
罗平生物群的发现与研究进展——据云南1∶5万罗平县等四幅区调和专题调查成果
2015-02-24 05:17张启跃胡世学文芠周长勇谢韬黄金元
中国地质调查 2015年2期
关键词:生态系统
张启跃, 胡世学, 文芠, 周长勇, 谢韬, 黄金元
(中国地质调查局成都地质调查中心,四川 成都 610081)
罗平生物群的发现与研究进展——据云南1∶5万罗平县等四幅区调和专题调查成果
张启跃, 胡世学, 文芠, 周长勇, 谢韬, 黄金元
(中国地质调查局成都地质调查中心,四川 成都610081)
摘要:成都地质调查中心云南区调项目组2007年在罗平县中三叠统关岭组二段中发现了罗平生物群。罗平生物群是三叠纪的化石宝库,化石包含了脊椎动物和无脊椎动物。脊椎动物以海生爬行类和鱼类为主。无脊椎动物以节肢动物为主,伴生有双壳类、腹足类、菊石、棘皮动物、腕足类、牙形石、有孔虫和植物类化石。牙形石研究显示罗平生物群处于Nicoraella kockeli牙形石带,时代上属于中三叠世安尼期Pelsonian亚期。罗平生物群保存在一个局限的台盆环境中。其化石的特异保存很可能与底层水缺氧和微生物席的密封作用有关。罗平生物群目前是研究三叠纪海洋生态系统复苏的最好的一扇窗口,也是三叠纪海生鱼类和节肢动物化石保存最好的产地之一,为研究二叠纪末大绝灭后海洋生态系统复苏以及中生代海洋生态系统的演化历史提供了新材料。
关键词:罗平生物群;三叠纪;复苏;生态系统;
0引言
云南1∶5万大同(G48E020009)、圭山(G48E020010)、罗平县(G48E019010)、捧乍(G48E019011)幅区域地质调查项目(项目编号:1212010610211)由中国地质调查局成都地质调查中心承担。该项目历时5年,野外工作完成填图面积 1 861 km2,地质路线 2 886 km,地质观察点4 923个,实测产状6 653个,照片17 065幅。实测了29条地层剖面计49.89 km, 2条构造剖面计11.07 km。采集样品总数5 719件。针对罗平生物群分布区主要完成了2.5万填图面积200 km2、地质调查路线655 km、实测地层剖面3.84 km,施工槽探10 000 m3,采获化石5 700余件。2012年6月提交了成果报告①张启跃, 周长勇, 楼雄英,等. 云南1∶5万大同、圭山、罗平县、捧乍幅区域地质调查报告[R].成都:中国地质调查局成都地质调查中心, 2012.②张启跃, 胡世学, 周长勇,等. 罗平生物群综合研究报告[R].成都:中国地质调查局成都地质调查中心, 2012.③张启跃, 周长勇,谢韬,等. 1∶5万区域地质图(大同幅、圭山幅、罗平县幅、捧乍幅)及分幅说明书[R].成都:中国地质调查局成都地质调查中心, 2012.④张启跃, 周长勇, 李军敏,等. 1∶2.5万云南省罗平县大凹子地区地质图及说明书[R].成都:中国地质调查局成都地质调查中心, 2012.,报告对测区内岩石地层、生物地层、层序地层与沉积相、岩石、遥感解译、构造背景、构造单元特征、地质发展史、罗平生物群综合研究、矿产地质特征、经济地质和生态环境等特征进行了全面而系统的描述与总结。
2012年10月26日由中国地质调查局组织专家组在成都进行了项目成果评审,专家组认为区域地质调查报告及专题报告反映了很高的工作和研究程度,罗平生物群[1]的发现和研究具有重大的科学意义,并产生了巨大的社会影响,一致建议通过评审。质量评为96分,为优秀级。专家组鉴于罗平生物群是显生宙三大生物演化辐射之一,在地球生命和环境协同演化中具有全球意义,对于探讨二叠纪末生物大绝灭后三叠纪海洋生物的复苏机制有重要意义,建议有关部门对于该项目的后续研究给予高度重视、大力支持。
1区域地质概况
联测图幅位于云南、贵州、广西三省交界,弥勒—师宗断层南东侧,大地构造位置位于南盘江—右江盆地北西边缘(图1)。研究区属华南地层大区,东南地层区,扬子地层分区和右江地层分区,出露地层主要是中生界三叠系,另有少量上古生界、古近系和第四系零星分布。
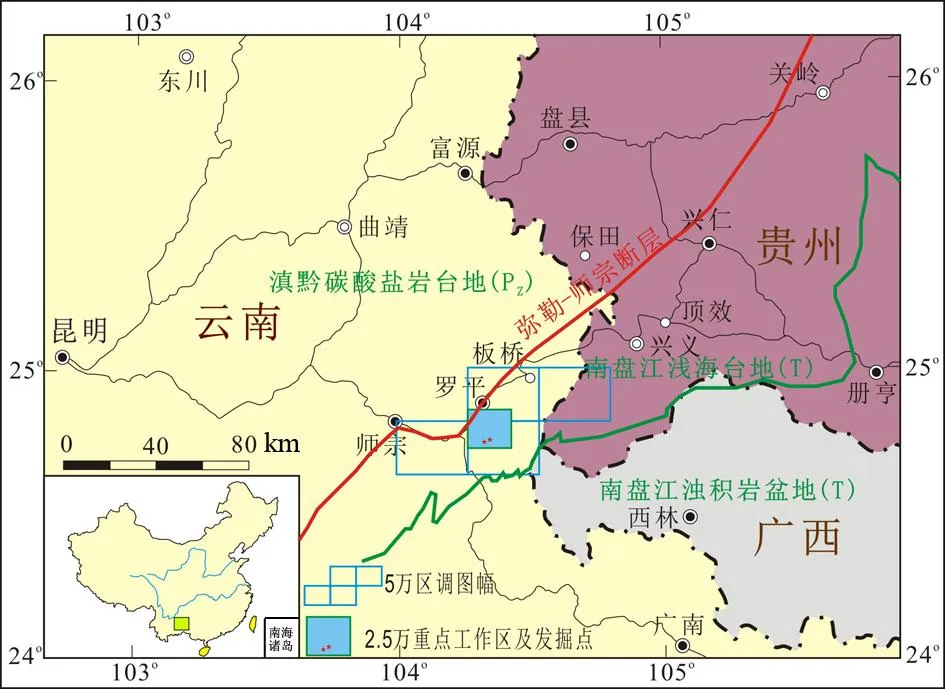
图1 工作区及大地构造位置图
区内中下二叠统为连续的海相沉积建造,未发育梁山组,中二叠统阳新组与龙潭组为嵌入不整合接触,未发育玄武岩。早三叠世为台地相与浅海陆棚相过渡区。早期(飞仙关组)为泥岩、粉砂岩夹生物屑灰岩、鲕状灰岩;晚期(嘉陵江组)为鲕状灰岩、内碎屑灰岩、蠕虫状灰岩夹杂色砂泥岩。中三叠世安尼期为潮坪相沉积。关岭组一段为钙质粉砂泥岩、泥岩夹泥质白云岩,底部见“绿豆岩”,属潮下、澙湖、潮间环境沉积;关岭组二段出现较多色深、层薄的泥质碳酸盐沉积,含较多燧石结核和燧石条带,产罗平生物群化石。上部逐渐过渡为白云岩、白云质灰岩夹角砾灰岩,以含膏盐假晶和盐溶角砾为特征的潮上蒸发坪沉积。中三叠世拉丁期关岭组三段(杨柳井段)为半闭塞台地相沉积,发育泥微晶白云岩、层纹石白云岩、角砾状白云岩等。藻纹层及鸟眼构造十分发育,生物稀少,属潮间—潮上滨海环境。关岭组四段(竹杆坡段)为台地边缘相—陆棚斜坡相沉积,以灰白色核形石灰岩、泥微晶砂屑砾屑灰岩为主,产贵州龙动物群化石。右江地层分区中拉丁期兰木组为盆地相砂泥质浊流沉积。卡尼期法郎组为浅海陆棚—陆棚边缘盆地相沉积,下部(小凹灰岩段)为灰黑色薄层泥晶灰岩,产关岭动物群化石。上部(赖石科碎屑岩段)为灰、灰黑色细碎屑岩。水平层理较发育。诺利期把南组、火把冲组为滨海平原沼泽相沉积,以页岩、粉砂岩为主夹细砂岩,局部互层,并夹煤层或透镜体,普遍具大型楔状、板状交错层理及水平层理。
2007年10月笔者在罗平大凹子村关岭组第二段泥晶灰岩中首次发现鱼类化石,成都地质调查中心丁俊主任等领导十分重视这一发现,建议向中国地质调查局申请经费深入开展研究工作。中国地质调查局经过专家论证批准了增量项目申请,同意在区调项目中增加古生物化石及地层研究专题,并将项目延期到2011年。项目组随后通过1∶2.5万区调填图、精细剖面测量以及在上石坎、门前坡两次化石采掘等工作,初步查明了罗平生物群分布特征、沉积背景及其生物面貌特征等。
2罗平生物群研究取得的主要进展
2.1 揭示了罗平生物群的基本面貌
项目组详细填绘了罗平生物群的分布范围①张启跃, 周长勇, 楼雄英,等. 云南1∶5万大同、圭山、罗平县、捧乍幅区域地质调查报告[R].成都:中国地质调查局成都地质调查中心, 2012.②张启跃, 胡世学, 周长勇,等. 罗平生物群综合研究报告[R].成都:中国地质调查局成都地质调查中心, 2012.③张启跃, 周长勇,谢韬,等. 1∶5万区域地质图(大同幅、圭山幅、罗平县幅、捧乍幅)及分幅说明书[R].成都:中国地质调查局成都地质调查中心, 2012.④张启跃, 周长勇, 李军敏,等. 1∶2.5万云南省罗平县大凹子地区地质图及说明书[R].成都:中国地质调查局成都地质调查中心, 2012.,根据牙形石成果厘定了罗平生物群时代为中三叠世安尼期Pelsonian亚期,牙形石Nicoraellakockeli带[2]。在同沉积的凝灰岩夹层中获得244.2 Ma的锆石SHRIMP U-Pb年龄②张启跃, 胡世学, 周长勇,等. 罗平生物群综合研究报告[R].成都:中国地质调查局成都地质调查中心, 2012.,并且在关岭组一段底部获得(246.6±1.4) Ma LA-ICP MS锆石U-Pb年龄[3]。初步研究揭示罗平生物群是目前世界上已知的多样性最为丰富的三叠纪海生化石库之一,代表了二叠纪末生物大绝灭后海洋生态系统的全面复苏,是中三叠世生物大辐射的典型代表[4-6]。罗平生物群是由多个门类构成的完整海洋生态系统,包括了海生爬行类的鱼龙[6-9]、原龙[4]、鳍龙类[7-13]等;鱼类包括了辐鳍鱼亚纲软骨硬鳞鱼次纲的古鳕鱼类、龙鱼类、裂齿鱼类、肋鳞鱼类,辐鳍鱼亚纲新鳍次纲的半椎鱼类和鲱口类,肉鳍鱼亚纲空棘鱼类等[4,6, 14,15];罗平生物群目前发现的化石以甲壳纲为主[4],另外还发现有螯肢亚门的肢口纲鲎类化石和多足亚门倍足纲的千足虫类化石[4,16]。值得指出的是,罗平生物群甲壳类数量巨大,种类多样,包括有糠虾类、十足目、等足目[17]、叶肢介以及可能属于甲壳纲的园蟹类等;软体动物门的菊石、双壳、腹足、箭石等;腕足动物门的舌形贝类;棘皮动物门的海胆、海星、海百合和海参;还包括牙形类、有孔虫类、植物化石(松柏类)和遗迹化石[1,4,18-21]。采获了罗平生物群化石5 000余件,目前已鉴定命名的化石有6门40属100多种②张启跃, 胡世学, 周长勇,等. 罗平生物群综合研究报告[R].成都:中国地质调查局成都地质调查中心, 2012.,其中30个新种为罗平发现并命名。在国土资源部公布的《国家重点保护古生物化石名录(首批)》中,罗平生物群有4种化石被列入该名录,其中丁氏滇肿龙(Dianopachysaurusdingi)[11]、利齿滇东龙(Diandongosaurusacutidentatus)[13]、云贵中国龟龙(Sinosaurosphargisyunguiensis)[22]被列为一级重点保护古生物化石,云南龙鱼(Saurichthysyunnanensis)[23]被列为三级重点保护古生物化石。
罗平生物群化石主要分布于薄层泥晶灰岩及泥灰岩中,含化石层位中滑塌构造[24]和微生物席[25]等沉积构造十分发育,总体上看固着生物极为少见,生物扰动程度也较低,水平层理发育,绝大多数生物化石保存完整,很少发现古生物死亡后身体腐烂而保存不完整,说明在罗平生物群生活时期的大部分时间底层水体是处于缺氧或贫氧的环境[26-28]。罗平生物群的全岩碳、氧同位素测定结果显示,δ13C发生了明显的负偏移,δ13C负偏移的原因主要受火山作用和沉积水体缺氧有关。运用δ18O值算出当时海水古温度分布在27~35 ℃之间,平均为30.92 ℃左右,说明中三叠世安尼期罗平地区的气候较为温暖[29]。
罗平生物群保存在一个局限的澙湖环境中,生物群的死亡和埋藏与突发性事件有着密切的关系,火山、地震(或海啸)等事件破坏了生物群赖于生活的海洋水体环境,另外海平面下降和盆地面积的不断缩小,也引发了水动力条件减弱,与外界广阔海水循环受阻,水体含盐度增高,水体成分发生变化,导致缺氧、滞流的环境,其化石的特异保存很可能与底层水缺氧和微生物席的密封作用有关。
2.2 发现了幻龙觅食足迹
中生代(252~66 Ma),当恐龙雄霸陆地时,海洋是由各种各样的海洋爬行动物统治着的。最早的一支族群是幻龙类,它们具有扁长的类似桨状的四肢和细长的身躯,是两栖类食肉动物,在三叠纪中期(2.45亿年前)海岸边活动,是那个时代的顶级掠食者,是海洋生态系统从二叠纪末生物大绝灭的灾难中完全复苏、中生代海洋生态系统建立的标志。由于海生爬行类在水下活动的遗迹很难保存为化石,对海生爬行动物运动方式的研究主要依靠形态功能学和与现生类型的对比,长期以来对其生活方式的认识一直较为模糊。项目组在罗平生物群研究过程中,首次在罗平县大凹子等地发现了数百枚保存完好的海生爬行类足迹,定名为罗平双桨迹DikoposichnusluopingensisZhang et al., 2014[30](图2,图3)。确定了其属于中生代海洋爬行动物——幻龙类在水下觅食所留下的足迹。研究结果显示,这些爬行动物在海底移动依靠其前肢协调地触地向前,这是幻龙类海洋生物在水下运动模式得以发现的第一个直接证据。该研究成果将为深化三叠纪海洋生物复苏这一重大地质事件,探讨罗平生物群生态与环境变化之间的关系提供新材料。

图2 罗平双桨迹El4

图3 罗平双桨迹Er4
足印遗址位于东经104°18′39″、北纬24°46′12″。门前坡采点现采场第107层面上保留120 m2范围中已发现各种海生爬行类动物足印化石300余个,其中有245个足迹有规律组成15条行迹。从这些有规律的行迹上来看,爬行动物的足迹是一种步履徜徉、悠闲觅食的轻松场面(图4)。

图4 幻龙在水下运动方式复原图
2.3 发现了张氏幻龙、丁氏滇龙等海生爬行动物
罗平生物群中的海生爬行类动物是2.44亿年前海洋生物系统全面复苏的代表。罗平生物群海生爬行类动物化石主要有鳍龙类(Sauropterygia)、鱼龙类(Ichthyopterygia)、海龙类(Thalattosauria)以及原龙类(Protosauria)。罗平生物群新发现命名的爬行动物化石主要有:丁氏滇龙DianopachysaurusdingiLiu et al.,2011[11]; 利齿滇东龙DiandongosaurusacutidentatusShang et al., 2011[13];云贵中华龟龙SinosaurosphargisyunguiensisLi et al., 2011[22];多腕骨大头龙LargocephalosauruspolycarponChen et al., 2013[31];奇特滤齿龙AtopodentatusunicusChen et al., 2014[12];巨型张氏幻龙NothosauruszhangiLiu et al., 2014[10]。其中以张氏幻龙、丁氏滇龙最具代表意义。
张氏幻龙属于中生代占海洋统治地位的鳍龙,张氏幻龙有迄今已知的三叠纪所有鳍龙中最大的完整下颌(图5)。张氏幻龙具有大型而锋利的圆锥形犬齿,其躯体总长估计有5~7 m,主要以伏击的方式捕食大型肉食性鱼类和其他海生爬行类动物。二叠-三叠纪大灭绝重塑了进化发展的方向,为现代生态系统结构的建立开辟了空间,导致了一些新的类群的出现,这其中包括鱼龙、蛇颈龙以及沧龙等在内的海生爬行类。当时生态系统的复苏在全球是否同步一直是科学界有待解决的一个难题。在张氏幻龙被发现之前,巨型的顶级捕食者在古代的特提斯洋东侧/古太平洋西侧是缺失的。现在随着这一巨型顶级捕食者在东特提斯洋的发现,我们有理由相信当时全球的浅海生态系统在中三叠世早期已经全面复苏。
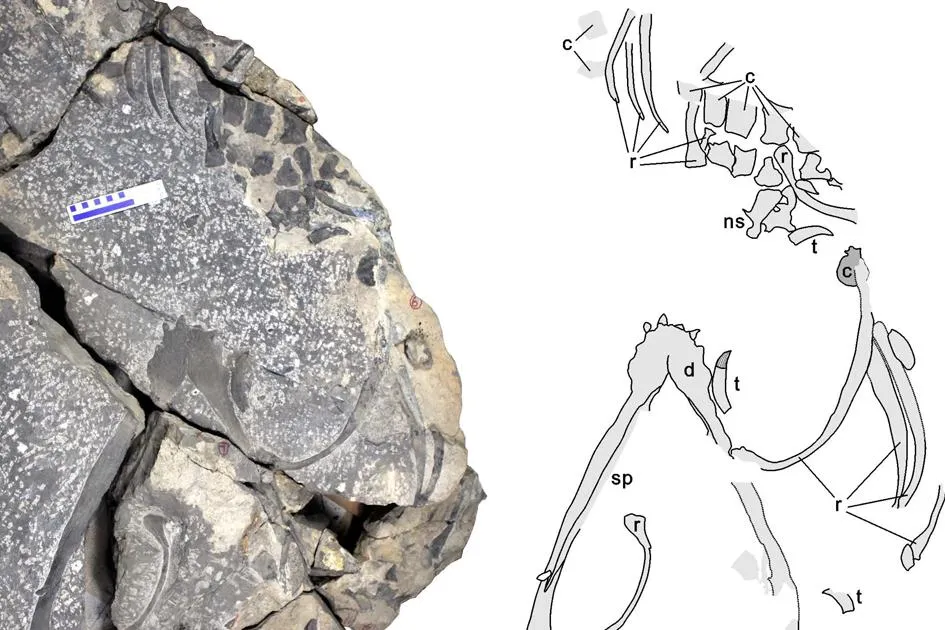
图5 张氏幻龙(引自Liu et al., 2014)
丁氏滇龙是一种食肉类的小型肿肋龙,牙齿异常锋利(图6)。丁氏滇龙的研究验证了15年前提出的肿肋龙起源于中国的生物地理学假说。恢复了中国古脊椎动物学的开创人杨钟健先生于1965年所建立的贵州龙科,此科包括贵州龙和滇龙两个属。分支分析的结果显示,滇龙构成了贵州龙的姐妹群,它们共同组成了欧洲肿肋龙的外类群,滇龙的发现大大缩短了这一生物地理假说所要求的谱系类群的存在[11]。

图6 丁氏滇龙(引自Liu et al.,2011)
2.4 发现了高背罗雄鱼、罗平空棘鱼等鱼类新属种
罗平生物群的鱼类化石数量丰富且种类繁多,初步发现有5目,7科,21属,26种,新命名的有19种(表1)。包含了三叠纪鱼类化石中的典型类型,分别有古鳕类、龙鱼类、肋鳞鱼类、裂齿鱼类、新鳍鱼类和空棘鱼类。新鳍鱼类是出现在晚二叠世,繁盛于中生代,一直延续到现代的真骨鱼类。新鳍鱼类与原始辐鳍鱼类在结构上有了很大的进步。上颌骨不再与前鳃盖相铰接,下颌具有冠状突,前鳃盖几乎垂直且背枝细长,具间鳃盖,每块支持骨只支持一根鳍条,尾鳍半歪尾型。支持骨与鳍条对应关系的改变使得新鳍鱼类能更好地控制背鳍和臀鳍,有了更灵活的游泳能力。颌部结构的改变使它们有强的咬合力,捕食能力得到加强。罗平生物群中新鳍鱼类包含有体型异化的高背罗雄鱼(图7)和格兰德拱鱼,有身体几乎无鳞的意外裸鱼和安氏马可波罗鱼,最普遍的苏氏圣乔治鱼,还有罗平强壮鱼。空棘鱼繁盛于三叠纪,其中的一支拉蒂迈鱼仍生活在现代印度洋的深海里,被称为“活化石”。罗平生物群中宽泪颧骨罗平空棘鱼标本内的两个胚胎显示了该类动物最早的卵胎生证据(图8),把晚侏罗世的记录提前到了中三叠世。罗平生物群中鱼类化石在属一级上与欧洲三叠纪化石群相同,证明了在中三叠世古特提斯洋的东西两岸的化石群存在紧密的联系[6,14,15,32]。

图7 高背罗雄鱼(引自Wen et al.,2012)

图8 宽泪颧骨罗平空棘鱼(引自Wen et al.,2013)

序号罗平生物群新发现命名的鱼类化石1大洼子龙鱼SaurichthysdawaziensisWuetal.,2009[33]2云南龙鱼SaurichthysyunnanensisZhangetal.,2010[23]3长奇鳍中华龙鱼SinosaurichthyslongimedialisWuetal.,2011[34]4小型中华龙鱼SinosaurichthysminutaWuetal.,2011[34]5贝氏罗平鱼LuopingichthysbergiSunetal.,2009[35]6苏氏罗平裂齿鱼LuopingperleidussuiGeng,2012[36]7中华裂齿鱼PerleidussinensisLombardo,2011[37]8邓氏富源裂齿鱼FuyuanperleidusdengiGeng,2012[36]9齿状滇东裂齿鱼DiandongperleidusdenticulatusGeng,2012[36]10包氏海博鱼HabroichthysbroughiLinetal.,2011[38]11苏氏圣乔治鱼SangiorgioichthyssuiLópez-arbarello,2011[39]12高背罗雄鱼LuoxiongichthyshyperdorsalisWenetal.,2012[14]13格兰德拱鱼KyphosichthysgrandeiXuetal.,2012[40]14意外裸鱼GymnoichthysinoponatusTintorietal.,2010[41]15安氏马可波罗鱼MarcopoloichthysaniTintorietal.,2008[32]16宽泪颧骨罗平空棘鱼LuopingcoelacanthuseurylacrimalisWenetal.,2013[15]17尖瘤云南空棘鱼YunnancoelacanthusacrotuberculatusWenetal.,2013[15]18罗平强壮鱼RobustichthysLuopingensisXuetal.,2014[42]19尼尔森翼鳕PteronisculusnielseniXuetal.,2014[43]
2.5 发现了罗平云南鲎、中华三指龙虾等节肢类新属种
节肢动物是罗平生物群数量最为丰富的门类,以甲壳纲为主,还有螯肢亚门的肢口纲鲎类、千足虫类。它们占罗平生物群化石数量的90%以上,包括糠虾类、十足目、等足目、叶肢介等,新命名的节肢动物有9种[4,6,44-46](表2)。罗平生物群是甲壳类节肢动物在中生代初期爆发性辐射的典型代表,许多种类如等足目化石和十足目的虾类(图9)、龙虾类化石都是首次在我国发现,对研究节肢动物在经历二叠纪末大绝灭后的复苏和辐射模式具有重要的科学意义。罗平生物群中的等足类化石与发现于澳大利亚中三叠世的等足目化石十分相似,这似乎说明当时南中国板块和澳大利亚板块并不像目前认为的那么遥远。十足目龙虾类化石是目前已知保存最为完好、数量最丰富、时代最早的龙虾化石群之一。对研究该类动物的起源演化及古生态学具有重要的意义。三叠纪的原蟹类化石此前偶有报道,主要见于欧洲和马达加斯加等地,罗平生物群中的原蟹类化石是该类化石在中国的首次发现。鲎类化石(图10)在中国的首次发现扩大了中生代鲎类节肢动物的分布范围,也为研究鲎类的演化、生态变迁等提供了重要的信息。

图9 壮美异形伊吉虾

图10 罗平云南鲎

序号罗平生物群节肢动物新发现命名化石1罗平云南鲎YunnanolimulusluopingensisZhangetal.,2009[16]2白氏原双节虫ProtamphisopusbaiiFuandWilson,2010[17]3罗平棒臂龙虾KoryncheirosluopingensisFeldmann,SchweitzerandZhang,2012[44]4中华三指龙虾TridactylastacussinensisFeldmann,SchweitzerandZhang,2012[44]5施氏云南龙虾YunannopalinuraschramiFeldmann,SchweitzerandZhang,2012[44]6泸西伊吉虾AegerluxiiHuang,FeldmannandSchweitzer,2013[45]7短额安尼伊吉虾AnisaegerbrevirostrusSchweitzeretal.,2014[46]8具刺安尼伊吉虾AnisaegerspiniferusSchweitzeretal.,2014[46]9壮美异形伊吉虾DistaegerprodigiosusSchweitzeretal.,2014[46]
2.6 开展了数字化建档工作,成功申报了野外科学观测研究基地和国家地质公园
古生物化石被发掘出来以后,失去了它原来的环境条件,裸露在空气、阳光和雨水中。由于所处条件的变化,化石容易被风化或腐蚀,日渐失去它本来的面目,造成无法弥补的损失。为了实现化石的长久保存和研究,也便于今后对化石的反复展示,项目组与四川大学光电学院合作对化石进行三维数字化处理建档工作。成功地采用非接触式激光扫描仪的测量原理对罗平古生物化石群中野外分布的古生物足迹化石进行了数字化测量,采用大视场wide和小视场tele镜头交替使用的方法获取全场和单一足迹化石三维点云数据,再利用三维数据处理Polyworks软件将两个镜头的三维数据进行对应的嵌入融合,最终测量结果既能全面记录足迹整体分布,真实反映古生物爬行运动轨迹,又能精细反映足迹局部特征,为今后类似工作提供了一个可借鉴的方法[47]。
经成都地质调查中心与罗平县政府的共同努力,罗平生物群于2011年被国土资源部命名为“全国首批野外科学观测研究基地”。2011年10月罗平生物群以全国最高评分入选第六批国家地质公园。罗平县属于“国家集中连片特困地区——滇、桂、黔石漠化片区”,罗平生物群国家地质公园的建设将为罗平旅游业的发展和地方经济建设开辟新的方向[48,49]。
3展望与思考
罗平生物群的发现,曾引起来自英国、德国、澳大利亚、美国、意大利、苏格兰、瑞士、阿根廷等多个国家的古生物学专家学者先后到罗平生物群野外实地考察,专家们考察后认为,罗平生物群是二叠纪末期生物灭绝后三叠纪海洋生态系统全面复苏和辐射最典型的代表,与著名的世界自然遗产Monte San Giorgio动物群相比而言,罗平生物群种类更加丰富,保存更加完整,规模更加庞大,具有十分重要的科研价值。
作为我国近年来新发现的特异埋藏生物群的典型代表,罗平生物群的研究在国际上已经产生了较大的影响,但作为一个世界级的化石库,与澄江动物群、热河生物群相对而言其研究程度还处于初级阶段,还有着巨大的研究潜力和发展空间。还需要充分揭示罗平生物群的面貌和生物多样性,揭示罗平生物群在海生爬行类、鱼类、节肢动物等主要门类演化方面的独特意义;在此基础上结合其他三叠纪生物群,进一步探讨三叠纪海洋生态系统复苏和生物辐射机制这一国际古生物学界的热点问题。可望在三叠纪海洋生态系统复苏、生物辐射化石记录方面取得一些新的重大发现,对这一时期地球-生命系统演化过程中若干关键科学问题的认识上取得一批重要的原创性成果,形成一支在国内外有较大影响并具有一定学术优势的团队,为罗平生物群的可持续发展、为申报世界地质公园和世界自然遗产奠定基础。
致谢:项目取得的成果与成都地质调查中心丁俊、王剑、尹福光、江新胜、潘桂棠、刘宝珺,中国地质调查局翟刚毅、庄育勋,中国地质大学(武汉)殷鸿福、陈中强、张克信,中国地质博物馆卢立伍,英国布里斯托大学Michael Benton、美国肯特州立大学Feldmann R. M.和Schweitzer C. E.,罗平县刘腾林、吕连松、尹振中等专家领导的关心和指导是分不开的,参加本项目工作的还有楼雄英、吕涛、李军敏、林仕良、刘伟、彭智敏、李富、 毛晓长、白建科、刘俊、罗茂等同志,陈克强同志审阅了文稿,并提出建设性意见,在此一并深表谢意。
参考文献:
[1]张启跃,周长勇,吕涛,等.云南罗平中三叠世安尼期生物群的发现及其意义[J].地质论评,2008,54(4):145-149.
[2]Zhang Q Y,Zhou C Y,Lu T,et al.A Conodont-based Middle Triassic age assignment for the Luoping Biota of Yunnan,China [J].Science In China Ser.D:Earth Sciences,2009,52(10):1673-1678.
[3]谢韬,周长勇,张启跃,等.罗平生物群下部凝灰岩锆石年龄及其地质意义[J].地质论评,2013,59(1):159-164.
[4]Hu S X,Zhang Q Y,Chen Z Q,et al.The Luoping biota:exceptional preservation,and new evidence on the Triassic recovery from end-Permian mass extinction[J].Proceeding of the Royal Society,Series B,2011,278:2274-2282.
[5]Chen Z Q,Benton M J.The timing and pattern of biotic recovery following the end-Permian mass extinction[J].Nature Geoscience,2012,5:375-383.
[6]Benton M J,Zhang Q Y,Hu S X,et al.Exceptional vertebrate biotas from the Triassic of China,and the expansion of marine ecosystems after the Permo-Triassic mass extinction[J].Earth Science Reviews,2013,125:199-243.
[7]Liu J,Motani R,Jiang D Y,et al.The first specimen of the Middle Triassic PhalarodonAtavus(Ichthyosauria:Mixosauridae)from South China,showing postcranial anatomy and Peri-Tethyan distribution[J].Palaeontology,2013,56(4):849-866.
[8]Liu J,Jonathan C A,Sun Y Y,et al.New Mixosaurid Ichthyosaur specimen from the Middle Triassic of SW China:further evidence for the diapsid origin of Ichthyosaurs[J].Journal of Paleontology,2011,85(1):32-36.
[9]陈孝红,程龙.混鱼龙(爬行动物:鱼龙类)在云南罗平中三叠统的发现[J].地质学报,2009,83(9):1214-1220.
[10]Liu J,Hu S X,Olivier R,et al.A gigantic nothosaur(Reptilia:Sauropterygia)from the Middle Triassic of SW China and its implication for the Triassic biotic recovery[J].Scientific Report,2014,4,7142;DOI:10.1038/srep07142.
[11]Liu J,Olivier R,Jiang D Y,et al.A new pachypleurosaur(Reptilia:Sauropterygia)from the Middle Triassic of southwestern China and the phylogenetic relationships of Chinese pachypleurosaurs[J].Journal of Vertebrate Paleontology,2011,31(2):292-302.
[12]Cheng L,Chen X H,Shang Q H,et al.A new marine reptile from the Triassic of China, with a highly specialized feeding adaptation[J].Naturwissenschaften,2014,DOI 10.1007/s 00114-014-1148-4.
[13]Shang Q H,Wu X C,Li C.A new eosauropterygian from the Middle Triassic of eastern Yunnan Province,southwestern China[J].Vertebrata PalAsiatica,2011,49:155-173.
[14]Wen W,Zhang Q Y,Zhou C Y,et al.A new genus of basal actinopterygian fish from the Anisian(Middle Triassic)of Luoping,Yunnan Province,Southwest China[J].Actapalaeontologicapolonica,2012,57(1):149-160.
[15]Wen W,Zhang Q Y,Hu S X,et al.Coelacanths from the Middle Triassic Luoping Biota,Yunnan,South China,with the earliest evidence of ovoviviparity[J].Acta Palaeontologica Polonica,2013,58(1):175-193.
[16]张启跃,胡世学,周长勇,等.鲎类化石(节肢动物)在中国的首次发现[J].自然科学进展,2009,19(10):55-58.
[17]Fu W L,Wilson G D F,Jiang D Y,et al.A new species of Protamphisopus Nicholls(Crustacea,Isopoda,Phreatoicidea)from Middle Triassic Luoping Fauna of Yunnan Province,China[J].Journal of Paleontology,2010,84:1001-1011.
[18]黄金元,张克信,张启跃,等.云南中三叠世罗平生物群中首次发现牙形石齿串[J].地球科学——中国地质大学学报,2010,35(4):512-514.
[19]黄金元,张克信,张启跃,等.云南罗平大凹子与上石坎剖面牙形类动物群研究进展[J].地质科技情报,2011,30(3):1-17.
[20]黄金元,胡世学,张启跃,等.云南中三叠世罗平生物群中海参骨片化石的发现[J].地质科技情报,2013,32(1):139-142.
[21]Hu S X,Zhang Q Y,Zhou C Y.Fossil Coprolites from the Middle Triassic Luoping Biota and Ecological Implications[J].Journal of Earth Science,2010,21(Special Issue):191-193.
[22]Li C,Olivier R,Wu X C,et al.A new Triassic marine reptile from southwestern China[J].Journal of Vertebrate Paleontology,2011,31:303-312.
[23]张启跃,周长勇,吕涛,等.云南罗平地区中三叠世龙鱼化石的发现[J].地质通报,2010,29(1):26-30.
[24]白建科,张启跃,尹福光,等.云南罗平生物群埋藏环境初步研究:来自沉积构造的证据[J].沉积学报,2010,28(3):762-767.
[25]Luo M,Chen Z Q,Hu S X,et al.Carbonate reticulated ridge structures from the Lower Middle Triassic of The Luoping area,Yunnan,Southwestern China:Geobiologic features and implications for exceptional preservation of the Luoping Biota[J].Palaios,2013,28:541-551.
[26]白建科,张启跃,尹福光,等.云南罗平中三叠统关岭组二段碳酸盐岩微相分析[J].沉积与特提斯地质,2009,29(3):15-21.
[27]周长勇,张启跃,吕涛,等.云南中三叠世罗平生物群产出地层的地球化学特征和沉积环境[J].地质论评,2014,60(2):285-298.
[28]Zhou C Y,Zhang Q Y,Hu S X,et al.Geochemical records of paleoenvironment Associated with the Middle Triassic Luoping Biota,Yunnan,Southwest China[J].Journal of Earth Science,2010,21(Special Issue):225-227.
[29]孙媛媛,刘俊,吕涛,等.罗平生物群产出层位氧碳稳定同位素的初步研究[J].地质学报,2009,83(8):1110-1117.
[30]Zhang Q Y,Wen W,Hu S X,et al.Nothosaur foraging tracks from the Middle Triassic of southwestern China[J].Nature.Commun,2014,5:3973.
[31]Cheng L,Chen X H,Zeng X W,et al.A new Eosauropterygian(Diapsida:Sauropterygia)from the Middle Triassic of Luoping,Yunnan Province[J].Journal of Earth Science,2013,23:33-40.
[32]Tintori A,Sun Z Y,Lombardo C,et al.Rusconi M.& Hao W.C.new specialized basal neopterygians(Actinopterygii)from Triassic of the Tethys realm[J].GeologiaInsbrica,2008,10/2(2007):13-20.
[33]Wu F X,Sun Y L,Hao W C,et al.New species of Saurichthys(Actinopterygii:Saurichthyidae)from Middle Triassic(Anisian)of Yunnan Province,China[J].Acta Geologica Sinica,2009,83(3):440-450.
[34]Wu F X,Sun Y L,Xu G H,et al.New saurichthyid fishes(Actinopterygii)from the Middle Triassic(Pelsonian,Anisian)of southwestern China[J].Acta Palaeontologica Polonica,2011,56:581-614.
[35]Sun Z Y,Tintori A,Jiang D Y,et al.A new perleidiform(Actinopterygii,Osteichthyes)from the Middle Anisian(Middle Triassic)of Yunnan,South China[J].Acta Geologica Sinica,2009,83:460-470.
[36]Geng B H,Jin F,Wu F X,et al.New perleidid fishes from the Middle Triassic strata of Yunan Province[J].Geological Bulletin of China,2012,31:915-927.
[37]Lombardo C,Sun Z Y,Tintori A,et al.A new species of the genus Perleidus(Actinopterygii:Perleidiformes)from the Middle Triassic of Southern China[J].Bollettinodella Società Palaeontologica Italiana,2011,50:75-83.
[38]Lin H Q,Sun Z Y,Tintori A,et al.A new species of HabroichthysBrough,1939(Actinopterygii;Peltopleuriformes)from the Pelsonian(Anisian,Middle Triassic)of Yunan Province,South China)[J].Neues Jahrbuchfür Geologie und Paläontologie,Abhandlungen,2011,262:79-89.
[39]López-Arbarello A,Sun Z Y,Sferco E,et al.New species of SangiorgioichthysTintori and Lombardo,2007(Neopterygii,Semionotiformes)from the Anisian of Luoping(Yunnan Province,South China)[J].Zootaxa,2011,2749:25-39.
[40]Xu G H,Wu F X.A deep-bodied ginglymodian fish from the Middle Triassic of eastern Yunnan Province,China,and the phylogeny of lower neopterygians[J].Chinese Science Bulletin,2012,57:111-118.
[41]Tintori A,Sun Z Y,Lombardo C,et al.A new basal neopterygian from the Middle Triassic of Luoping County(South China)[J].RivistaItaliana di Paleontologia e Stratigrafia,2010,116:161-172.
[42]Xu G H,Zhao L J,Michael l C.The oldest ionoscopiform from China sheds new light on the early evolution of halecomorph fishes[J].Biology Letter,2014,10.20140204:1-5.
[43]Xu G H,Shen C C,Zhao L J.Pteronisculusnielseni sp.nov.,a new stem-actinopteran fish from the Middle Triassic of Luoping,Yunnan Province,China[J].Acta Geologica Sinica,2014,52(4):364-380.
[44]Feldmann R M,Schweitzer C E,Hu S X,et al.MacrurousDecapoda from the Luoping Biota(Middle Triassic)of China[J].Journal of Paleontology,2012,86:425-441.
[45]Huang J Y,FeldmannR M,Schweitzer C E,et al.A new shrimp(Decapoda,Dendrobranchiata,Penaeoidea)from the Middle Triassic of Yunnan,Southwest China[J].Journal of Paleontology,2013,87(4):603-611.
[46]Schweitzer C E,Feldmann R M,Hu S X,et al.PenaeoidDecapoda(Dendrobranchiata)from the Luoping Biota(Middle Triassic)of China:systematics and taphonomic framework[J].Journal of Paleontology,2014,88(3):457-474.
[47]黄德志,周长勇,李岩,等.古生物群遗迹化石的三维数字化测量[J].光学与光电技术,2012,10(2):37-41.
[48]谢韬,张启跃,周长勇,等.建立云南罗平生物群地质公园刍议[J].沉积与特提斯地质,2009,29(3):110-112.
[49]谢韬,甘云兰,张启跃,等.罗平生物群国家地质公园地质遗迹特征及其综合评价[J].中国地质,2013,40(6):1959-1967.
Research Achievements and Prospect on the Luoping Biota:according to 1∶50000 Regional Geological Survey and achievement of Specific Study for Luoping, Guishan, Datong, Pengzha, Yunnan
ZHANG Qi-yue, HU Shi-xue, WEN Wen, ZHOU Chang-yong, XIE Tao, HUANG Jin-yuan
(ChengduCenterofChinaGeologicalSurvey,Chengdu,Sichuan610081,China)
Abstract:The Luoping Biota was discovered by the Chengdu Centre of China Geological Survey during geological mapping in 2007 at Luoping County, Yunnan Province, SW China. The intervals containing the fossils belong to the Second Member of the Guanning Formation, Middle Triassic. The Luoping Biota is a Triassic Lagerstätten including both vertebrates and invertebrates.The vertebrates comprise marine reptiles and fish fossils. The invertebrates are mainly consist of arthropods, and associated with bivalves, gastropods, ammonites, echinoderms, lingulid brachiopods, conodonts, foraminifers, and plants. A conodont-based dating indicates that this biota lies within the conodont(Nicoraella kockeli)zone, and belongs to the Pelsonian, Substage of the Anisian Stage, Middle Triassic. The Luoping Biota is preserved within a restricted intra-platform basin. The exceptional preservation of fossils probably resulted from anoxic condition of bottom water and microbial sealing. The Luoping Biota is a window showing the recovery of Triassic marine ecosystem, as well as one of the best localities for Triassic fishes and arthropods fossils. The Luoping Biota provides new information for studying the recovery and evolution of the marine ecosystem after end Permian mass extinction.
Key words:Luoping biota; Triassic; recovery; ecosystem
作者简介:张启跃(1970—)男,研究员,主要从事区域地质矿产调查研究工作。Email:yxzqy@sina.com。
基金项目:中国地质调查“云南1∶5万大同(G48E020009)、圭山(G48E020010)、罗平县(G48E019010)、捧乍(G48E019011)幅区域地质调查项目(编号:1212010610211 )”、“罗平生物区区域地质专项地质调查(编号:12120114068001)”、“罗平生物群及邻区三叠纪化石库专项研究(编号:1212011140051)”和“南盘江—右江成矿区构造演化与矿产空间分布关系调查(编号:12120114030601)”项目联合资助。
收稿日期:2015-01-02; 改回日期: 2015-01-20。
中图分类号:Q91
文献标志码:A
文章编号:2095-8706(2015)02-0024-09
猜你喜欢
现代园艺(2016年7期)2017-01-09
绿色科技(2016年21期)2016-12-27
现代管理科学(2017年1期)2016-12-26
考试周刊(2016年21期)2016-12-16
当代经济管理(2016年11期)2016-12-14
出版广角(2016年14期)2016-12-13
商情(2016年40期)2016-11-28
科教导刊(2016年27期)2016-11-15
科技视界(2016年18期)2016-11-03
科技视界(2016年15期)2016-06-30
