
5个温带树种冠层枝叶非结构性碳水化合物浓度的空间变异
2015-01-19 07:06:38张海燕王传宽王兴昌
生态学报 2015年19期
张海燕, 王传宽, 王兴昌
东北林业大学生态研究中心, 哈尔滨 150040
5个温带树种冠层枝叶非结构性碳水化合物浓度的空间变异
张海燕, 王传宽*, 王兴昌
东北林业大学生态研究中心, 哈尔滨 150040
非结构性碳水化合物(NSC)是树木存活和生长的重要能源物质。冠层NSC不但是全树NSC的来源,也是全树NSC的重要储存库。然而,冠层NSC空间变异的研究较少,因而影响了树木NSC分配的估算精度。以红松(Pinuskoraiensis)、兴安落叶松(Larixgmelinii)、水曲柳(Fraxinusmandshurica)、蒙古栎(Quercusmongolica)和白桦(Betulaplatyphylla)5个温带树种为研究对象,测定了不同高度冠层叶和细枝(直径≤ 3 cm)NSC浓度,分析了粗枝(一级侧枝)枝皮、边材和心材NSC浓度轴向变化及其与枝径的关系。结果表明:(1)除了5月末兴安落叶松树冠中层叶淀粉浓度显著高于树冠下层,以及8月中旬树冠上层叶可溶性糖浓度显著高于树冠中层之外,其他树种冠层叶NSC浓度的垂直变化不显著。常绿树种红松叶龄对NSC浓度的影响在生长季中期显著,但在生长季末期和休眠季节的影响不显著。(2)除了5月末红松树冠上层细枝可溶性糖浓度显著高于树冠中层之外,其他树种不同高度冠层间细枝NSC浓度差异不显著。(3)在纵向上,阔叶树种蒙古栎、水曲柳和白桦粗枝的枝皮、边材和心材NSC浓度多随着距树枝基部距离的增加而升高;在径向上,NSC浓度(除了水曲柳边材淀粉和白桦枝皮淀粉之外)多随着枝径增加而降低,表明树枝中的NSC浓度随着远离碳源而降低。总体上,5个温带树种冠层叶、细枝NSC浓度的空间变异不显著,但枝径和叶龄对NSC浓度的影响因树种、组织和季节而异,这在未来研究中应予考虑。
非结构性碳水化合物(NSC); 叶; 枝; 空间变异; 温带森林
非结构性碳水化合物(NSC)是植物光合作用的主要产物[1],是光合作用(碳吸收)与生长利用(碳消耗)之间主要的中间贮存物,也是植物生长和代谢的重要物质[2]。在温带和北方森林生态系统中,树木春季萌发生长[3]、冬季维持呼吸[4]、缓解水分胁迫[5-6]以及刈割后的萌蘖[7]等都与植物贮藏碳水化合物的水平密切相关,这表明多年生植物碳水化合物的贮藏特性反映了其对环境的适应策略[8-9]。全球变化背景下,树木干旱死亡机理的碳饥饿假说[6,10]和树木生长的碳限制假说[11]的提出及其验证研究,将植物水力特征、植物碳收支和植物病理领域紧密联系起来[12]。然而,NSC存储与生长和存活的关系尚存在争议[13-14],亟需更多的数据阐明NSC在树木中的利用和分配[15]。
树木的NSC浓度因树体高大、形态各异而呈现出明显的时空异质性[16-18],量化树木NSC库存在很大的困难[12,15]。由于成年树木冠层高大,难以直接测定整个冠层NSC,通常利用某一径级树枝的NSC测定值推算整个冠层树枝的NSC。阐明树木NSC的空间变化规律,不仅可以了解树木生理特征随树木发育的变化过程[19-21],还可以显著降低树木NSC含量估算的不确定性[18]。
树冠既是树木光合作用的场所,也是树木NSC的主要储存组织之一。以往冠层NSC研究主要集中在季节动态方面[22-26],冠层NSC空间变异的研究不但较少,而且研究结果不一[18,20-21]。通常树冠上层接受光照较强,其光合能力较高[27],因而上层叶片可同化更多的CO2,产生更多的NSC,其NSC浓度也可能高于下层叶片。然而,瑞士五针松(Pinuscembra)[19]、长苞冷杉(Abiesgeorgei)和方枝柏(Juniperussaltuaria)[20]以及红松(Pinuskoraiensis)[21]不同高度冠层间叶NSC浓度差异不显著。另一方面,叶龄对这4种树种的叶NSC浓度影响显著[19-21]。树枝NSC的空间变异研究较少。北美黄杉(Pseudotsugamenziesii)[16]和北美黄松(Ponderosapine)[28]枝条NSC浓度随着高度增加而升高,但欧洲山毛榉(Fagussylvatica)和无梗花栎(Quercuspetraea)[18]不同高度冠层间树枝NSC浓度差异不显著;此外,树枝NSC浓度却随着枝径和枝龄的增加而降低[18]。这些观点和矛盾之处尚需更多的研究结果加以检验。
尽管我国温带树种NSC研究不少[17,21,26,29-33],但尚未涉及不同特性树种冠层枝叶NSC时空变异的综合测定。本研究以中国东北东部温带森林5个树种为对象,树种包括:红松、兴安落叶松(Larixgmelinii)、水曲柳(Fraxinusmandshurica)、蒙古栎(Quercusmongolica)和白桦(Betulaplatyphylla);它们涵盖了不同的叶性状(常绿和落叶)和生活型(针叶和阔叶)。本文研究目的:(1)分析5个树种不同高度冠层间叶和细枝NSC浓度是否存在显著差异,常绿针叶NSC是否受叶龄影响;(2)研究3个阔叶树种粗枝3种组织(枝皮、边材和心材)NSC浓度的轴向(与枝基的距离)变化;(3)探索落叶树种粗枝NSC浓度与枝径的关系。
1 材料与方法
1.1 研究地概况
研究地位于黑龙江省尚志市境内的帽儿山森林生态系统国家野外科学观测研究站(45.40° N,127.66° E),平均海拔400 m,平均坡度10°—15°,地带性土壤为暗棕色森林土。属于大陆性季风气候, 年平均气温3.1 ℃,1月平均气温-18.5 ℃,7月平均气温22.0 ℃;年降水量629 mm,主要集中在6—8月;年蒸发量864 mm;无霜期为120—140 d。现有植被是原地带性植被——阔叶红松林遭受人为干扰后演替成的天然次生林和人工林,代表东北东部山区典型的森林类型。
1.2 试验方法
所有样木均选自典型森林类型的永久样地(表1)附近。各林分类型林龄40—70a,胸高断面积30—38 m2/hm2之间[34]。
叶和细枝采样:为了尽可能减小光照条件对树木NSC的影响,每个树种选取胸径相近的健康优势木各3株(表1),于2010年4—5月参照以往观测的叶物候规律[35]对5个树种的枝叶分冠层采样。采样时间分别为:4月中旬(发芽前)、5月中旬(展叶较早的树种开始发芽)、5月末(展叶较晚的树种开始发芽)。为了分析季节变化是否影响冠层叶、细枝NSC浓度的垂直变化以及叶龄对NSC的影响,分别对红松(生长季中后期仅冠中)和兴安落叶松(3个冠层高度)按照物候采样7次[26]。从树冠南向的上层、中层和下层3个高度用高枝剪随机选取树枝,采取叶和细枝(带皮直径≤3 cm,不含当年生枝)作为样品。
粗枝采样:由于红松和兴安落叶松为单轴分枝树种,其树枝较短且枝径较小,因此只研究3个阔叶树种(水曲柳、蒙古栎和白桦)粗枝NSC浓度的轴向变化。为了避免样品处理期间的NSC损失,在2010年10月树木进入休眠期后对3个树种的粗枝(一级侧枝)采样。从每个树种的3株优势木中随机选取1株,冠层中下部选取一个粗大的一级侧枝并从基部锯下,从枝基部(紧挨着树干部位)开始,每隔1 m或2 m截取一5—10 cm厚圆盘直至枝梢。每个枝盘分为枝皮、边材和心材3种组织(白桦不能区分心材[32])作为样品,置于冷藏箱中(0—4 ℃)保存;带回实验室后微波炉高温(600 W)杀青90 s,在65 ℃下烘干至恒重,粉碎以作NSC分析。
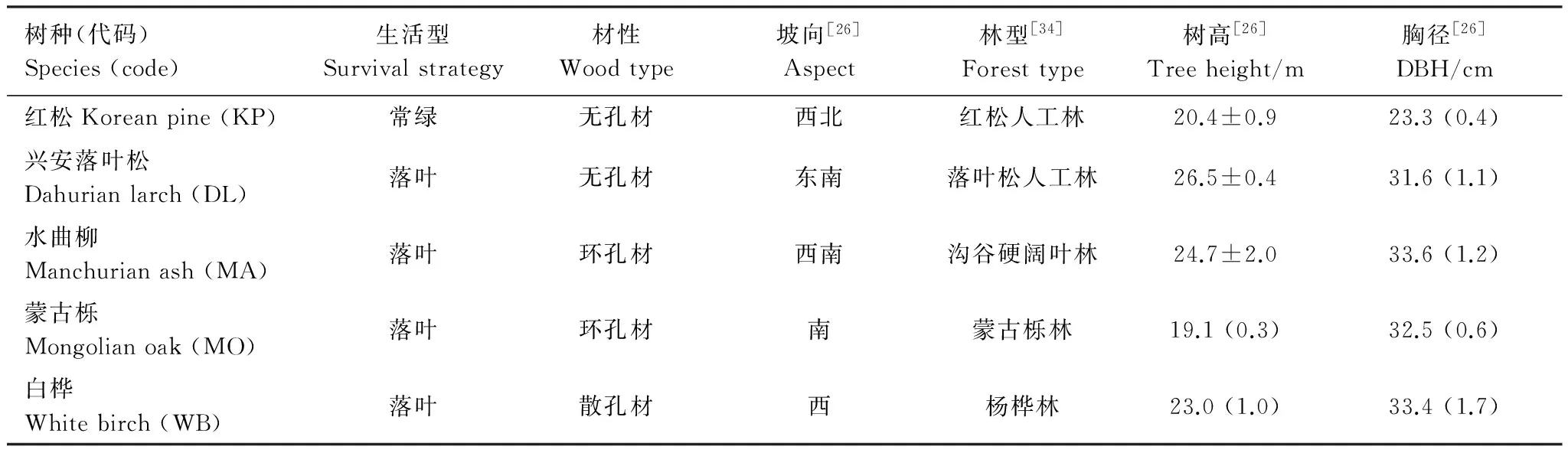
表1 样木基本特征(平均值±标准误,n=3)
1.3 项目测定
可溶性糖和淀粉浓度均采用改进的苯酚硫酸法[36]测定,具体实验方法参照于丽敏等[17]。NSC浓度用百分比干重(% DM)表示。总非结构性碳水化合物(TNC)为可溶性糖与淀粉的总和;NSC泛指可溶性糖、淀粉和TNC[26]。
1.4 数据分析
采用单因素方差分析(ANOVA)比较每个树种冠层高度间NSC浓度的差异和不同叶龄红松针叶NSC浓度的差异,显著性水平设定为α=0.05。粗枝NSC采用线性回归检验其随着枝径的变化趋势。所有数据均采用SPSS 16.0软件进行分析,SigmaPlot 10.0软件作图。
2 结果
2.1 不同冠层叶NSC浓度及其与叶龄的关系
除了5月末(生长季初)兴安落叶松冠层中部叶淀粉浓度显著高于冠层下部之外(P=0.043),其他树种不同高度冠层间叶NSC浓度差异不显著(图1)。5月末5个树种叶NSC浓度相比,白桦可溶性糖浓度显著高于其他树种,蒙古栎最低;红松淀粉浓度显著高于其他树种,落叶松、水曲柳和蒙古栎最低;红松TNC浓度显著高于其他树种,蒙古栎最低(图1)。
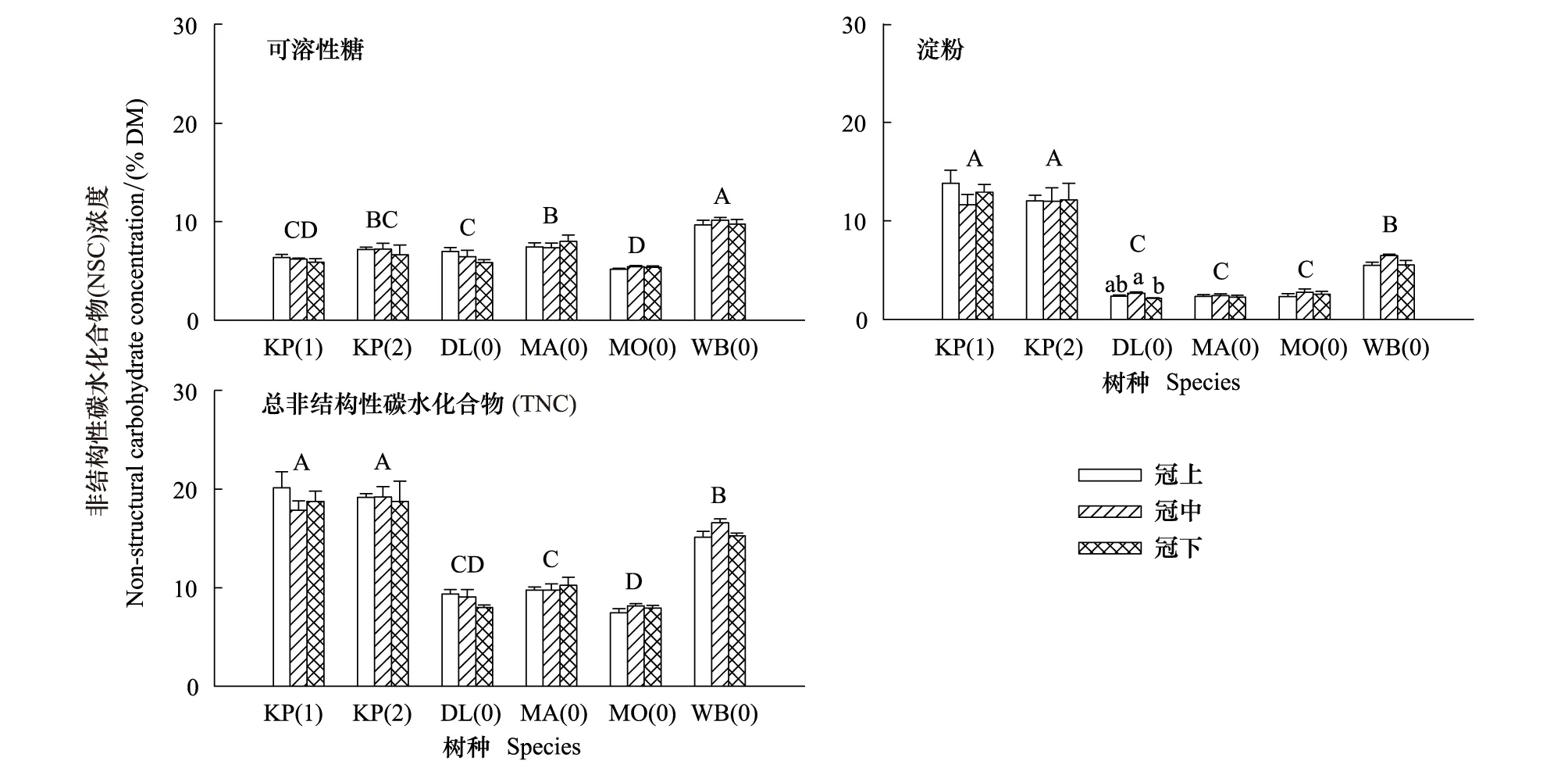
图1 5月末5个树种冠层叶非结构性碳水化合物(NSC)浓度的垂直变化(平均值±标准误,n=3)Fig.1 Vertical variation in non-structural carbohydrates (NSC) concentrations in foliages for the five tree species in late May (mean±se,n=3)TNC: total non-structural carbohydrates; KP(1)和KP(2)分别表示红松1年生和多年生针叶,DL(0)、MA(0)、MO(0)和WB(0)分别表示兴安落叶松、水曲柳、蒙古栎和白桦叶; 误差线上方不同小写和大写字母分别表示冠层间和种间显著差异组别(P < 0.05)
以树体最高的兴安落叶松为例,在4—10月连续7次采样(4月、5月初和10月没有叶)检验冠层高度间NSC浓度的差异是否具有季节动态。结果表明,除了5月末冠中叶淀粉浓度显著高于冠下和8月中旬冠上叶可溶性糖浓度显著高于冠中(P= 0.034)之外,其余月份冠层高度间叶NSC浓度差异不显著(图2)。
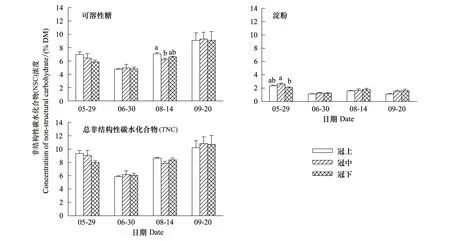
图2 生长季兴安落叶松冠层叶非结构性碳水化合物(NSC)浓度的垂直变化(平均值±标准误,n=3)Fig.2 Vertical variations in non-structural carbohydrates (NSC) concentrations in foliage for Dahurian larch along the growing season (The vertical bars indicate standard errors) (mean+se,n=3)误差线上方不同字母表示显著差异组别(P < 0.05)
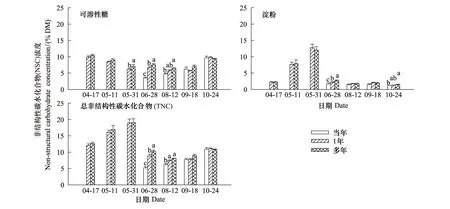
图3 红松针叶非结构性碳水化合物(NSC)浓度的叶龄效应(平均值±标准误,n=3)Fig.3 Needle-age related effects on non-structural carbohydrates (NSC) concentrations for Korean pine (mean±se,n=3)误差线上方不同字母表示显著差异组别(P < 0.05)
叶龄显著影响红松针叶5月末、6月和8月的可溶性糖浓度、6月和10月的淀粉浓度、6月和8月的TNC浓度(图3)。总体而言,NSC浓度随着叶龄增加而升高。叶龄对红松针叶NSC浓度的影响主要表现在生长季中期,之后随着当年生叶逐渐成熟,叶龄的影响逐渐减小。5月末红松多年和1年生叶中淀粉居多,而此时落叶树种当年生叶中可溶性糖居多(图1),但其余月份红松针叶则以可溶性糖为主(图3)。
2.2 不同冠层细枝NSC浓度
除了红松冠上细枝可溶性糖浓度显著高于冠中之外(P=0.046),4—5月其他树种冠层高度间细枝的可溶性糖、淀粉和TNC浓度差异不显著(图4)。冠层间细枝NSC浓度无一致的变化规律,仅水曲柳和白桦在4月、兴安落叶松在5月末的可溶性糖和TNC浓度随着冠层部位的上升呈递增趋势,但5月中旬水曲柳和蒙古栎细枝NSC浓度表现出与之相反的变化趋势(图4)。兴安落叶松连续7次采样(4—10月)表明,冠层高度间细枝NSC浓度差异均不显著(P> 0.05)。
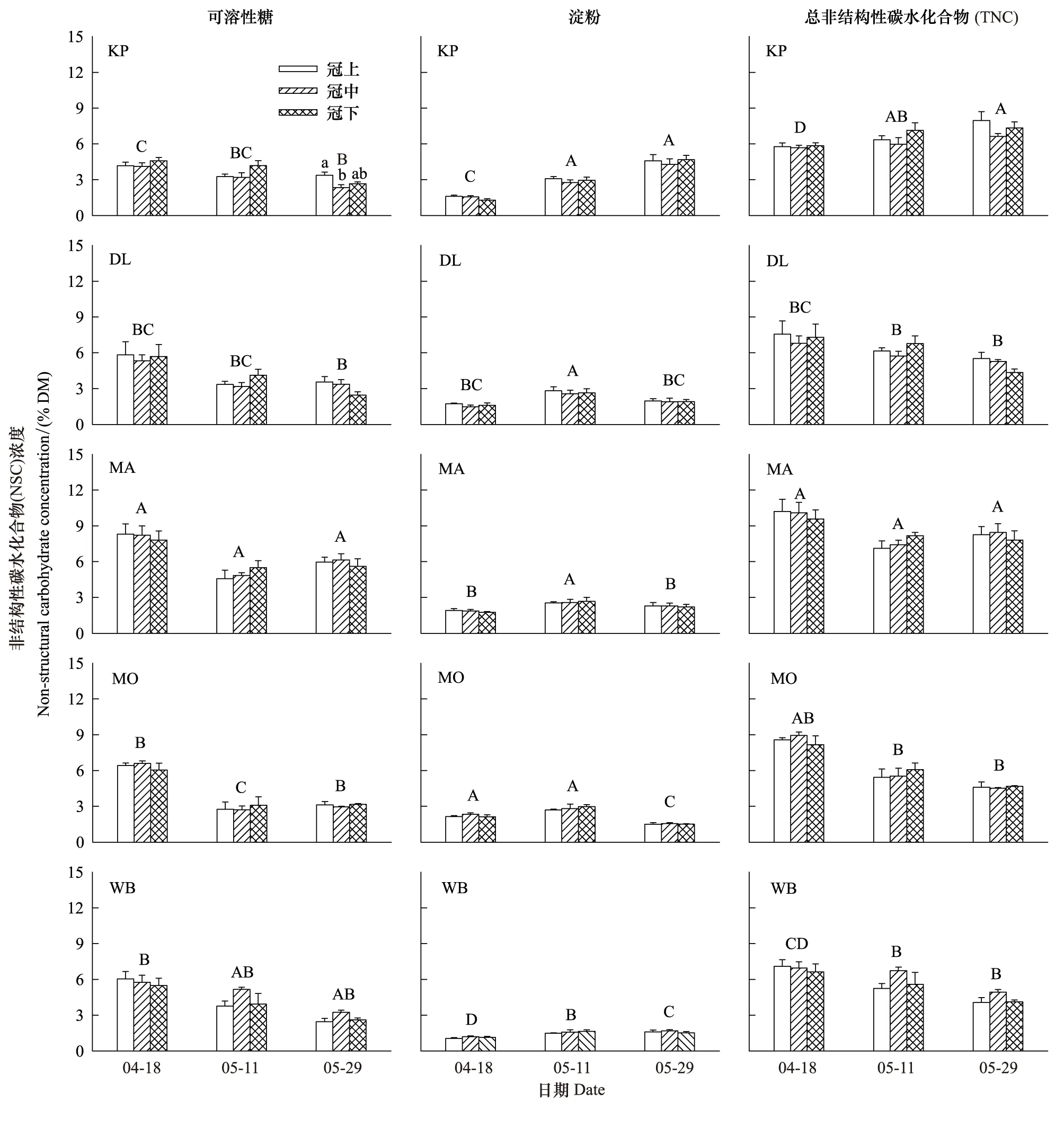
图4 生长季初期5个树种冠层细枝非结构性碳水化合物(NSC)浓度的垂直变化(平均值±标准误,n=3)Fig.4 Vertical variations in non-structural carbohydrates (NSC) concentrations in small branches for the five tree species in the early growing season(mean±se,n=3)误差线上方不同小写和大写字母分别表示冠层间和种间显著差异组别(P < 0.05);树种代码参见表1
5个树种细枝NSC浓度的高低因采样时间而异。4月水曲柳可溶性糖显著高于其他树种,蒙古栎淀粉显著高于其他树种,水曲柳TNC显著高于其他树种(蒙古栎除外);红松可溶性糖、淀粉和TNC均显著低于其他树种。5月中旬水曲柳可溶性糖显著高于其他树种(白桦除外),蒙古栎最低;白桦淀粉显著低于其他树种;水曲柳TNC显著高于其他树种(红松除外)。5月末水曲柳可溶性糖显著高于其他树种(白桦除外),红松淀粉显著高于其他树种,红松和水曲柳TNC显著高于其他树种(图4)。
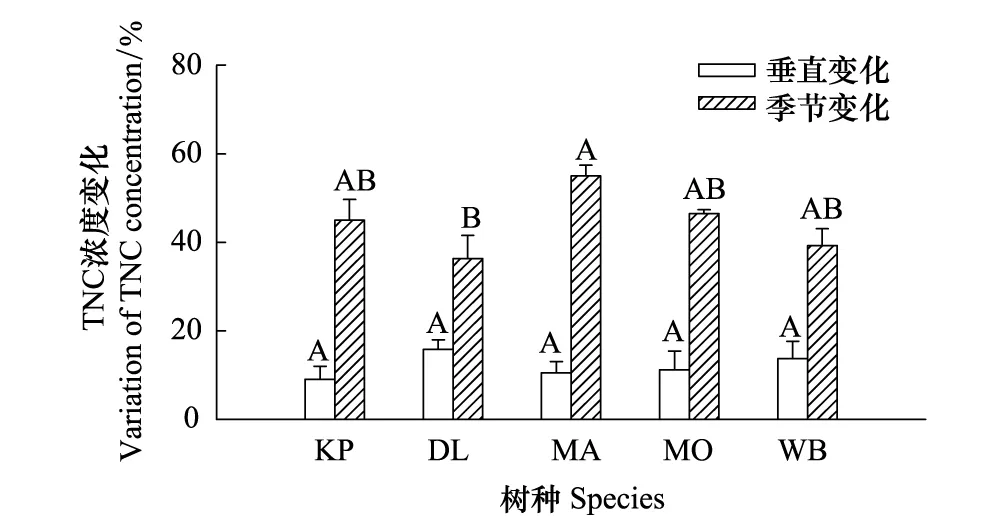
图5 5个树种细枝总非结构性碳水化合物(TNC)浓度的季节和垂直相对变化比较(平均值±标准误,n=3)Fig.5 Comparison of vertical and seasonal relative fluctuations in total non-structural carbohydrates (TNC) concentration in small branches among the five tree species(mean+se,n=3)误差线上方不同字母表示种间显著差异组别(P < 0.05);树种代码参见表1
5个树种冠层细枝TNC垂直变化波动于9.1%(红松)—15.8%(兴安落叶松)之间,平均为12.1%(图5)。相对季节变化(季节最大值与最小值之差与季节最大值的比值)波动于36.4%(兴安落叶松)—55.1%(水曲柳)之间,平均为44.5%(图5)。5个树种冠层细枝TNC的垂直变异显著大于季节动态(P< 0.05),前者平均仅占后者的27%。
2.3 粗枝NSC浓度的轴向变化
水曲柳、蒙古栎和白桦粗枝的枝皮、边材和心材的NSC浓度变化趋势大体相似,总体上随着距树枝基部距离的增加而升高(图6)。3个树种可溶性糖和TNC浓度均为:枝皮 > 边材 > 心材(白桦无法根据颜色区分心材);而淀粉浓度较低,且枝皮和边材浓度接近(图6)。3个树种相比,水曲柳粗枝组织中可溶性糖和TNC浓度略高于蒙古栎和白桦。
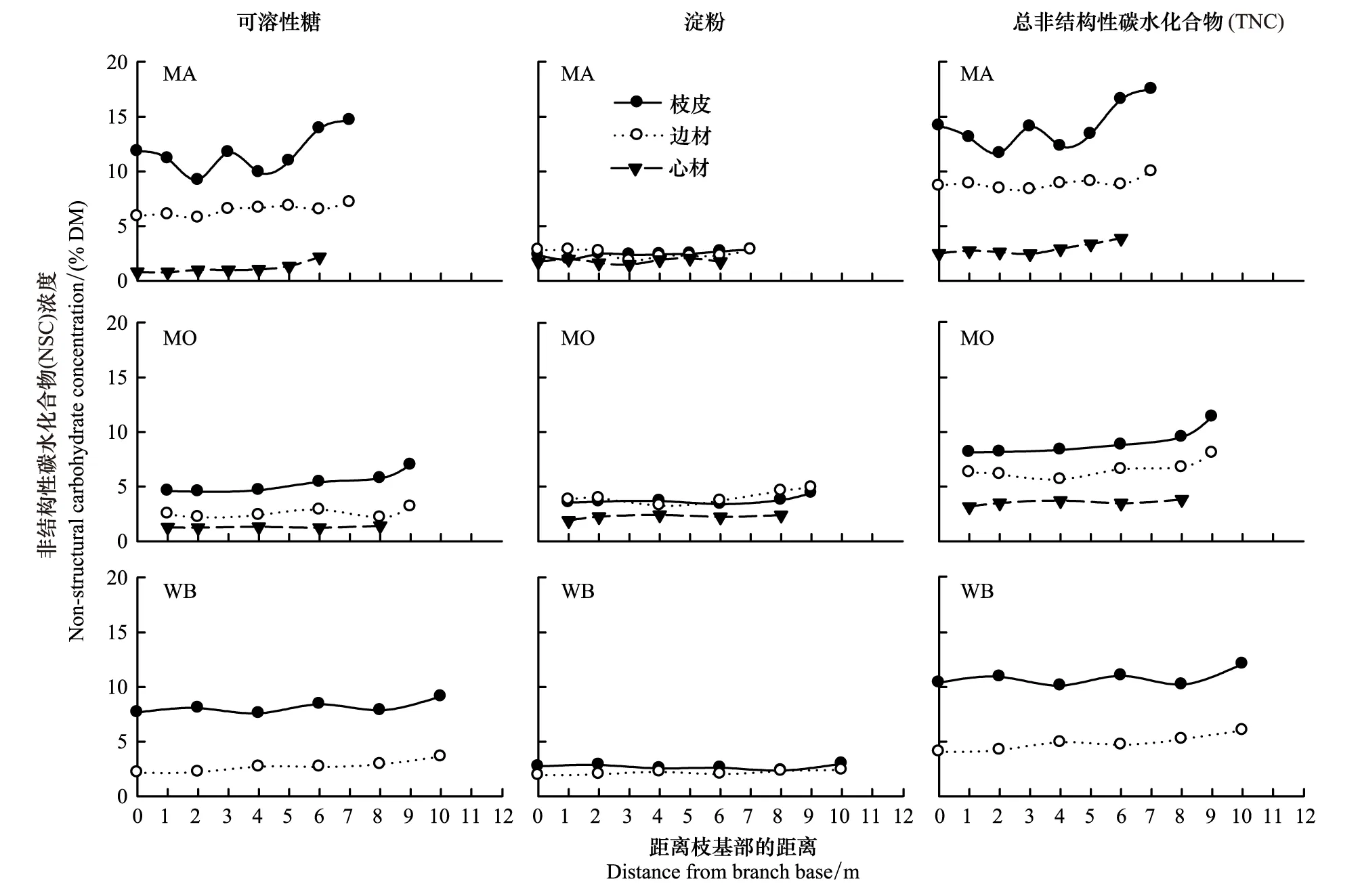
图6 3个阔叶树种粗枝的枝皮、边材和心材非结构性碳水化合物(NSC)浓度的轴向变化Fig.6 Axial variations in non-structural carbohydrate (NSC) concentrations in bark, sapwood and heartwood of coarse branches for the three broadleaved tree species树种代码参见表1
2.4 粗枝NSC浓度的径向变化
水曲柳、蒙古栎和白桦粗枝各组织的NSC浓度总体上随着枝径增加而降低(除了水曲柳边材淀粉和白桦枝皮淀粉之外,线性回归方程的斜率均为负值;表2)。具体而言,可溶性糖浓度与枝径显著负相关的组织有:水曲柳边材和心材、蒙古栎枝皮和白桦边材;淀粉仅水曲柳枝皮与枝径显著负相关;TNC随着枝径增加而显著降低的组织有:水曲柳边材和心材、蒙古栎枝皮和边材以及白桦边材。从总线性回归方程的斜率来看,环孔材树种水曲柳和蒙古栎枝皮NSC浓度随着枝径增加更明显(蒙古栎淀粉除外),而散孔材树种白桦边材降低更明显。
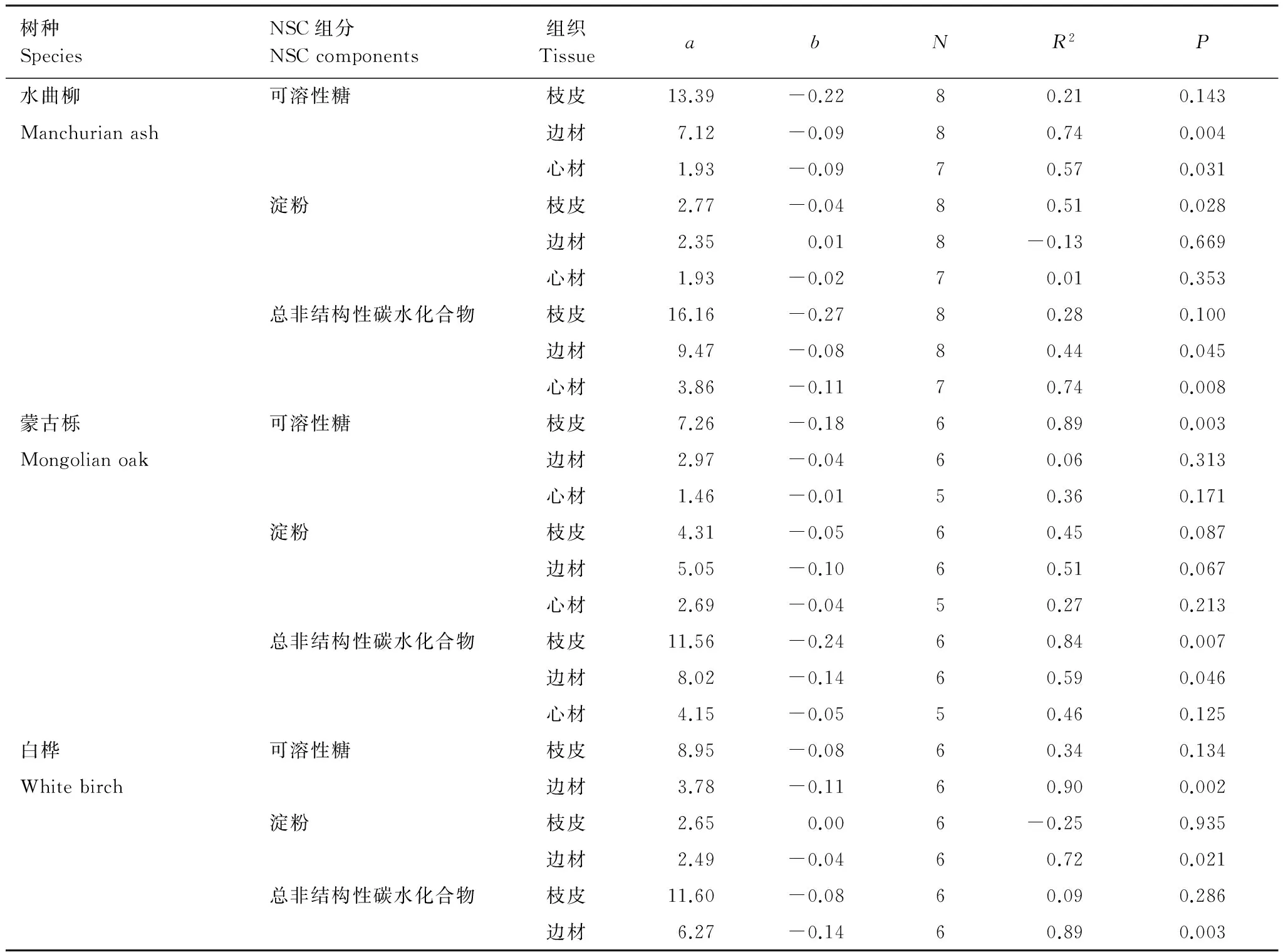
表2 3个阔叶树种粗枝NSC浓度与枝径的回归方程
3 讨论
3.1 叶NSC浓度的冠层内变异
本研究5个树种冠层高度间叶NSC浓度在大多数采样时间差异不显著(图1)。这与已往的研究结果大体一致:中国长白山10月初红松冠层上中下当年和1年生叶可溶性糖和TNC[21]、阿尔卑斯山生长季瑞士五针松冠层上中下2年生叶TNC[19]、中国藏东南的色季拉山7月底长苞冷杉和方枝柏冠层上中下不同年龄叶NSC[20]、巴拿马热带雨林17个树种不同位置的阳生叶和阴生叶NSC[24]均差异不显著。以往研究仅在部分采样时间发现有几个常绿针叶树种冠层高度间NSC浓度差异显著:红松冠上层叶淀粉和长苞冷杉冠上层当年生叶可溶性糖显著高于下层[20-21],湿地松(Pinuselliottii)春季冠上层叶可溶性糖和淀粉浓度也显著高于下层[37],挪威云杉(Piceaabies)针叶NSC浓度在9月中旬随着高度增加而增加[38]。红松[39]、兴安落叶松[40]、白桦[40]以及蒙古栎和水曲柳叶片NSC浓度并没有像叶片结构性状和光合速率一样随着冠层部位的下降而普遍递减。这些研究结果表明,通常冠层高度并没有显著影响叶NSC浓度,可能与树冠上层较强的输出能力有关。尽管其深层次的原因仍需进一步研究,但冠层高度间的差异通常并不是估算整个树冠叶NSC含量的主要误差来源。
叶龄是影响常绿树种针叶光合作用速率的重要因素,因此,NSC浓度也可能具有类似的变化规律。本研究发现,生长季中期红松针叶NSC浓度随着叶龄增加而升高,但生长季后期和休眠期叶龄间差异减小(图3)。Li等[19]研究发现,7月阿尔卑斯山瑞士五针松1年生叶NSC浓度均显著高于当年生叶,但在生长季末期这种差异消失。7月末长苞冷杉当年生到5年生叶的NSC浓度随着叶龄先增大(< 2a)后减小(> 2a)[20],挪威云杉当年到2年生叶TNC浓度逐渐增加[41]。本研究红松多年生叶为2—5a混合采样,但以2年生叶为主,因此与长苞冷杉[20]的结果一致。生长季中期当年生针叶尚未成熟,NSC浓度较低可能与其光合能力低有关。此外,本研究10月下旬红松多年生叶淀粉浓度显著高于当年生叶(图3),与Yan等[21]同期对红松1年生叶的研究结果一致。老龄叶的NSC浓度较高,可能与碳运输过程中酶活性降低或与韧皮部从老叶输出光合产物的能力降低有关[41]。Li等[20]认为叶开始衰老时光合能力逐渐降低,其NSC浓度也开始下降。综上所述,常绿针叶树的叶龄是冠层NSC库估算应该考虑的因素。
本研究发现5月末红松针叶TNC以淀粉为主,而落叶树种的当年生叶则以可溶性糖为主,这与王兆国和王传宽[25]春季红松和白桦的结果一致,但与樟子松(Pinussylvestrisvar.mongolica)春季可溶性糖和淀粉浓度接近不同。常绿树种红松针叶TNC明显高于落叶阔叶树种的当年生叶(图1),可能是因为5月末红松新枝叶尚未开始迅速生长,老龄叶以淀粉形式积累了大量NSC的缘故[22,42]。红松针叶NSC浓度与以往同一物候期的研究结果很接近[17,21,25,31];而兴安落叶松和蒙古栎叶TNC浓度略低于于丽敏等[17]的结果,可能是由于本研究中5月末叶片尚未完全成熟的缘故[23]。本研究白桦叶TNC浓度15%—17% DM与印婧婧等[29]在夏季内蒙古白桦的17% DM一致,同时也处于王兆国和王传宽[25]的14%—22% DM范围之内。
3.2 不同冠层细枝NSC浓度
本研究5个树种冠层高度间细枝的可溶性糖、淀粉和TNC浓度大部分采样时间差异不显著(图4)。这与欧洲山毛榉和无梗花栎[18]的结果吻合,但与北美黄杉细枝(40—70 cm长,包括枝皮和木质部)NSC浓度[16]和北美黄松树枝边材TNC浓度[28]随着高度增加而增加的结果不一致,这可能与其特别高大的冠层有关(两树种高度分别为2—57 m和6—38 m)。高大的树木树冠上层光合作用与生长速率极有可能受水分限制[16,28]。然而湿地松冠层最高处2 m的NSC浓度高于最低处2 m[37],可能是由于该树种为强喜光树种的缘故。本研究5个树种冠层细枝NSC浓度垂直差异不显著,可能是因为水分并没有明显限制韧皮部从树冠上层向树干和根系的NSC运输[16],亦或是树冠上层树枝较强的碳汇(生长和呼吸旺盛)抵消了较强的碳源功能。
由于细枝TNC浓度存在很大的季节动态[26],而且TNC随着枝径产生很大的变化(图6),同时树枝去皮与否也会对TNC产生较大的影响[18],因此不同研究之间的比较存在很大的困难。总体而言,5月末红松、兴安落叶松和蒙古栎细枝TNC浓度与于丽敏等[17]的结果很接近。本研究发现,5个树种冠层细枝TNC的垂直变异平均仅占其季节波动的27%。由于连续采样对树木产生较大的破坏,因此研究季节动态时采用冠层中上部的树枝代表整个冠层是行之有效的策略。
3.3 粗枝NSC浓度的变化
粗枝是树冠生物量的主体,因此量化粗枝NSC浓度的空间变异对估算整个树冠的NSC含量非常重要。本研究发现,多数树种粗枝组织的NSC浓度从树枝基部到梢头总体上呈增加趋势(图6),即NSC浓度随着枝径增加而逐渐降低(表2)。这与无梗花栎和欧洲山毛榉树枝(枝基部除外)NSC浓度随着枝径增加而降低[18]的结果一致。Li等[19]研究发现,瑞士五针松树枝边材NSC浓度从外围向中心逐渐降低,认为相同质量的幼龄组织比老龄组织存储的TNC更多。这可能是因为幼龄组织通常生长在枝条的顶端,可优先获取叶片的光合产物。因此,随着组织活力由树冠外侧向中心(或由梢头到枝基)逐渐降低,粗枝皮和边材的NSC逐渐降低。此外,带皮或去皮测量树枝TNC,不仅影响树枝TNC大小,还会影响可溶性糖与淀粉的分配比例,但不影响树枝TNC随着枝径的变化规律[18]。枝皮中的NSC主要存储在韧皮部,环孔材树种水曲柳和蒙古栎枝皮厚度随着枝径增加而增厚主要是木栓层和死皮增厚的结果。因此,环孔材树种粗枝皮NSC随着枝径增加而降低,并且比白桦更明显。
很少有研究测定树枝心材的NSC。令人惊奇的是,本研究发现水曲柳和蒙古栎(环孔材树种)的粗枝心材NSC也表现出与枝皮和边材相似的变化规律。另外,本研究NSC浓度随着枝径和枝龄的增加呈逐渐降低的趋势,表明树枝心材中的NSC并非完全失去生理活性。尽管以往研究认为树木心材不存在NSC或即便存储NSC也是不可利用的[43-44],但一些环孔材树种和针叶树种的树干心材(或树干深处)的确存在NSC[22,33,45],而成年散孔材树种白桦和紫椴(Tiliaamurensis)树干最内部仍然存在大量NSC[32]。由于很多成年树木(尤其是环孔材树种)的生物量以心材为主,将树木心材认为是一个死亡的组织可能低估了整株树木的存储功能[33]。心材中的老龄NSC随着组织年龄增加而降低表明心材NSC可以移动,但心材NSC在何种程度上可以利用以及从心材(通常认为没有活细胞)向边材的运输机理尚不清楚,是树木生理生态学中值得进一步研究的课题。
4 结论
5个温带树种冠层叶和细枝NSC浓度的空间变异特征表明,光环境和光合作用的垂直变化梯度并没有引起生物量组织的NSC浓度产生类似梯度变化。5个树种冠层细枝TNC垂直变异与季节动态相比很小,因此采用冠层中上部的树枝代表整个冠层是研究树枝NSC季节动态的实用而有效的采样策略。粗枝NSC浓度总体上随着枝径增加而降低,在估算整个冠层NSC含量时应予考虑。
致谢:感谢帽儿山森林生态站提供野外基础支持。
[1] 潘庆民, 韩兴国, 白永飞, 杨景成. 植物非结构性贮藏碳水化合物的生理生态学研究进展. 植物学通报, 2002, 19(1): 30-38.
[2] Chapin F S, Schulze E, Mooney H A. The ecology and economics of storage in plants. Annual Review of Ecology and Systematics, 1990, 21(1): 423-447.
[3] Schädel C, Blöchl A, Richter A, Hoch G. Short-term dynamics of nonstructural carbohydrates and hemicelluloses in young branches of temperate forest trees during bud break. Tree Physiology, 2009, 29(7): 901-911.
[4] Ögren E. Fall frost resistance in willows used for biomass production. II. predictive relationships with sugar concentration and dry matter content. Tree Physiology, 1999, 19(11): 755-760.
[5] Sala A, Woodruff D R, Meinzer F C. Carbon dynamics in trees: feast or famine?. Tree Physiology, 2012, 32(6): 764-775.
[6] 董蕾, 李吉跃. 植物干旱胁迫下水分代谢、碳饥饿与死亡机理. 生态学报, 2013, 33(18): 5477-5483.
[7] Bond W J, Midgley J J. Ecology of sprouting in woody plants: the persistence niche. Trends in Ecology and Evolution, 2001, 16: 45-51.
[8] Poorter L, Kitajima K. Carbohydrate storage and light requirements of tropical moist and dry forest tree species. Ecology, 2007, 88(4): 1000-1011.
[9] Myers J A, Kitajima K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest. Journal of Ecology, 2007, 95(2): 383-395.
[10] McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739.
[11] Körner C. Carbon limitation in trees. Journal of Ecology, 2003, 91(1): 4-17.
[12] Martínez-Vilalta J. Carbon storage in trees: pathogens have their say. Tree Physiology, 2014, 34(3): 215-217.
[13] Palacio S, Hoch G, Sala A, Körner C, Millard P. Does carbon storage limit tree growth?. New Phytologist, 2014, 201(4): 1096-1100.
[14] Wiley E, Helliker B. A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth. New Phytologist, 2012, 195(2): 285-289.
[15] Dietze M C, Sala A, Carbone M S, Czimczik C I, Mantooth J A, Richardson A D, Vargas R. Nonstructural carbon in woody plants. Annual Review of Plant Biology, 2014, 65: 667-687.
[16] Woodruff D R, Meinzer F C. Water stress, shoot growth and storage of non-structural carbohydrates along a tree height gradient in a tall conifer. Plant, Cell and Environment, 2011, 34(11): 1920-1930.
[17] 于丽敏, 王传宽, 王兴昌. 三种温带树种非结构性碳水化合物的分配. 植物生态学报, 2011, 35(12): 1245-1255.
[18] Barbaroux C, Bréda N, Dufrêne E. Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species (QuercuspetraeaandFagussylvatica). New Phytologist, 2003, 157(3): 605-615.
[19] Li M H, Hoch G, Körner C. Spatial variability of mobile carbohydrates withinPinuscembratrees at the alpine treeline. Phyton, 2001, 41(2): 203-213.
[20] Li M C, Kong G Q, Zhu J J. Vertical and leaf-age-related variations of nonstructural carbohydrates in two alpine timberline species, southeastern Tibetan Plateau. Journal of Forest Research, 2009, 14(4): 229-235.
[21] Yan C F, Han S J, Zhou Y M, Wang C G, Dai G H, Xiao W F, Li M H. Needle-age related variability in nitrogen, mobile carbohydrates, and δ13C withinPinuskoraiensistree crowns. PloS One, 2012, 7(4): e35076.
[22] Hoch G, Richter A, Körner C. Non-structural carbon compounds in temperate forest trees. Plant, Cell and Environment, 2003, 26(7): 1067-1081.
[23] Landhäusser S M, Lieffers V J. Seasonal changes in carbohydrate reserves in mature northernPopulustremuloidesclones. Trees-Structure and Function, 2003, 17(6): 471-476.
[24] Würth M K R, Peláez-Riedl S, Wright S J, Körner C. Non-structural carbohydrate pools in a tropical forest. Oecologia, 2005, 143(1): 11-24.
[25] 王兆国, 王传宽. 三种温带树种叶片呼吸的时间动态及其影响因子. 生态学报, 2013, 33(5): 1456-1464.
[26] 张海燕, 王传宽, 王兴昌. 温带12个树种新老树枝非结构性碳水化合物浓度比较. 生态学报, 2013, 33(18): 5675-5685.
[27] Niinemets Ü. Photosynthesis and resource distribution through plant canopies. Plant, Cell and Environment, 2007, 30(9): 1052-1071.
[28] Sala A, Hoch G. Height-related growth declines in ponderosa pine are not due to carbon limitation. Plant, Cell and Environment, 2009, 32(1): 22-30.
[29] 印婧婧, 郭大立, 何思源, 张蕾. 内蒙古半干旱区树木非结构性碳、氮、磷的分配格局. 北京大学学报: 自然科学版, 2009, 45(3): 519-527.
[30] 周永斌, 吴栋栋, 于大炮, 隋琛莹. 长白山不同海拔岳桦非结构碳水化合物含量的变化. 植物生态学报, 2009, 33(1): 118-124.
[31] Yan C F, Han S J, Zhou Y M, Zheng X B, Yu D D, Zheng J Q, Dai G H, Li M H. Needleδ13C and mobile carbohydrates inPinuskoraiensisin relation to decreased temperature and increased moisture along an elevational gradient in NE China. Trees, 2013, 27(2): 389-399.
[32] 张海燕, 王传宽, 王兴昌, 成方妍. 白桦和紫椴树干非结构性碳水化合物的空间变异. 应用生态学报, 2013, 24(11): 3050-3056.
[33] Zhang H Y, Wang C K, Wang X C. Spatial variations in non-structural carbohydrates in stems of twelve temperate tree species. Trees, 2014, 28(1): 77-89.
[34] Zhang Q Z, Wang C K. Carbon density and distribution of six Chinese temperate forests. Science China Life Sciences, 2010, 53(7): 831-840.
[35] 祝宁, 江洪, 金永岩. 中国东北天然次生林主要树种的物候研究. 植物生态学与地植物学学报, 1990, 14(4): 336-349.
[36] Buysse J, Merckx R. An improved colorimetric method to quantify sugar content of plant tissue. Journal of Experimental Botany, 1993, 44(10): 1627-1629.
[37] Gholz H L, Cropper W P. Carbohydrate dynamics in maturePinuselliottiivar.elliottiitrees. Canadian Journal of Forest Research, 1991, 21(12): 1742-1747.
[38] Niinemets Ü. Distribution patterns of foliar carbon and nitrogen as affected by tree dimensions and relative light conditions in the canopy ofPiceaabies. Trees, 1997, 11(3): 144-154.
[39] 霍宏, 王传宽. 冠层部位和叶龄对红松光合蒸腾特性的影响. 应用生态学报, 2007, 18(6): 1181-1186.
[40] 王秀伟, 毛子军. 兴安落叶松人工林冠层气体交换的时空特性. 林业科学, 2007, 43(11): 43-49.
[41] Niinemets Ü. Acclimation to low irradiance inPiceaabies: influences of past and present light climate on foliage structure and function. Tree Physiology, 1997, 17(11): 723-732.
[42] Fischer C, Höll W. Food reserves of scots pine (PinussylvestrisL.) I. seasonal changes in the carbohydrate and fat reserves of pine needles. Trees, 1992, 6(3): 147-155.
[43] Taylor A M, Gartner B L, Morrell J J. Heartwood formation and natural durability-a review. Wood and Fiber Science, 2002, 34(4): 587-611.
[44] Hoch G. Cell wall hemicelluloses as mobile carbon stores in non-reproductive plant tissues. Functional Ecology, 2007, 21(5): 823-834.
[45] Newell E A, Mulkey S S, Wright J S. Seasonal patterns of carbohydrate storage in four tropical tree species. Oecologia, 2002, 131(3): 333-342.
Within-crown variation in concentrations of non-structural carbohydrates of five temperate tree species
ZHANG Haiyan, WANG Chuankuan*, WANG Xingchang
CenterforEcologicalResearch,NortheastForestryUniversity,Harbin150040,China
Non-structural carbohydrates (NSC) are important carbon reserves, particularly in trees, because they represent a large pool of biomass. Diverse strategies for carbon use and allocation are closely connected to ecological and evolutionary theories of trade-offs and niche partitioning. Crown is not only the source of NSC in a tree, but also a key NSC storage pool. However, few studies investigated within-crown variation in NSC concentrations, although such variation may introduce uncertainties in estimating NSC allocation in trees. In this study, we measured NSC concentrations in foliage and branches of five temperate tree species in northeastern China. The species included one evergreen conifer (Pinuskoraiensis), one deciduous conifer (Larixgmelinii), and three deciduous broadleaved species (Quercusmongolica,Fraxinusmandshurica, andBetulaplatyphylla). The objectives were to (1) examine vertical differences in NSC concentrations among foliage and small branches of different ages (diameter ≤ 3 cm) within the crown, (2) examine axial variations in NSC in bark, sapwood and heartwood of coarse branches (i.e. first order branches), and (3) explore the relationship between NSC concentration and diameter in coarse branches. Branch samples were randomly taken from the upper-, mid-and lower-crown layers 3—7 times across the season. The samples were divided into foliage (current-year, one-year-old, and older than one year) and small branches. In late October, one coarse branch from each of the three broadleaved species was sampled and separated into 1—2 m-long sections consisting of bark, sapwood, and heartwood. All samples were analyzed for NSC concentration (including soluble sugars and starch) with a modified phenol-sulfuric method. The results showed that: (1) There were no significant vertical variations in foliage NSC concentration for any species exceptL.gmelinii, for which the starch concentration in late May was significant higher in the mid-crown and soluble sugar concentration in mid-August was higher in the upper-crown. Needle-age significantly affected needle NSC concentration ofP.koraiensisin the mid-growing season, but this effect was insignificant in the late growing season and dormant season. (2) There were no significant vertical variations in NSC concentration in small branches for any species exceptP.koraiensis, for which the concentration of soluble sugars in the upper-crown was significant higher than in the mid-crown in late May. (3) For the three deciduous species, the NSC concentrations in the bark, sapwood and heartwood of the coarse branches tended to increase with increasing distance from the branch base to the top, and they decreased with increasing branch diameter for all species except for starch in sapwood ofF.mandshuricaand starch in bark ofB.platyphylla. This suggests a declining trend in branch NSC concentration with source distance or tissue age. The trend of NSC concentration in branch heartwood indicates that C reserves in old heartwood are still accessible or even physiologically available. Overall, NSC concentrations in foliage and small branches varied insignificantly within the crowns of these tree species, but the effects of needle-age and branch diameter on NSC concentrations varied among species, tissue, or season; these within-crown variations should be taken into account when making NSC pool estimates for the whole tree.
non-structural carbohydrates (NSC); foliage; branch; spatial variation; temperate forest
国家“十二五”科技支撑项目(2011BAD37B01); 教育部长江学者和创新团队发展计划(IRT1054)
2014-02-08; < class="emphasis_bold">网络出版日期:
日期:2014-12-04
10.5846/stxb201402080223
*通讯作者Corresponding author.E-mail: wangck-cf@nefu.edu.cn
张海燕, 王传宽, 王兴昌.5个温带树种冠层枝叶非结构性碳水化合物浓度的空间变异.生态学报,2015,35(19):6496-6506.
Zhang H Y, Wang C K, Wang X C.Within-crown variation in concentrations of non-structural carbohydrates of five temperate tree species.Acta Ecologica Sinica,2015,35(19):6496-6506.
猜你喜欢
《学习方法报》小学语文四年级(2023年26期)2023-04-29 00:44:03
格言·校园版(2022年17期)2022-07-06 22:01:48
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
林业科技情报(2021年1期)2021-04-14 09:10:02
文苑(2019年20期)2019-11-20 02:12:33
文苑·经典美文(2019年10期)2019-10-16 02:47:16
农民致富之友(2019年11期)2019-05-23 01:14:52
森林工程(2018年4期)2018-08-04 03:23:10
农民致富之友(2017年23期)2018-01-02 02:02:06
现代营销(创富信息版)(2016年6期)2016-11-21 06:00:03
