
西双版纳山地雨林乔木层树种20年动态研究
2015-02-06 03:42张高磊李敏敏孟凡荣
生态学报 2015年12期
张高磊, 杜 凡, 王 欢, 李敏敏, 孟凡荣
西南林业大学, 昆明 650224
西双版纳山地雨林乔木层树种20年动态研究
张高磊, 杜 凡*, 王 欢, 李敏敏, 孟凡荣
西南林业大学, 昆明 650224
热带雨林中乔木层树种对群落的结构和功能起着决定性的作用,监测乔木树种的动态,能更好的了解群落的演替。基于西双版纳地区5块共1.25 hm2山地雨林固定样地20年的监测资料,从物种组成、多样性指数、个体数量、主要树种组成和径级结构等方面分析树种动态。研究对象为近成熟的山地雨林,1992—1994年初测时样地中记录到乔木层树种215种,隶属于61科128属;到2012年,乔木层树种增加到239种,隶属于62科148属。分析表明,20年来样地中乔木层树种在科、属、种三级组成水平上均呈缓慢增加趋势,增加率分别为1.64%、15.63%、11.16%。退出乔木层和新进入乔木层的树种主要是偶见种。20年来,样地乔木层总株数从1464株增加至1478株,累计死亡620株,总死亡率29.55%,年均死亡率1.48%;累计进界634株,总进界率30.22%,年均进界率1.51%;群落的多样性指数在小幅度波动中略有增加,Simpson指数、Shannon-Wiener指数和Pielou均匀度指数的增加率分别为0.28%、2.69%、0.55%;乔木层的径级组成未有明显变化,不同径级的死亡和进阶基本处于持平状态。
山地雨林; 乔木层树种; 动态研究
热带森林生物多样性最为丰富却一直面临人类过度利用的重大威胁,因而其研究和保护等相关问题备受关注[1- 3]。国际上对热带森林的研究正在向长期性、大尺度的方向发展,其动态研究逐渐地从空间代替时间的比较研究进入到定位研究[4- 6]。其中以美国热带森林科学研究中心 (Center for Tropical Forest Science, CTFS) 建立的热带森林监测网络最具代表性,自1970年以来,已陆续在世界各地建立了大量的固定监测样地,基于大量的监测数据,在物种共存机制、森林生物多样性的动态变化、树种空间分布格局及其形成机制、采伐与生境片段化的效应等方面得到了丰硕的成果,在生态学领域产生了很大影响[7- 10]。我国从2004年开始实施森林生物多样性监测网络建设,在全国主要气候带内建立监测样地,如吉林长白山温带针阔混交林样地、浙江古田山中亚热带常绿阔叶林样地、广东鼎湖山南亚热带常绿阔叶林样地以及西双版纳热带季节雨林样地。主要关注的科学问题包括植物功能性状与物种共存机制[11- 12]、碳动态与全球变化的关系[13- 15]、植物空间分布格局与生境的关系[16- 18]等。山地雨林是热带山地垂直带上的重要植被类型,在我国主要分布于云南南部和海南岛[19- 20]。之前国内对山地雨林的研究主要集中在群落结构[21- 22]、生态学特征[23- 24]、物种多样性[25- 26]等方面,但由于缺乏长期定位观测的资料,多是根据一次性调查数据或者采用空间代替时间的方法进行,其研究结果具有一定的局限性[27]。目前,虽然国内陆续建立了许多监测样地,但由于监测年限较短,而且缺少对热带山地雨林的监测内容,因此有关我国热带山地雨林的长期定位研究尚属空白。本文基于1992—1994年西南林业大学在西双版纳热带山地雨林建立的固定样地的20年监测数据,探讨山地雨林树种的演变规律,填补这一研究领域的空白。
1 研究地概况
研究地位于西双版纳国家级自然保护区尚勇片区,行政上隶属于勐腊县尚勇镇龙门村,地理位置101°27′—101°41′E,21°10′—22°26′N。本区属低中山浅切割地貌,为北热带湿润季风气候,热量丰富,降水充沛,旱、雨两季分明,年均温度21°C,年均降雨量约1540mm。保护区建立前,因距离村寨较近,林地有轻度的人为干扰,保护区建立后,林下虽然偶尔也存在人为活动,但群落中的树木除了自然枯倒之外,未发现有砍伐等人为破坏的痕迹,并且固定建立样地后,聘请护林员加强监管,样地保持了自然演替状态 (图1)。
图1 固定样地位置示意图
2 研究方法
2.1 野外调查
样地建立于1992—1994年 (表1),样地面积50 m×50 m,分布海拔895—1090 m。以胸径5 cm为乔木的起测径阶,记录乔木树种的胸径、树高、树冠级、冠幅、生活力、物候、坐标等8项因子,用油漆在树干上编号,并在乔木的胸径位置 (1.3 m处) 用油漆画横线标记。以后每隔2—3a复测1次,主要复测胸径,并对样地中胸径新达到5 cm的植株 (进界木) 继续顺序编号,并记录上述8项因子。

表1 山地雨林固定样地情况Table 1 The information of montane rain forest
2.2 数据处理和分析方法
自样地建立以来,初测及复测共计10次。本文采用1992—1994 (初测)、1996年、2001年、2007年和2012年的数据分析树种动态。采用以下多样性指数公式计算:
丰富度指数 (Abundance index)
R=S
(1)
辛普森多样性指数 (Simpson′s Diversity index)[28]
(2)
香农-威钠指数 (Shannon-Wiener index)[28]
(3)
Pielou物种均匀度指数 (Pielou Species Evenness index)[29]
J=H/lnS
(4)
式中,S为样地内总物种数,N为样地内某物种个体总数,Ni为第i种的个体数。
重要值= (相对多度+相对胸高断面积+相对频度)/3
(5)
进界率是累计进界株数除以累计总株数,死亡率是累计死亡株数除以累计总株数。
采用Microsoft Excel 2007、SPSS等软件对数据进行处理。
3 结果与分析
3.1 群落外貌和结构
以2012年复测数据为依据说明群落特征。乔木层盖度70%—80%,主林层高约30—35 m,次林层高约7—15 m;以樟科 (Lauraceae)、壳斗科 (Fagaceae)、大戟科 (Euphorbiaceae) 和茜草科 (Rubiaceae) 为主;树种以披针叶楠 (Phoebelanceolata)、截头石栎 (Lithocarpustruncatus) 为优势,其他有毛银柴 (Aporusavillosa)、湄公栲 (Castanopsismekongensis)、长柄油丹 (Alseodaphnepetiolaris)、木奶果 (Baccaurearamilfora) 等;林内藤本植物发达,主要种类有风车藤 (Hiptagebenghalensis)、大果油麻藤 (Mucunamacrocarpa)、托叶黄檀 (Dalbergiastipulacea) 等;灌木层盖度20%—30%,主要种类为小叶臭黄皮 (Clausenaexcavate)、扇形狗牙花 (Ervatamiaflabelliformis)、猪肚木 (Canthiumhorridum) 等;草本层盖度约50%,主要有全缘赤车 (Pellioniaheyneana)、穿鞘花 (Amischotolypehispida)、刺苞老鼠筋 (Acanthusleucostachyus)、大叶沿阶草 (Ophiopogonlatifolius)、长叶实蕨 (Bolbiteshteroclita)、羽裂鳞毛蕨 (Dryopterisintergriloba) 等;在样地11中,竹芋科的草本植物柊叶 (Phryniumcapitatum) 生长茂盛,盖度可达70%。
3.2 树种组成变化3.2.1 科、属、种变化
5块样地中,初测时共记录到胸径≥5cm的树种215种,隶属于61科128属。20年间,24个树种因个体死亡而在样地中消失,48个树种因胸径增长而进界 (表2)。2012年,样地内共有树种239种,隶属于62科 148属。20年来,样地01树种数从84增加到111种,其中有11个树种消失,有38个树种进入;样地02初测时有72个树种,2012年达97种;样地03和样地04分别增加9种和16种;样地11的树种由82种减少到72种。总的来看,除样地11的树种有所减少外,其余样地树种均有所增加。样地02树种增加最为明显,为36.11%,其次为样地01,为32.14%,样地04和样地03树种的增加率分别为25.00%、18.75%。各样地乔木层树种组成变化较大,但因同一树种在不同样地内有增有减,因此整个群落乔木层树种数变化相对较小,仅增加24种,增加率为11.16%。

表2 1992—2012年的科、属、种变化Table 2 The change of families, genera and species numbers from 1992 to 2012
图2为初测时和2012年复测时乔木层的种序图,可以看出,大多数乔木层树种的个体非常少,只有少数种类的个体较多。初测时,样地乔木层中有129种为偶见种,仅有1—3个个体。20年后,偶见种中有70个种的个体出现死亡,其中49种为部分个体死亡,21种为个体全部死亡而退出乔木层,占退出乔木层总种数 (24种) 的87.50%。2012年,含1—3个个体的偶见种比初测时更多,增加至160种,其中41种是新进入乔木层的树种,占进入乔木层总种数 (48种) 的85.42%。可见,20年来群落乔木层中消失或进入的树种多为个体数量少的偶见种。

图2 初测年和2012年时乔木层的种序图比较
3.2.2 树种多样性指数

图3 树种多样性指数
20年间,5块样地的乔木层树种丰富度在215—252之间波动。乔木层树种的阶段性变化情况是:由1992年的215种增加到1996年218种,再增加到2001年的252种,达到20年间的最高值;2007年减少为219种;2012年又升到到239种。多样性指数的变化幅度较小,Simpson指数1992年为0.9696,2012年为0.9723;Shannon-Wiener指数从4.3238增加到4.4401,Pielou均匀度指数从0.8051增加到0.8095,增加率分别为0.28%、2.69%和0.55%(图3)。
3.3 树种数量变化3.3.1 乔木层株数
树木死亡是调节群落组成和结构的重要方式。20年间,5个样地中胸径≥5 cm的乔木植株由初测时1464株增加至2012年1478株,净增14株(表3),增加率极小,仅0.96%。有142个树种的个体不同程度进界,累计进界634株,进界率30.22%,年均进界率1.51%;有138个树种的个体不同程度死亡,初测个体死亡526株,进界后出现死亡94株,共累计死亡620株,总死亡率为29.55%,年均死亡率1.48%。5个样地中,年均死亡率最大的是样地11,达2.06%,其次是样地01,为1.54%,样地03和样地02的年死亡率稍低,分别为1.51%、1.45%,样地04的年死亡率最低,为1.19%。年均进界率最大的是样地02,为2.23%,其余依次为样地01、样地04、样地03和样地11,年均进界率分别为2.20%、1.65%、0.98%和0.96%。其中,样地11的年均死亡率最高而年均进界率最低,这与其生境特点有着密不可分的关系。样地11林下草本植物盖度高达80%以上,其中仅柊叶盖度就达到70%,高约2m,使得乔木树种的种子难以接触到土壤,即使少量接触土壤萌发后的幼苗也往往因光照严重不足而难以成活,密闭的林下环境对乔木幼树的更新和生长造成了很大的阻碍。其他样地草本层盖度较小,约40%—60%,对幼树的更新影响较小,所以每次复测的进界个体相对较多。
3.3.2 主要树种的株数变化
森林群落的乔木,尤其是优势种或建群种对构建群落特征起着决定性作用,研究它们的变化,能更好的反映出森林的演变动态[30]。本文以重要值来综合衡量树种在群落中的地位和作用。计算表明排名前26位树种的重要值累计达到50%。其中,排名前10位的树种不仅重要值大,且个体较多或径级大,对群落的性质和结构特征起关键作用,所以选择重要值较大的前10位作为主要树种加以分析。
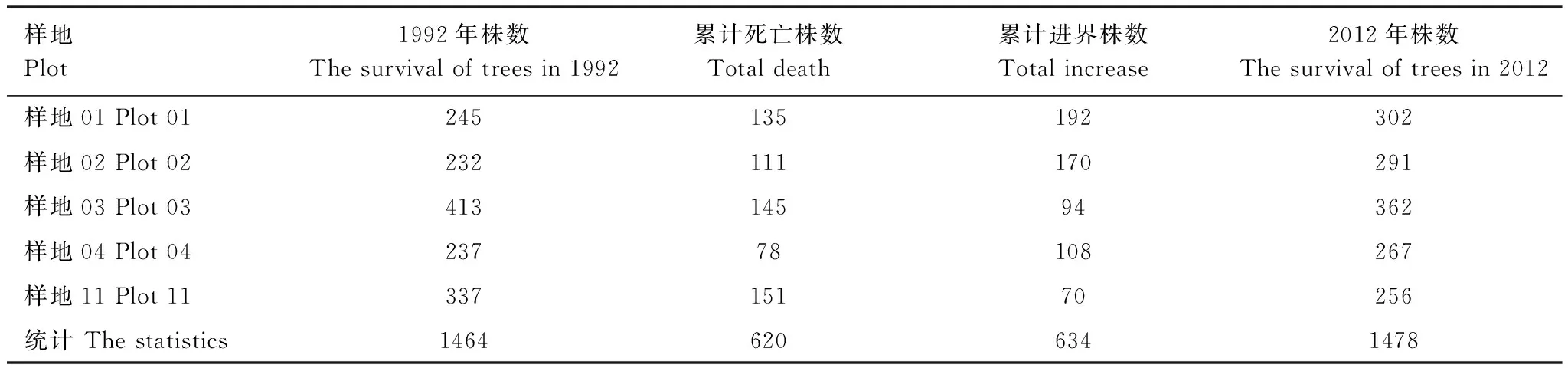
表3 1992—2012年间样地乔木个体变化情况Table 3 The change of number of individual trees from 1992 to 2012
从表4可以看出:20年来主要树种的株数从616株减少到575株,重要值之和从32.21%下降到29.64%,株数和重要值均呈现下降趋势。初测时的主要树种在2012年均存在,除浆果乌桕被肉实树 (Sarcospermaarboreum) 替代外,其余树种仍处于优势地位,但优势度发生了变化。披针叶楠死亡54株,进界62株,总株数变化不大,大量的进界个体导致平均胸径减少,但仍然是群落里最优势的树种。截头石栎个体从94株减少到58株,20年间仅有1株 (2007年) 进界,年死亡率从3.19%上升到2012年的19.44%,群落重要值从5.19下降到3.64,衰退速度加快。主林层的浆果乌桕和次林层的毛银柴,因死亡率较高而进界率持续降低,导致优势度明显下降。次林层中,披针叶楠的重要值从5.36下降到4.63;毛银柴在初测时是次林层中的第二优势种,也因为死亡率较高和进界率低,优势度明显下降。同样位于次林层的木奶果,从65株增加到92株,重要值从2.22增加到3.03。到2012年,木奶果的优势度高于毛银柴,成为第二优势种。相对于其他主要种来说,长柄油丹、红木荷的死亡和进界的株数均很少,但因胸径增加而使其优势度上升,重要值分别从3.14、2.50上升到3.34、3.02,逐渐成为群落的建群种。
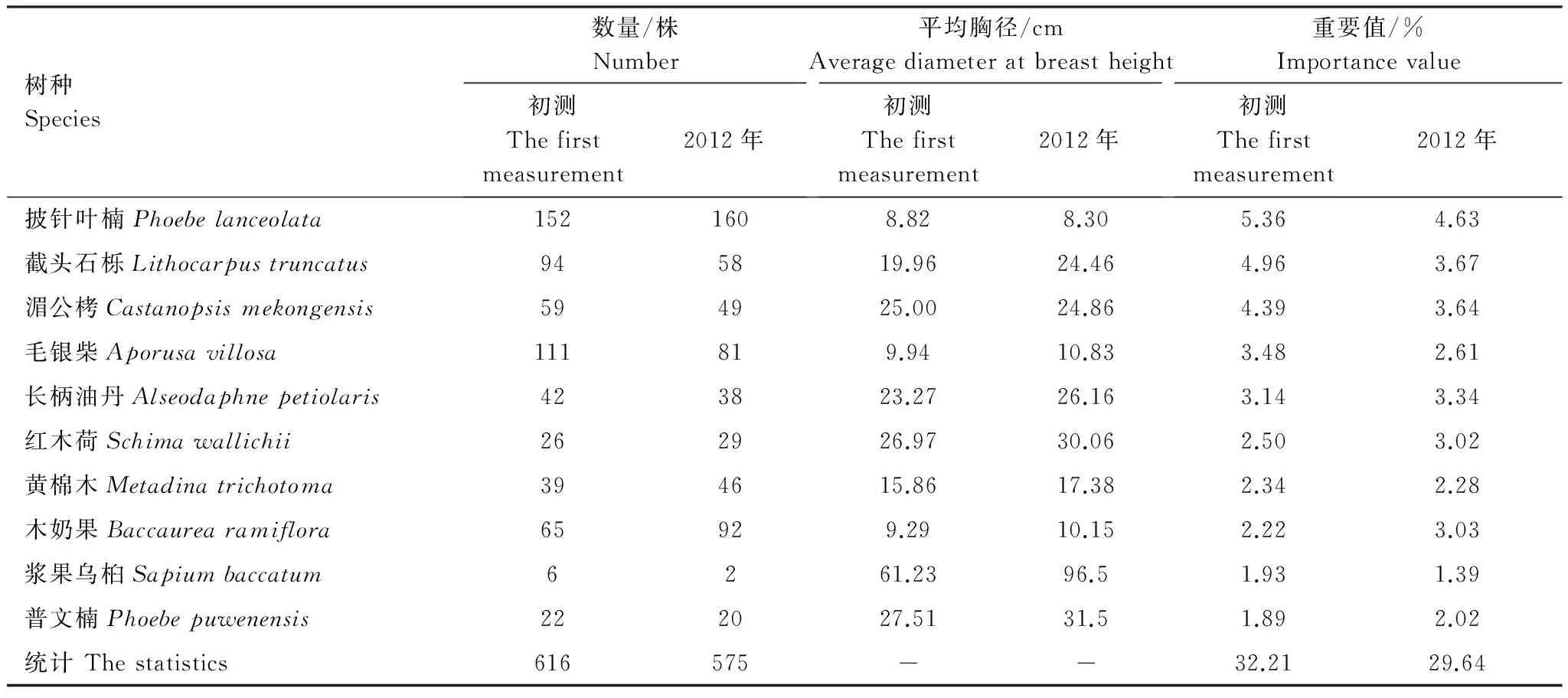
表4 主要树种变化Table 4 The change of the main tree species
3.4 径级结构
径级结构是反映群落结构的重要指标。本文以10 cm为区间划分径级,将样地内的乔木划分为7个径级,即<10 cm、10—20 cm、20—30 cm、30—40 cm、40—50 cm、50—60 cm和≥60 cm。各样地乔木植株的平均胸径在19.16—24.51 cm之间,小、中径级植株 (5≤DBH<60 cm) 比例占91.75%—98.90%。图4是初测和2012年复测的径级分布图,可以看出,样地以小径级的植株 (DBH<20 cm) 数量最多,占77.27%—77.32%;中径级 (20 cm≤DBH<60 cm) 占20.77%—21.04%;大径级 (DBH≥60 cm) 乔木很少,占1.69%—1.91%,胸径最大者为样地04中浆果乌桕 (Sapiumbaccatum),达138.49 cm。20年来,各径级的死亡株数分别为354、191、52、13、7、6和6;存活个体随着时间变化,其胸径也在增加,各径级的增加株数分别为355、200、29、26、21、9和3。7个径级中,0—10 cm径级的死亡率最大,为32.78%;其次是10—20 cm径级,为30.76%;死亡率最低的是30—40 cm径级,为12.15%;50—60 cm径级中,截头石栎的个体死亡较多,使得该径级的死亡率有所上升,达到21.43%。各径级由于死亡株数和径级基数的不同,径级死亡率也表现出差别,不同径级的死亡率随胸径增加大体呈“V”型分布 (图5)。20年来,初测1464株个体中有526株死亡,占原有株数的35.93%,尽管样地中的植株个体发生了较大变化,但由于进阶个体的补充,因此各径级的数量结构总体上变化甚微。从径级株数看,中小径级的株数多,随着径级的增加,株数急剧减少,到50 cm径级以后,株数逐渐趋于平稳。这种金字塔形的径级结构保证了群落演替发展,足够多的小径级个体能够维持群落乔木层的自我更新,适量密度的大径级个体保证群落的稳定性[18,31-32]。
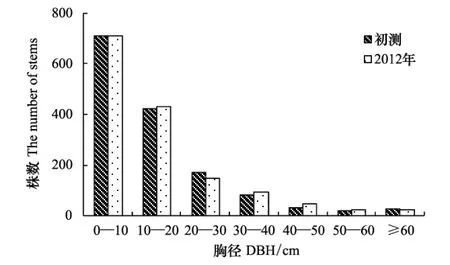
图4 乔木植株径级频度分布
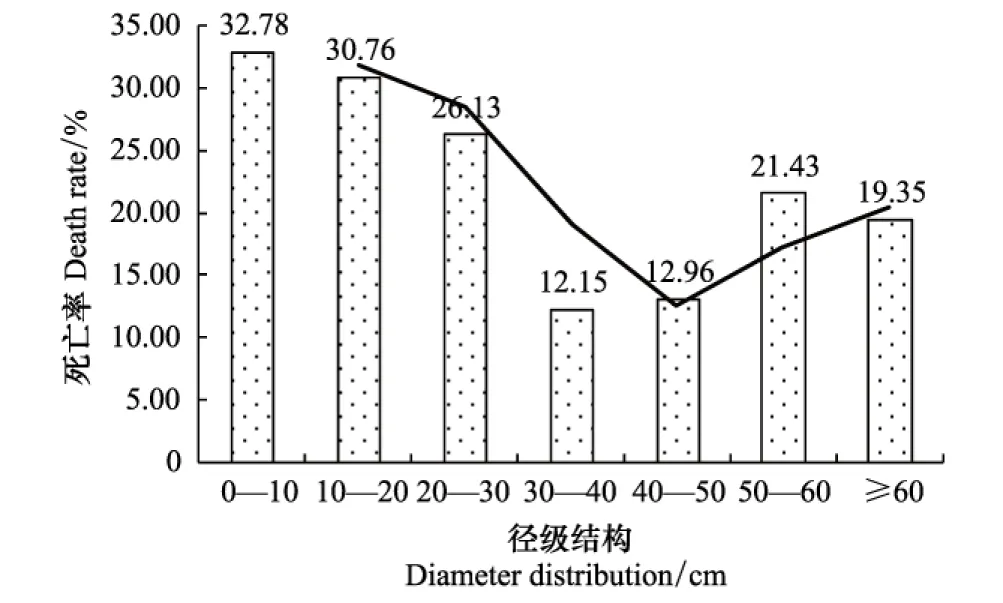
图5 不同径级结构的死亡率
4 讨论与结论
在植被群落的动态中,物种组成和多样性的变化是最值得关注的。在热带雨林群落中,大多数乔木树种的个体是非常少的,只有少数种类有较多个体[33]。在演替过程中,个体少的树种一旦出现死亡就较易导致它们所属的科属消失。通过初测的历史记录与现今的调查资料比较,24种树种从该群落乔木层中消失,48种树种进入乔木层,3.39%科、4.69%的属和10.23%的种类被后来的成分替换。在这些消失或进入乔木层的树种中,绝大多数属于偶见种。研究地相对靠近村寨和林缘,容易受到外界环境干扰和边缘效应的影响,因此出现偶见种的频度较高。
20年间,乔木层多数树种的个体均有不同程度死亡和进界,年均死亡率1.48%,年均进界率1.51%。各样地的死亡率在1.19%—2.06%之间变动,进界率在0.96%—2.26%之间变动。国外研究表明[34- 35],在正常情况下,热带雨林顶级群落的乔木层年死亡率为0.9%—1.1%。本研究地的死亡率均高于1.1%,显示群落更新速率较快。一方面可能是由于群落未达到顶级阶段,其乔木更新相对较快;另外一方面,样地所在地形较为陡峻,坡度均在20°以上,而本区雨季降雨量大而集中,使得上层大径级乔木较容易翻根倒伏,并进而压断压倒其下层的其他乔木,由此产生的林窗也为阳性成分提供了生境,因而本区山地雨林的死亡率和进界率也高于一般的热带雨林。
通过对比分析,优势树种中除浆果乌桕被肉实树替代外,其余树种并未退出优势树种之列,仅仅是树种间的优势度排序发生了一定的变化。调查表明,截头石栎、湄公栲、毛银柴等树种的种子在林缘、林窗条件下更新较好,属于阳性倾向树种;披针叶楠、木奶果、肉实树等树种在适宜郁闭的林下,种子萌发和生长更具优势,属于阴性倾向树种。研究地在保护区建立之前,由于受到部分人为干扰,阳性成分在林窗中更新并成长起来,但原来雨林成分的幼、小树并未大量死亡。随着林窗的消失,阳性成分也随之滞后消失,当其他林窗出现,阳性成分又会随之短暂出现,这是热带雨林维持生物多样性的机制之一。20年来,这些山地雨林原生成分慢慢的恢复并成长起来,但尚未恢复到顶级状态,披针叶楠、截头石栎、湄公栲等仍然具有较大的相对多度和优势度,该群落的整体性质未发生大的改变。但是具有阳性倾向树种的优势度明显降低,阴性树种的优势度在上升,表明群落向着山地雨林顶级阶段演替。
西双版纳山地雨林是分布于较高海拔的热带山地下的雨林类型,物种组成十分丰富,群落演替过程更是复杂多变。本文的研究反映了20年来该区山地雨林乔木层树种的动态变化过程,研究表明所监测的山地雨林正在缓慢地向顶级群落阶段演替,但是由于监测的时间跨度仍显不足,目前尚难于预测此类山地雨林何时能够演替到最为顶级的群落阶段,所以,今后进行更加长久的观测研究仍然是需要的。
致谢:感谢提供数据的单位和个人。
[1] Schnitzer S A, Carson W P. Treefall gaps and the maintenance of species diversity in a tropical forest. Ecology, 2001, 82(4): 913- 919.
[2] Potts M D. Drought in a Bomean everwet rain forest. Journal of Ecology, 2003, 91: 467- 474.
[3] Wright S J. Plant diversity in tropical forest: a review of mechanism of species coexistence. Oecologia, 2002, 130: 1- 14.
[4] Condit R, Hubbell S, Lafrnkie J V, Sukumar R, Manokaran N, Foster R B, Ashton P S. Species-area and species-individual relationships for tropical trees: a comparison of three 50-ha plots. Journal of Ecology, 1996, 84(4): 549- 562.
[5] Lieberman D, Lieberman M, Peralta R, Hartshorn G S. Tropical forest structure and composition on a largescale altitudinal gradient in Costa Rica. Journal of Ecology, 1996, 84(2): 137- 152.
[6] Harms K E, Hubbell S P, Condit R, Foster R B. Habitat association of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology, 2001, 89(6): 947- 959.
[7] Condit R. Research in large, long-term tropical forest plot. Trends in Ecology and Evolution, 1995, 10(1): 18- 23.
[8] Hubbell S P, Foster R B, Brien S T O, Harms K E, Condit R, Wechsler B, Wright S J, Lao S L. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science, 1999, 283(5401): 554- 557.
[9] Hooper E R, Legendre P, Condit R. Factors affecting community composition of forest regeneration in deforested, abandoned land in Panama. Ecology, 2004, 85(12): 3313- 3326.
[10] Hubbell S P. Neutral theory and the evolution of ecological equivalence. Ecology, 2006, 87(60): 1387- 1398.
[11] 戚裕锋, 杨徐烽, 张奇平, 宋坤, 康敏明, 达良俊. 浙江天童受损常绿阔叶林实验生态学研究 (V): 不同干扰下植被恢复初期主要树种五年的恢复和更新. 华东师范大学学报: 自然科学版, 2010, (3): 10- 25.
[12] 康敏明. 浙江天童受损常绿阔叶林恢复动态及主要树种适应策略 [D]. 上海: 华东师范大学, 2010.
[13] 莫江明, 方运霆, 彭少麟, Sandra B, 周国逸. 鼎湖山南亚热带常绿阔叶林碳素积累和分配特征. 生态学报, 2003, 23(10): 1970- 1976.
[14] 李冬. 西双版纳季风常绿阔叶林的碳贮量及其分配特征研究 [D]. 勐腊: 中国科学院研究生院 (西双版纳热带植物园), 2006.
[15] 吕晓涛. 西双版纳热带季节雨林碳储量的研究 [D]. 勐腊:中国科学院研究生院 (西双版纳热带植物园), 2006.
[16] 刘文平, 曹洪麟, 刘卫, 练琚愉, 吴林芳. 鼎湖山季风常绿阔叶林不同生境物种多样性研究. 安徽农业科学, 2011, 39(26): 16159- 16163.
[17] 方运霆, 莫江明, 周国逸, 张德强, 薛璟花. 鼎湖山南亚热带常绿阔叶林植物和土壤微量元素含量. 广西植物, 2005, 25(6): 504- 510.
[18] 兰国玉, 胡跃华, 曹敏, 朱华, 王洪, 周仕顺, 邓晓保, 崔景云, 黄建国, 刘林云, 许海龙, 宋军平, 何有才. 西双版纳热带森林动态监测样地——树种组成与空间分布格局. 植物生态学报, 2008, 32(2): 287- 298.
[19] Grubb P J, Lloyd J R, Pennington T D. A comparison of montane and lowland rain forest in Ecuador. The forest structure, physiognomy and floristics. Journal of Ecology, 1963, 51(3): 567- 601.
[20] Whitmore T C. Tropical Rain Forest of the far East. 2nd ed. Oxford: Clarendon Press, 1984: 352- 352.
[21] 曾觉民. 西双版纳普文的山地雨林及其生态演替. 云南林业科技, 2002, (4): 11- 16.
[22] 臧润国, 蒋有绪, 余世孝. 海南霸王岭热带山地雨林森林循环与树种多样性动态. 生态学报, 2002, 22(1): 24- 32.
[23] 张国成, 施济普, 周仕顺, 朱华. 西双版纳勐养山地雨林群落生态学研究. 应用与环境生物学报, 2006, 12(6): 761- 765.
[24] 朱华, 王洪, 李保贵. 滇南勐宋热带山地雨林的物种多样性与生态学特征. 植物生态学报, 2004, 28(3): 351- 360.
[25] 李宗善, 唐建维, 郑征, 李庆军, 罗成昆, 刘正安, 李自能, 段文勇, 郭贤明. 西双版纳热带山地雨林的植物多样性研究. 植物生态学报, 2004, 28(6): 833- 843.
[26] 王峥峰, 安树青, Campell D G, 杨小波, 朱学雷. 海南岛吊罗山山地雨林物种多样性. 生态学报, 1999, 19(1): 61- 67.
[27] 朱华, 许再富, 王洪, 李保贵. 西双版纳片断热带雨林30多年来植物种类组成及种群结构的变化. 云南植物研究, 2001, 23(4): 415- 427.
[28] Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana, IL: University of Illinois Press, 1949.
[29] Pielou E C. The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 1966, 13: 131- 144.
[30] Ni J. Plant functional types and biomes of China at a regional scale. Acta Botanica Sinica, 2001, 43(4): 219- 425.
[31] 孙伟中, 代力民, 章一平. 椴树阔叶红松林群落主要树种径级结构研究. 生态学杂志, 1997, 16(1): 19- 22.
[32] Karl J N, Jeremy J M, Richard H R. Tree size frequency distributions, plant density, age and community disturbance. Ecology Letters, 2003, 6(5): 405- 411.
[33] Cao M, Zhang J H. Tree species diversity of tropical forest vegetation in Xishuangbanna, SW China. Biodiversity and Conservation, 1997, 6(7): 995- 1006.
[34] Nakagawa M, Tanaka K, Nakashizuka T, Ohkubo T, Maeda T, Sato K, Miguchi H, Nagamasu H, Ogino K, Teo S, Hamid A A, Seng L H. Impact of severe drought associated with the 1997- 1998 El Nino in a tropical forest in Sarawak. Journal of Tropical Ecology, 2000, 16(3): 355- 367.
[35] Williamson G B, Laurance W F, Oliveira A A, Delamonica P, Gascon C, Lovejoy T E, Pohl L. Amazonian tree mortality during the 1997 El Nino drought. Conservation Biology, 2000, 14(5): 1538- 1542.
Study on tree layer dynamic in Xishuangbanna montane rain forest based on 20 years′ monitoring
ZHANG Gaolei, DU Fan*, WANG Huan, LI Minmin, MENG Fanrong
SouthwestForestryUniversity,Kunming650224,China
Based on 20 years′ monitoring in montane rain forest plots (1.25hm2within 5 blocks) in Xishuangbanna, we analyzed the dynamics of tree layer in terms of species composition, diversity index, changes of individual tree numbers, changes of dominant tree species, and diameter distribution. The results showed that in 20 years, the tree species numbers were increased from 215 (in 61 families and 128 genera) to 239 (in 62 families and 148 genera). The numbers of families, genera and species were slowly increased at the rates of 1.64%, 15.63% and 11.16%. Those entering into or withdrawing from the tree layer belonged to accidental species with less individuals. The total individual numbers of trees were increased from 1464 to 1478 while 620 were died with a mortality rate at 29.55%, the average annual mortality rate at 1.48%. Meanwhile, 634 trees entered into the tree layer and the total in growth rate was 30.22%, the average annual growth rate at 1.51%. The diversity index showed a slight fluctuation, with increasing rates of Simpson index, Shannon-Wiener index and Pielou′s index at 0.28%, 2.69% and 0.55%, respectively. The death and advanced of different diameter classes were basically at flat state.
mountain rainforest; tree layer; dynamic research
西南林业大学植物学重点学科建设项目
2013- 11- 08;
2014- 06- 12
10.5846/stxb201311082694
*通讯作者Corresponding author.E-mail: kmdufan@163.com
张高磊, 杜凡, 王欢, 李敏敏, 孟凡荣.西双版纳山地雨林乔木层树种20年动态研究.生态学报,2015,35(12):4053- 4062.
Zhang G L, Du F, Wang H, Li M M, Meng F R.Study on tree layer dynamic in Xishuangbanna montane rain forest based on 20 years′ monitoring.Acta Ecologica Sinica,2015,35(12):4053- 4062.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
广西林业科学(2021年6期)2022-01-21
小哥白尼(神奇星球)(2020年9期)2021-01-18
小哥白尼(神奇星球)(2020年9期)2021-01-18
临床检验杂志(电子版)(2020年1期)2020-04-03
少儿美术(快乐历史地理)(2019年9期)2020-01-18
小哥白尼(趣味科学)(2019年7期)2019-11-16
新农民(2019年9期)2019-02-19
学苑创造·B版(2015年12期)2016-06-23
