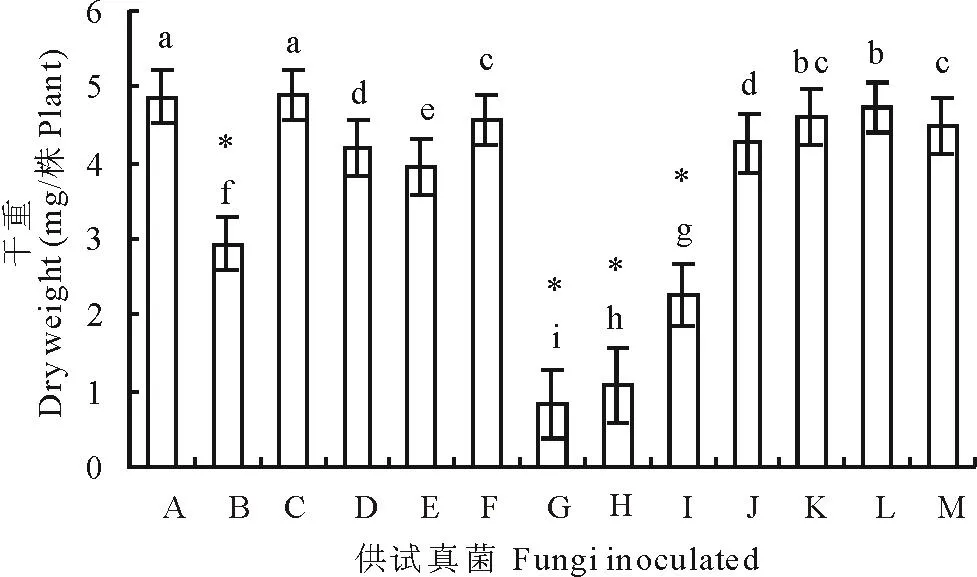
图6 真菌对老芒麦幼苗干重的影响Fig.6 Seedling dry weight of E. sibiricus after inoculation with various seed-borne fungi
种子平均发芽时间是其固有的一种特性,但是在受到不同程度的胁迫后可能被延长或者缩短。如阮松林和薛庆中[30]研究发现高盐胁迫可延缓水稻(Oryzasativa)种子萌发时间,而颜宏等[31]研究发现不同盐溶液处理缩短了向日葵(Helianthusannuus)种子的萌发时间。究其原因,可能是不同植物种子对盐胁迫的响应机制不同,需要更深入的研究。在本研究中,种带真菌对老芒麦种子的萌发时间表现不一,这与杨颜霞等[22]在结缕草种子上的研究结果相类似。
种带真菌在种子中的存在部位是病害发生的先决条件,几乎决定了病害发生的强度[32]。陈林[33]在红豆草(Onobrychisviciaefolia)种子外部组织(荚皮和种皮)中检测到27种真菌,而在内部组织(胚和胚乳)中仅检测出10种真菌。然而,张颖[34]研究发现,多年生黑麦草(Loliumperenne)种带真菌主要存在于胚乳,其次为稃壳,甚少存在于胚中。究其原因,一方面可能是由于不同物种间病原真菌侵染种子的能力不同,即种子形态差异对真菌侵染阻力不一,另一方面气候条件等因素差异也会影响病原真菌对种子的侵染[35]。本研究只对带有稃片的老芒麦种子进行了种带真菌检测,关于其种带真菌的存在部位有必要进行更深入的研究。
温度和湿度对种带病害的发生和发展有着极为重要的影响[10]。因此,老芒麦种子应选择在天气晴朗时快速收获,及时晒干,且储藏时注意通风透气,从而降低病原真菌对种子的侵染。另外,种子播种时用杀菌剂拌种可防止病害在田间传播与流行[36],从而改进草地建植,提高草地的生产力[37]。甲基托布津是一种广谱内吸性杀菌剂,对细交链孢等多种种带病原真菌有较好的抑制效果[38]。例如,南志标[14]对沙打旺(Astragalusadsurgens)种带真菌的研究发现,甲基托布津处理种子后细交链孢的检出率降低了46.1%。Buchenauer[39]研究发现三唑酮对串珠镰孢有很好的抑制效果。而且,Papavizas和Lewis[40]研究发现两种或多种杀菌剂混合使用效果优于单一杀菌剂。本研究发现镰孢菌对老芒麦种子萌发和幼苗生长的影响最为严重,而细交链孢也显著地降低了幼苗的生物量。因此,采用甲基托布津和三唑酮混合拌种也许能降低老芒麦种带病害的发生,然而其能否有效抑制老芒麦种带病害则需要更深入的研究。
Reference:
[1]Yan J J, Bai S Q, Chang D,etal. Genetic diversity of wildElymussibiricusL. germplasm from Qinghai-Tibetan Plateau in China detected by SSR Markers. Chinese Agricultural Science Bulletin, 2010, 26(9): 26-33.
[2]Dong J S, Zhang S Z. Study ofElymussibiricusL. Chinese Journal of Grassland, 1980, 2: 29-33.
[3]Chen G, He L F. Evaluation of ecological adaptability and productivity of two species ofElymusin alpine region. Pratacultural Science, 2004, 21(9):39-42.
[4]Yu Z, Sun Q Z, Deng B,etal. Studies on making Siberian Wildryegrass silage. Journal of Agricultural Science and Technology, 2008, 10(1): 98-102.
[5]Gu X Y, Guo Z H, Zhang X Q,etal. Genetic diversity ofElymussibiricusgermplasm resources revealed by SRAP makers. Acta Prataculturae Sinica, 2014, 23(1): 205-216.
[6]Dong S K, Hu Z Z, Long R J,etal. Community characteristics of mixed grassland with perennial grass in Alpine Region of Tibetan Plateau. Chinese Journal of Ecology, 2003, 22(5): 20-25.
[7]Sinclair J B. The seed: a microcosm of microbes. Journal of Seed Technology, 1979, 4(2): 68-73.
[8]Mirocha C, Pathre S, Schauerhamer B,etal. Natural occurrence ofFusariumtoxins in feedstuff. Applied and Environmental Microbiology, 1976, 32(4): 553-556.
[9]Nan Z B, Liu R. Detection of seed-borne fungi ofAstragalusadsurgens. Acta Prataculturae Sinica, 1997, 6(4): 11-16.
[10]Li C J, Wang Y R, Zhu T H,etal. Responses of alfalfa seed to stress storage conditions. Chinese Journal of Applied Ecology, 2002, 13(8): 957-961.
[11]Zhao L H, Xie G L, Li W,etal. Plant pathology in retrospect and in propect.Seed, 2006, 25(2): 40-42.
[12]Dhingra O, Lustosa D, Maia C,etal. Seedborne fungal pathogens of jacaranda (Dalbergianigra) tree. Seed Science and Technology, 2003, 31(2): 341-349.
[13]Li C J, Nan Z B. Seed-borne fungi of lucerne and their pathogenicity to lucerne seed and seedling. Acta Prataculturae Sinica, 2000, 9(1): 27-36.
[14]Nan Z B. Seed-borne fungi ofAstragalusadsurgens-environment, pathogenicity and control. Acta Prataculturae Sinica, 1998, 7(1): 12-18.
[15]Wiewióra B, Prończuk M.Rhizoctoniasolaniand other fungi detected in perennial ryegrass seeds after harvest and storage. Phytopathologia Polonica, 2002, 26: 19-27.
[16]Yuan Q H, Sun J H, Li Q,etal. Detection of seed-borne fungi onSorghumsudanenseandCoronillavaria. Gansu Animal and Veterinary Sciences, 1993, 23(2): 19-21.
[17]Mao P S, Han J G, Wu X C. Effects of harvest time on seed yield of Siberian Wildrye. Acta Agrestia Sinica, 2003, 11(1): 33-37.
[18]National Bureau of Standard. Rules for Forage Seed Testing(GB 2930-82). Beijing: Technological Standard Press, 1982.
[19]Xu X L, Wu X H, Zhang G Z,etal. Analysis of correlation between seed-associated fungi and seed vigor of sweet corn. Scientia Agricultura Sinica, 2006, 39(8): 1565-1570.
[20]Wei J C.Fungi Idenfication Handbook. Shanghai: Shanghai Science and Technology Press, 1979.
[21]Lu J Y. Pathogenic Phytomycology. Beijing: China Agriculture Press, 2001.
[22]Yang Y X, Han J G, Li S T,etal. Effects of seed-borne fungi on germination and vigor ofZoysiajaponicaseed. Seed, 2006, 25(10): 16-19.
[23]Baskin J M, Baskin C C. A classification system for seed dormancy. Seed Science Research, 2004, 14(1): 1-16.
[24]Liu X Y, Pu M K, Bian Z G,etal. Effect of storage time on seed germination characteristics ofElymussibiricusL. cv.Chuancao No.1. Cao ye Yu Xumu, 2012, (2): 16-18.
[25]Huang X H, Hu X W, Xu Z H,etal. Effects of exogenous hormones on the dormancy and germination ofLeymuschinensis. Acta Prataculturae Sinica, 2013, 22(5): 183-189.
[26]Chen S P. The relationship between seed germination rate and storage time. Seed, 1989, (3): 19-22.
[27]Mao P S, Han J G. Effects of storage treatments on seed vigor of sibiricus wildrye. Pratacultural Science, 2003, 20(4): 16-19.
[28]Meyer S E, Stewart T E, Clement S. The quick and the deadly: growth vs virulence in a seed bank pathogen. New Phytologist, 2010, 187(1): 209-216.
[29]Dalling J W, Davis A S, Schutte B J,etal. Seed survival in soil: interacting effects of predation, dormancy and the soil microbial community. Journal of Ecology, 2011, 99(1): 89-95.
[30]Ruan S L, Xue Q Z. Germination characteristics of seeds under salt stress and physiological basis of salt-tolerance of seedling in hybrid rice. Chinese Journal of Rice Science, 2002, 16(3): 281-284.
[31]Yan H, Zhao W, Chen W J,etal. Effects of different salt solutions pretreatment on the germination ofHelianthusannuus. Seed, 2007, 26(2): 69-71.
[32]Sharada R, Shetty H S. Location and transmission ofMacrophominaphaseoliain black germ seeds. Indina Phytopathology, 1987, 40(2):194-196.
[33]Chen L. Study on Seedborne Fungi ofOnobrychisviciaefolia. Lanzhou: Lanzhou University, 2010.
[34]Zhang Y. Effects of Seed-borne Fungi on the Quality of Perennial Ryegrass (LoliumperenneL.) Seeds. Beijing: China Agricultural University, 2005.
[35]Agarwal V K, Sinclair J B. Principles of Seed Pathology (2nd edn). Boca Raton, FL: CRC Press, 1996.
[36]Wang J H. Seeds-coating technique and its application to establishing turf. Pratacultural Science, 1997, 14(4): 54-56.
[37]Falloon R. Fungicide seed treatment to improve establishment of ryegrasses and other forage plants. Proceedings of the 34th New Zealand Weed and Pest Control Conference. Blenheim, New Zealand, 1981: 43-47.
[38]Nan Z B. Fungicide seed treatments of sainfoin control seed-borne and rootinvading fungi. New Zealand Journal of Agricultural Research, 1995, 38(3): 413-420.
[39]Buchenauer H. Mode of action and selectivity of fungicides which interfere with ergosterol biosynthesis. England:Proceedings 9th British Crop Protection Conference-Pests and Diseases. London,British Crop Protection Council, 1977: 699-711.
[40]Papavizas G, Lewis J. Effect of seed treatment with fungicides on bean root rots. Plant Disease Reporter, 1975, 59: 24-28.
参考文献:
[1]鄢家俊, 白史且, 常丹, 等. 青藏高原老芒麦种质遗传多样性的SSR分析. 中国农学通报, 2010, 26(9): 26-33.
[2]董景实, 张素贞. 老芒麦的研究. 中国草地学报, 1980, 2: 29-33.
[3]陈功, 贺兰芳. 高寒地区两种老芒麦生态适应性和生产性能评价. 草业科学, 2004, 21(9): 39-42.
[4]玉柱, 孙启忠, 邓波, 等. 老芒麦青贮研究. 中国农业科技导报, 2008, 10(1): 98-102.
[5]顾晓燕, 郭志慧, 张新全, 等. 老芒麦种质资源遗传多样性的 SRAP 分析. 草业学报, 2014, 23(1): 205-216.
[6]董世魁, 胡自治, 龙瑞军, 等. 高寒地区多年生禾草混播草地的群落学特征研究. 生态学杂志, 2003, 22(5): 20-25.
[9]南志标, 刘若. 沙打旺种带真菌检测. 草业学报, 1997, 6(4): 11-16.
[10]李春杰, 王彦荣, 朱廷恒, 等. 紫花苜蓿种子对逆境贮藏条件的反应. 应用生态学报, 2002, 13(8): 957-961.
[11]赵丽涵, 谢关林, 李雯, 等. 种子病理学的回顾与展望. 种子, 2006, 25(2): 40-42.
[13]李春杰, 南志标. 苜蓿种带真菌及其致病性测定. 草业学报, 2000, 9(1): 27-36.
[14]南志标. 沙打旺种带真菌——环境, 致病力及防治. 草业学报, 1998, 7(1): 12-18.
[16]袁庆华, 孙建华, 李琪, 等. 苏丹草和小冠花等种子带真菌检验. 甘肃畜牧兽医, 1993, 23(2): 19-21.
[17]毛培胜, 韩建国, 吴喜才. 收获时间对老芒麦种子产量的影响. 草地学报, 2003, 11(1): 33-37.
[18]国家标准局. 牧草种子检验规程(GB 2930-82). 北京: 技术标准出版社, 1982.
[19]徐秀兰, 吴学宏, 张国珍, 等. 甜玉米种子携带真菌与种子活力关系分析. 中国农业科学, 2006, 39(8): 1565-1570.
[20]魏景超. 真菌鉴定手册. 上海: 上海科学技术出版社, 1979.
[21]陆家云. 病原植物真菌学. 北京: 中国农业出版社, 2001.
[22]杨颜霞, 韩建国, 李寿田, 等. 种带真菌对结缕草种子发芽和活力的影响. 种子, 2006, 25(10): 16-19.
[24]刘晓英, 蒲珉锴, 卞志高, 等. 贮藏时间对 “川草 1 号” 老芒麦种子发芽特性的影响. 草业与畜牧, 2012, (2): 16-18.
[25]黄晓辉, 胡小文, 徐宗海, 等. 羊草种子休眠和萌发的激素调控研究. 草业学报, 2013, 22(5): 183-189.
[26]陈叔平. 贮存年限与种子发芽率关系. 种子, 1989, (3): 19-22.
[27]毛培胜, 韩建国. 贮藏处理对老芒麦种子活力的影响. 草业科学, 2003, 20(4): 16-19.
[30]阮松林, 薛庆中. 盐胁迫条件下杂交水稻种子发芽特性和幼苗耐盐生理基础. 中国水稻科学, 2002, 16(3): 281-284.
[31]颜宏, 赵伟, 陈文静, 等. 不同盐溶液浸种对向日葵种子萌发的影响. 种子, 2007, 26(2): 69-71.
[33]陈林. 红豆草 (Onobrychisviciaefolia) 种带真菌研究. 兰州:兰州大学, 2010.
[34]张颖. 种带真菌对多年生黑麦草 (LoliumperenneL.) 种子质量的影响. 北京: 中国农业大学, 2005.
[36]王建华. 种子包衣技术及其在草坪建植中的应用. 草业科学, 1997, 14(4): 54-56.
Seed-borne fungi infection of Siberian wildrye: Effects on seed germination and seedling growth
CHEN Tao, NAN Zhibiao*
StateKeyLaboratoryofGrasslandAgro-ecosystems,CollegeofPastoralAgriculturalScienceandTechnology,LanzhouUniversity,Lanzhou730020,China
Abstract:We investigated seed-borne fungi in Siberian wildrye seed samples collected from Qinghai in five different harvest seasons (S1to S5) and tested the effects of fungal infection on seed germination and seedling growth. Seed germination ranged from 56%to 80%. S2had the highest seed germination (80%), significantly higher than S1and S5(P<0.05).The fungal infection rate of seed samples varied from 24% to 38% and declined with the extension of storage time.S5had the highest infection rate of 38%, significantly higher than all other samples (P<0.05).17 fungal species from 15 genera were identified; the isolation rate ranged from 0.25% to 8.75%. Penicillium sp. and Aspergillus sp. were the most common seed-borne species found and were isolated in all seasons. Pathogenicity tests showed that Fusarium avenaceum,F. monilifome,Fusarium sp.1,Drechslera sp. and Bipolaris sp. were the most pathogenic fungi, significantly reducing seed germination and seedling growth (P<0.05). Alternaria alternata significantly reduced seedling growth but did not affect seed germination. Pithomyces sp.,Aspergillus sp. and Bipolaris sp. significantly prolonged the mean seed germination period, while Fusarium avenaceum significantly shortened it (P<0.05).
Key words:Siberian wildrye (Elymus sibiricus);germination;seed-borne fungi;pathogenicity
*通讯作者
Corresponding author. E-mail:zhibiao@lzu.edu.cn
作者简介:陈焘(1989-),男,甘肃会宁人,在读博士。E-mail:chent2007@lzu.edu.cn
基金项目:国家“973”项目(2014CB138702)资助。
*收稿日期:2013-09-03;改回日期:2014-05-12
DOI:10.11686/cyxb20150212
http://cyxb.lzu.edu.cn
陈焘, 南志标. 不同储存年限老芒麦种子种带真菌检测及致病性测定. 草业学报, 2015, 24(2): 96-103.
Chen T, Nan Z B. Seed-borne fungi infection of Siberian wildrye: Effects on seed germination and seedling growth. Acta Prataculturae Sinica, 2015, 24(2): 96-103.
猜你喜欢
植物研究(2019年2期)2019-03-19
种子科技(2018年11期)2018-09-10
许昌学院学报(2018年8期)2018-09-05
农民致富之友(2018年9期)2018-06-27
中成药(2017年12期)2018-01-19
湖南畜牧兽医(2016年3期)2016-06-05
湖南畜牧兽医(2016年3期)2016-06-05
天津农林科技(2015年1期)2015-12-30
川北医学院学报(2015年5期)2015-12-05
西藏科技(2015年5期)2015-09-26
