
芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应
2015-01-19 07:06:37张文涛王明昌张凌楠董满宇
生态学报 2015年19期
张文涛, 江 源, 王明昌, 张凌楠, 董满宇
1 北京师范大学资源学院, 北京 100875 2 北京师范大学地表过程与资源生态国家重点实验室, 北京 100875 3 北京师范大学生命科学学院, 北京 100875 4 北京市环境保护科学研究院, 北京 100037
芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应
张文涛1,3,4, 江 源1,2,*, 王明昌1, 张凌楠1, 董满宇1
1 北京师范大学资源学院, 北京 100875 2 北京师范大学地表过程与资源生态国家重点实验室, 北京 100875 3 北京师范大学生命科学学院, 北京 100875 4 北京市环境保护科学研究院, 北京 100037
为研究树木生长对气候变化的响应状况,选取芦芽山阳坡的3个海拔高度建立了华北落叶松(Larixprincipis-rupprechtii)的树轮宽度年表。年表的统计参数表明,3条年表均为研究气候信息的可靠资料。结果表明,芦芽山阳坡华北落叶松的径向生长和生长与气候的关系均具有海拔差异,中海拔(2440 m)和高海拔(2540 m)的华北落叶松具有相似年际生长变化,而二者均与低海拔(2330 m)华北落叶松的年际生长不同。低海拔华北落叶松的生长与4月平均气温和上一年11月降水量显著负相关,而中海拔和高海拔的生长均与上一年10月平均气温和6月降水量显著负相关。通过年表与气候因子之间的滑动相关分析发现,3个海拔高度华北落叶松生长与气候因子的关系均不稳定,生长与气温条件之间的显著相关关系是随着气温升高而出现的。气温的升高引起了华北落叶松生长与气温因子关系的海拔差异,以及径向生长的海拔差异。这一结果对于气候变化对植被垂直梯度影响的研究具有一定参考价值。
气候变化; 树木年轮; 海拔梯度; 芦芽山; 华北落叶松
树木年轮是研究气候变化对树木生长影响的重要工具[1]。在过去的研究中,不同生境条件下树木生长与气候因子的密切关系已经被大量证实[2-7]。然而,气候条件的改变将会影响树木生长与气候因子的关系,并进一步影响树木的径向生长过程。20世纪80年代以来,北半球高海拔高纬度地区树木生长对气温升高表现出的敏感度降低现象[8]就是这一过程的例证。此外,在阿拉斯加地区,D′Arrigo等[9]报道了白云杉(Piceaglauca)生长与气温的关系在1970年前后出现差别,随着气温升高,生长与气温之间的关系逐渐减弱。Daniels和Veblen[10]研究发现安第斯山脉林线位置南水青冈(Nothofaguspumilio)的生长在不同时段存在不同的限制因子,在湿冷的1957—1976年生长受到夏季气温的限制,在暖干的1977—1996年则受到夏季降水的限制。在中国,长白山林线红松(Pinuskoraiensis)生长对春季水分和最低气温的响应逐渐加强[11]。这些都证明了气候变化能够引起树木生长对气候响应的改变。
芦芽山位于中国华北地区,该区域自20世纪70年代开始经历了显著的增温过程[12]。增温过程将对这一地区的森林动态和树木生长产生影响。张文涛等[13]研究发现芦芽山阳坡的白杄(Piceameyeri)生长会受到气候变暖的影响,随着气温的升高,气候因子对低海拔白杄生长的限制增强,而对高海拔白杄生长的限制减弱。在过去的研究中,芦芽山阴坡的华北落叶松径向生长与气候因子的关系表现出了海拔差异[14]。这一结果在阳坡是否依然存在?并且,在对气温变化更为敏感的阳坡,气温的升高对于华北落叶松的生长具有怎样的影响?本文选取了芦芽山阳坡不同海拔高度的华北落叶松树轮样本作为对象,对生长的海拔差异以及气候变化对生长的影响进行了研究,为华北地区气候变化背景下的森林保护提供理论依据。
1 研究区域
芦芽山(北纬38 °35 ′—38 °45 ′,东经111 °50 ′—112 °5 ′)位于山西省西北部,吕梁山脉北端,是汾河的源头,其主峰荷叶坪海拔2783 m。芦芽山属于暖温带半湿润气候区,具有明显的大陆性气候特点。根据距离最近的五寨气象站(北纬38 °55 ′,东经111 °49 ′)的资料,其年平均气温为4.9℃,7月平均气温最高(22.4℃),1月平均气温最低(-10.2℃),年降水量472.7 mm,6—9月降水量约占全年降水量的76.0%。芦芽山具有明显的植被垂直更替,根据马子清[15]的研究,随海拔升高,植被依次为:森林草原带(1300—1500 m)、落叶阔叶林带(1350—1700 m)、针阔叶混交林带(1700—1850 m)、寒温性针叶林带(1750—2600 m)和亚高山灌丛草甸带(2450—2772 m)。土壤类型随海拔升高依次为山地褐土、山地淋溶褐土、棕色森林土和亚高山草甸土[16]。其中,华北落叶松主要分布于寒温性针叶林带内。
2 材料和方法
2.1 样本采集
2009年8月进行了样本的采集工作。在主峰荷叶坪附近的阳坡选取3个海拔高度设立采样点,低海拔采样点L位于2330 m,中海拔采样点M为2440 m,高海拔采样点H为2540 m(表1)。实际工作中,阳坡华北落叶松分布下限约在2050 m,但通过树轮测量,该海拔的华北落叶松树龄过短,平均树龄仅在20a左右,幼龄效应明显,因此该采样点放弃,而使用2330 m的采样点作为气候影响显著的最低海拔采样点。此外,高海拔采样点H位于华北落叶松在阳坡的分布上限,为天然林线。
采样点均选在中坡位,坡度控制在20 °左右,郁闭度在0.6左右。所有采样点林下土壤为棕色森林土,排水性能良好。在每个采样点内,选择生长良好的华北落叶松约15株,钻取胸径位置的树轮样本,每株树沿垂直与平行坡向的方向钻取树芯2个。同时,在每个采样点随机设立20 m×20 m的乔木样方3个,记录样方内树木的胸径(DBH)、树高和树种。
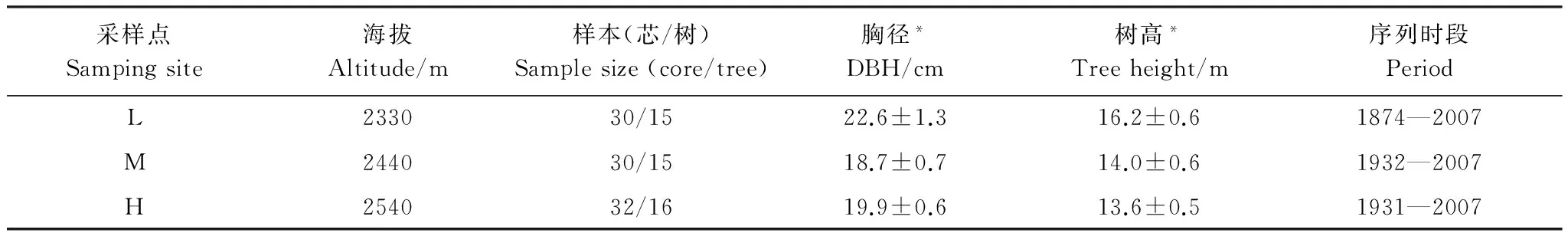
表1 采样点概况
2.2 建立年表
树轮样本带回实验室后,依次进行样本的晾干、固定和打磨。在显微镜下进行定年[17]。然后使用测量精度为0.01 mm的LINTAB树轮宽度测量仪(TM5, Rinntech, Heidelberg, Germany)进行轮宽的测量。此后,运行COFECHA程序[18]对原始轮宽序列和交叉定年结果进行质量检验,对出现问题的序列进行检查。最终使用ARSTAN程序完成年表的制作,其中的去趋势方法选取负指数函数。由于RES年表(Residual chronology)具有高质量的高频信息[19],因此被选择作为后续分析的对象。
2.3 气候资料
参考Kimball和Keifer[20],分析过程中选取了采样点附近五寨(海拔1401.1 m)、原平(北纬38 °44 ′,东经112 °43 ′,海拔828.2 m)和河曲(北纬39 °23 ′,东经110 °09 ′,海拔862 m)3处气象站气象资料的平均值作为气候数据来源。数据长度为1957—2007年,所选指标包含月平均气温,月平均最低气温,月平均最高气温和月降水量。
通过对研究区1957—2007年的年平均气温和年降水量的突变检验和时间变化趋势分析发现(图1),研究区气温在1996年前后存在突变点,1996—2007年的年平均气温明显高于1957—1995年。与之对应的,降水量虽然减少,但趋势并不明显。气温的显著升高,为本次树木生长对气候变化响应的研究提供了可能。
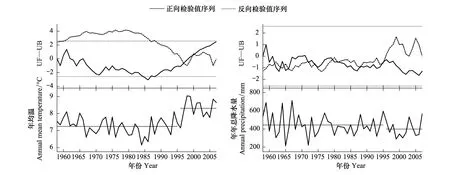
图1 研究区年均温和年总降水量的变化趋势和突变检验Fig.1 The variation and the abrupt change of annual mean temperature and annual precipitation突变检验方法使用Mann-Kendall方法,UF:正向检验值序列,UB:反向检验值序列,点状直线表示95%的显著水平
2.4 分析方法
通过计算年表的统计参数来评估年表的可靠性。其中信噪比(SNR)和样本总体解释量(EPS)表明了样本的代表性,二者值越大表明年表中用于分析的环境信息越多。样本总体解释量存在0.85的阈值,若大于该值则表明年表达到分析气候信息的条件[21]。
计算不同海拔高度的3个年表之间的滑动相关系数,用以评估华北落叶松生长节律的海拔差异及其时间动态。同样计算3个年表与月份气候因子之间的Pearson相关系数和滑动相关系数,从而获得不同海拔高度树木生长与气候因子之间的关系,以及关系的稳定性。气候因子来自上一年9月至当年11月。Pearson相关系数使用SPSS软件计算实现,滑动相关系数通过DendroClim2002[22]计算完成。
3 结果与分析
3.1 年表的统计特征
表2给出了3条华北落叶松年表的统计参数,其中平均敏感度介于0.16—0.21,标准差介于0.14—0.19,序列间的相关系数为0.39—0.59,第一主成分的解释量达到47.9%—61.1%,表明3个采样点的树木生长对气候变化均具有一定的敏感性。其中,中海拔采样点M的年表各项统计参数均最小,生长对气候的敏感性最低,这是因为中海拔位置气候条件较为适宜,可能存在其它地形或者微环境因子对生长的影响。然而,3个年表的信噪比达到6.5—28.6,EPS达到0.87—0.97,均超过了0.85的最低阈值[21],证明3个年表都是研究气候响应的可信资料。
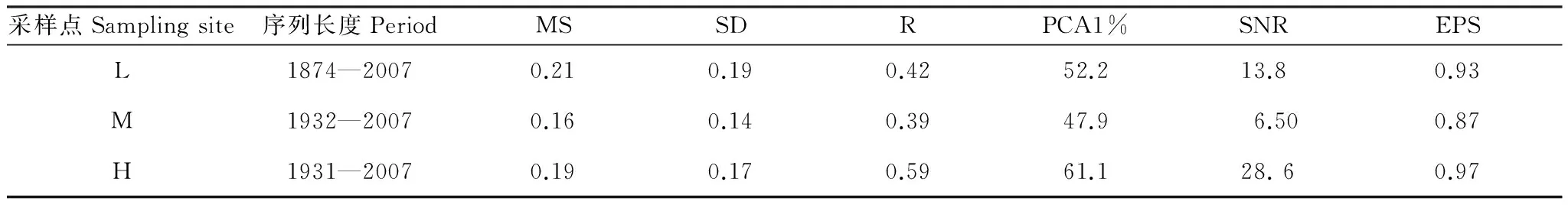
表2 年表的统计参数
3.2 不同海拔华北落叶松的径向生长节律
不同年表之间的滑动相关系数体现了不同海拔华北落叶松径向生长年际变化的差异(图2)。图中表明,随着气温的变化,不同海拔华北落叶松年表之间的相关关系并不稳定。其中,中海拔采样点M和高海拔采样点H年表之间的相关系数保持增大,最终甚至达到0.9左右。而低海拔采样点L与高海拔采样点H之间的相关系数呈降低趋势,它与中海拔采样点M之间的相关系数虽然有所增加,但始终在一个较低的水平。以上结果表明,芦芽山阳坡中海拔和高海拔的华北落叶松具有相似的年际生长变化,而低海拔的华北落叶松径向生长则与二者存在差别,而且这一差别在一定程度上与气温的改变同步。
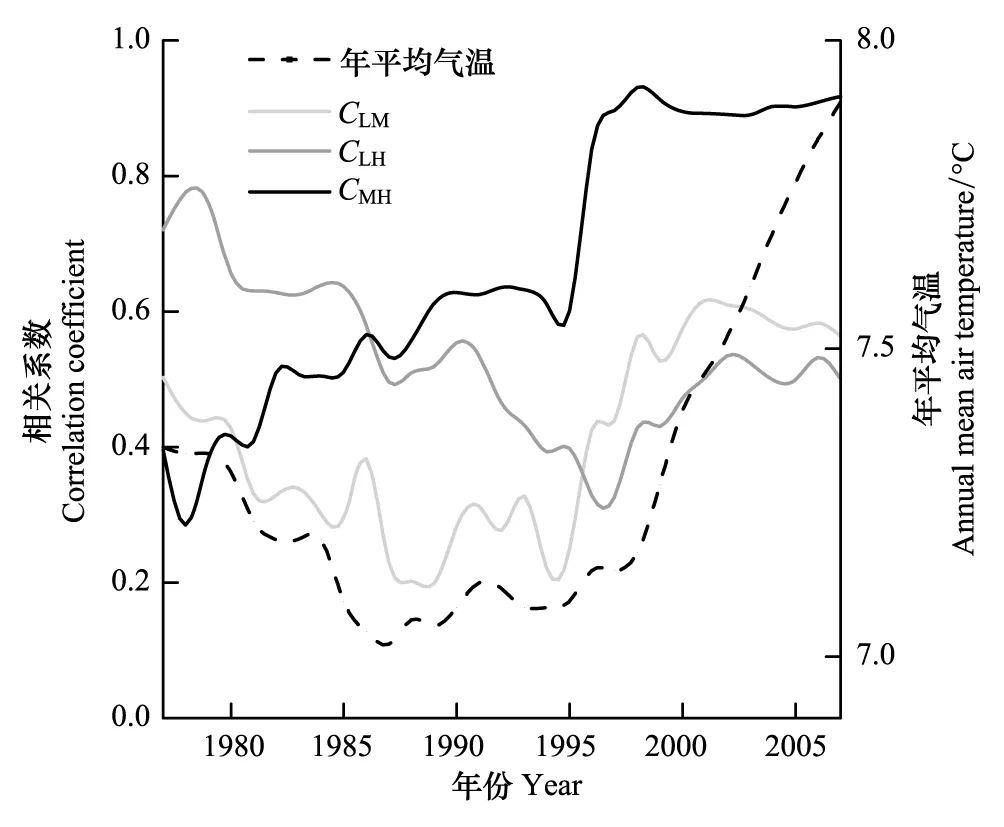
图2 不同采样点年表之间的滑动相关系数 Fig.2 Moving correlation coefficients calculated between each pair of chronologies at different sites滑动窗口为20a; CLM,CLH和CMH分别为采样点L和M,L和H,以及M和H之间的相关系数
3.3 树木生长与气候因子的关系
图3表明了不同海拔高度采样点的华北落叶松年表与月份气候因子之间的相关关系。其中,中海拔采样点M和高海拔采样点H华北落叶松生长与气候因子的关系相似,而与低海拔采样点L则存在差异,这对应了3个采样点华北落叶松径向生长年际变化的异同。具体而言,低海拔采样点L的华北落叶松生长与上一年10月和当年4月的气温存在负相关,与上一年11月的降水量也存在显著负相关关系。中海拔采样点M和高海拔采样点H的华北落叶松生长与上一年10月和11月的气温存在负相关关系,与当年6月的降水量存在负相关关系。综合以上,不同海拔高度华北落叶松与气候因子的关系存在差异。
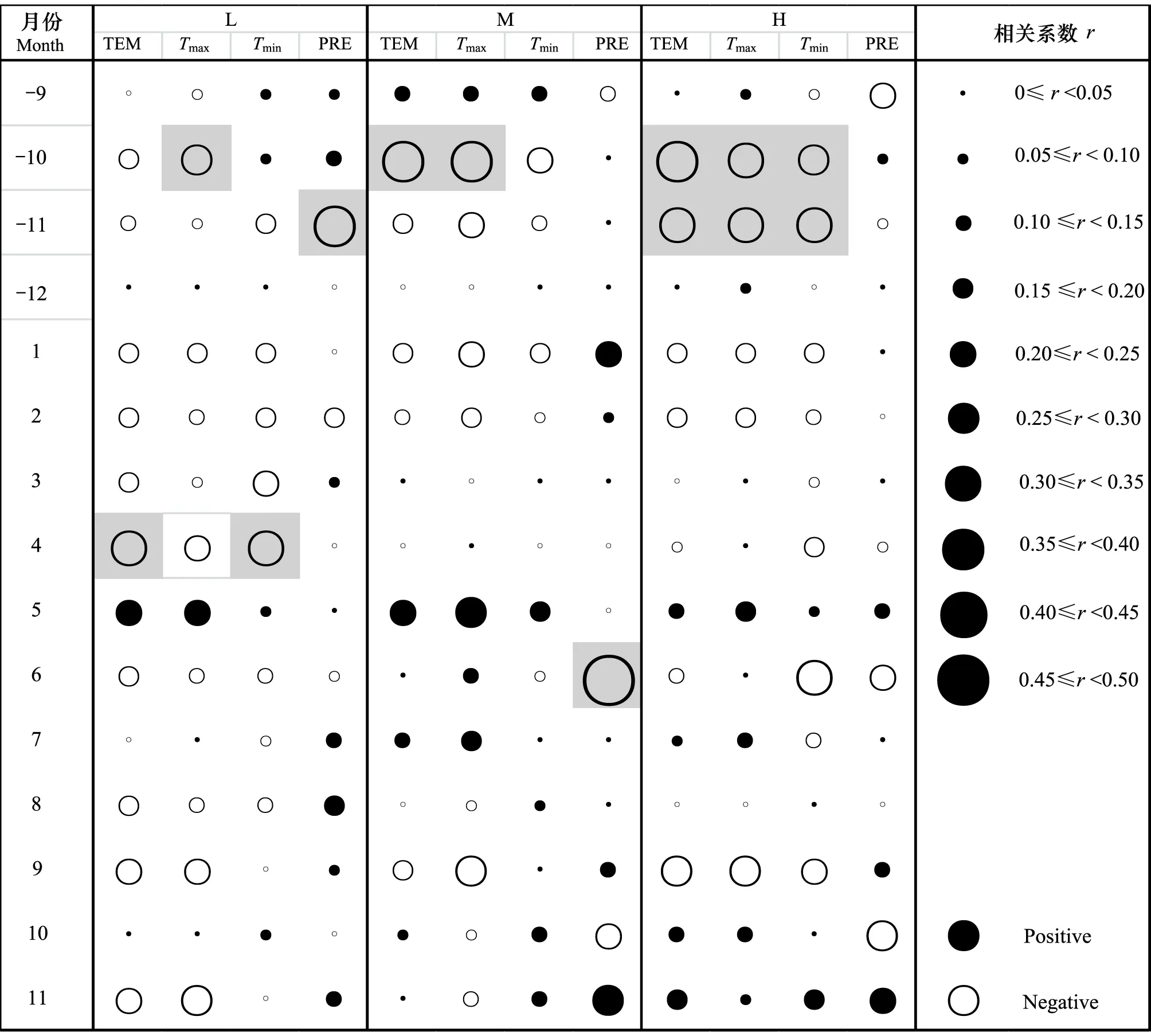
图3 不同采样点华北落叶松径向生长与月份气候因子之间的相关关系Fig.3 Correlation coefficient between monthly climatic factors and chronologies at different sampling sitesTEM: 月平均气温,Tmax: 月平均最高气温,Tmin: 月平均最低气温,PRE为月降水量; -: 上一年的月份
3.4 树木生长与重要气候因子的动态关系
使用滑动相关分析的方法对树木生长与气候因子关系的时间动态进行了分析(图4)。其中通过总结生长与气候因子的相关分析的结果,气候因子选取了当年4月和上一年10月的平均气温,以及当年6月和上一年11月的降水量,结果表明随着气候条件的改变,3个海拔高度华北落叶松生长与气候因子的关系均不稳定。在气候因子的时间变化角度,上一年10月和当年4月平均气温在1996年之前,分别呈现下降和平稳趋势,而在1996年之后,均呈现升高趋势。上一年11月降水量呈现波动降低,6月降水量则是先升高后降低。
低海拔采样点L的华北落叶松与上一年10月平均气温的关系减弱,而与4月平均气温的关系增强,与上一年11月降水量的关系存在波动但总体维持较强的负相关水平,与当年6月降水量的关系较弱。中、高海拔采样点M和H的华北落叶松生长与气候因子关系的时间动态比较相似。二者与上一年10月平均气温的关系增强,与4月气温关系较弱,与上一年11月降水量的关系减弱,与6月降水量的关系则趋向于稳定的较强负相关关系。这些结果反映了同一气候因子的变化对于不同海拔华北落叶松生长会有不同的影响,气候条件的改变可以引起树木生长与气候因子关系,以及径向生长的海拔差异。
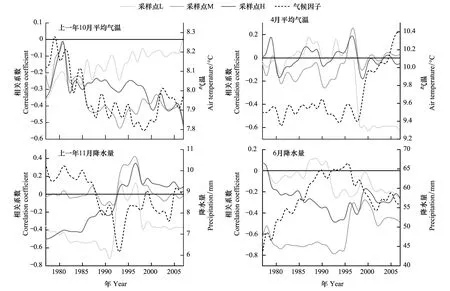
图4 不同采样点年表与主要气候因子之间的滑动相关系数Fig.4 Moving correlation coefficients calculated between chronologies and main climatic factors at different sites滑动相关和气候因子移动平均的窗口均为20a
4 讨论
4.1 华北落叶松生长与气候因子关系的海拔差异
通过不同海拔华北落叶松年表之间的相关性分析,芦芽山阳坡华北落叶松生长可以分为低海拔和中-高海拔两种年际径向生长类型,生长与气候因子关系的海拔差异也同样验证了这一点。低海拔的华北落叶松表现为与上一年11月降水量的显著负相关。11月的降水全部体现为降雪,此时降雪的覆盖会引起土壤温度的升高,从而使树木根部呼吸作用增强,消耗树木养分,对于树木春天的生长萌发不利[23]。此外,低海拔华北落叶松的生长与4月平均气温同样显著负相关,这一结果与王劲松等[24]在天山东段的研究结果较为相似。低海拔生长季开始之前气温越高会使树木呼吸作用越强,引起养分的消耗增多,从而不利于树木生长。
与低海拔不同,中-高海拔华北落叶松的生长与上一年10月的平均气温显著负相关。此时段气温偏高会引起水分蒸发,降低水分的储备,将对下一个生长季开始时树木萌发不利[24]。此外,中-高海拔的树木生长还表现为与6月降水量的显著负相关关系。在高海拔,生长季中过多的降水将引起云量过多和太阳辐射降低,从而降低光合作用的效率,致使树木生长缓慢。这一结果在许多树轮气候学研究中报道[25-26]。在芦芽山地区使用高海拔油松(Pinustabulaeformis)和白杄样芯进行的气候学研究中也同样显示了这一结果[7,27]。
4.2 增温对华北落叶松径向生长的影响
滑动相关结果表明不同海拔高度华北落叶松径向生长与气候因子的关系并不稳定,而是随着时间推移表现出一定的变化。通过前文分析,低海拔华北落叶松的生长与4月平均气温显著负相关,而中-高海拔华北落叶松生长与上一年10月平均气温显著负相关。然而滑动相关结果显示这两种负相关关系均在1958—1995年内较弱,而自1996年前后开始增强(图4),表明气温的升高导致了低海拔和中-高海拔华北落叶松生长与气温因子之间显著相关关系的出现。在低海拔,气温升高后,呼吸作用增强,生长所需养料开始消耗,对生长的不利影响开始出现。在中-高海拔,气温升高致使水分的蒸发加剧,使土壤中的水分储备不足开始影响树木生长。这证明,气温的升高引起了华北落叶松树木生长与气候因子关系的改变。
研究区气温升高引起了不同海拔高度华北落叶松生长与气候因子关系的变化,进一步引起了生长节律的海拔差异。在树木生长与气温因子的滑动相关结果中,1996年之前3个海拔高度树木生长与气温因子(上一年10月和当年4月平均气温)的关系比较相似,进入1996年之后,3个采样点与同一气温因子的相关关系开始出现分化,低海拔采样点L生长与气温因子的关系与海拔较高的M和H采样点分离(图4)。在生长节律方面,3个采样点年表之间的相关系数也是自1996年前后开始分化,中-高海拔采样点M和H年表之间相关系数维持在高水平,而低海拔采样点L年表与二者的相关系数维持在低水平(图2)。从这一角度来讲,气温升高引起了芦芽山阳坡华北落叶松径向生长节律的海拔差异。但是,这一结论仅考虑了气温条件的改变,而气温和降水以及其它环境因子之间具有复杂的交互影响和非线性关系。本研究在探索树木生长与气候因子关系的动态过程方面取得一定的结果,而在响应机理和结果验证方面还存在挖掘的空间。
5 讨论
(1)芦芽山阳坡3个海拔高度华北落叶松年表的样本总体解释量(EPS)均超过0.85阈值,是研究树木生长与气候因子关系的可靠资料。
(2)华北落叶松的年际径向生长变化具有海拔差异,海拔2440 m和2540 m的径向生长相关性较高,而与海拔2330 m的径向生长相关系数较低。
(3)华北落叶松与气候因子的关系具有海拔差异。低海拔(2330 m)华北落叶松生长与4月平均气温和上一年11月降水量显著负相关。中海拔(2440 m)和高海拔(2540 m)华北落叶松生长与上一年10月平均气温和6月降水量显著负相关。
(4)不同海拔高度华北落叶松生长与气候因子的关系均不稳定。气温升高引起了华北落叶松生长与气温因子显著相关关系的出现,同时引起了生长与气温因子的海拔差异和华北落叶松径向生长节律的海拔差异。
[1] 吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990.
[2] Fritts HC, Smith DG, Cardis JW, Budelsky CA. Tree-ring characteristics along a vegetation gradient in northern Arizona. Ecology, 1965, 46(4): 394-401.
[3] Liu Y, Cai Q F, Park W K, An Z S, Ma L M. Tree-ring precipitation records from Baiyinaobao, Inner Mongolia since A. D. 1838. Chinese Science Bulletin, 2003, 48(11): 1140-1145.
[4] Zhang Q B, Hebda R J. Variation in radial growth patterns ofPseudotsugamenziesiion the central coast of British Columbia, Canada. Canadian Journal of Forest Research, 2004, 34(9): 1946-1954.
[5] Gou X H, Chen F H, Yang M X, Li J B, Peng J F, Jin L Y. Climatic response of thick leaf spruce (Piceacrassifolia) tree-ring width at different elevations over Qilian Mountains, northwestern China. Journal of Arid Environments, 2005, 61(4): 513-524.
[6] Shao X M, Wang S Z, Zhu H F, Xu Y, Liang E Y, Yin Z Y, Xu X G, Xiao Y M. A 3585-year ring-width dating chronology of Qilian Juniper from the northeastern Qinghai-Tibetan Plateau.IAWA Journal, 2009, 30(4): 379-394.
[7] Zhang W T, Jiang Y, Dong M Y, Kang M Y, Yang H C. Relationship between the radial growth ofPiceameyeriand climate along elevations of the Luyashan Mountain in North-Central China. Forest Ecology and Management, 2012, 265: 142-149.
[8] Briffa K R, Schweingruber F H, Jones P D, Osborn T J, Shiyatov S G, Vaganov E A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature, 1998, 391(6668): 678-682.
[9] D′Arrigo R, Mashig E, Frank D, Wilson R, Jacoby G. Temperature variability over the past millennium inferred from Northwestern Alaska tree rings. Climate Dynamics, 2005, 24(2/3): 227-236.
[10] Daniels L D, Veblen T T. Spatiotemporal influences of climate on altitudinal treeline in northern Patagonia. Ecology, 2004, 85(5): 1284-1296.
[11] 高露双, 王晓明, 赵秀海. 长白山阔叶红松林共存树种径向生长对气候变化的响应. 北京林业大学学报, 2013, 35(3): 24-31.
[12] IPCC.Climate change 2007—the physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. Cambridge:Cambridge University Press, 2007.
[13] 张文涛, 江源, 王明昌, 张凌楠, 董满宇, 郭媛媛. 芦芽山阳坡不同海拔白杄径向生长对气候变暖的响应. 植物生态学报, 2013, 37(12): 1142-1152.
[14] 张文涛, 江源, 董满宇, 杨艳刚, 杨浩春. 芦芽山不同海拔华北落叶松径向生长与气候因子关系的研究. 北京师范大学学报: 自然科学版, 2011, 47(3): 304-309.
[15] 马子清. 山西植被. 北京: 中国科学技术出版社, 2001.
[16] 张金屯. 山西芦芽山植被垂直带的划分. 地理科学, 1989, 9(4): 346-353.
[17] Stokes M A, Smiley T L. An Introduction to Tree-ring Dating.Tucson: University of Arizona Press, 1996.
[18] Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69-78.
[19] 邵雪梅, 吴祥定. 华山树木年轮年表的建立. 地理学报, 1994, 49(2): 174-181.
[20] Kimball K D, Keifer M. Climatic comparisons with tree-ring data from montane forests: Are the climatic data appropriate? Canadian Journal of Forest Research, 1988, 18(4): 385-390.
[21] Wigley T W L, Briffa K R, Jones P D. On the average value of correlated time series, with applications in Dendroclimatology and Hydrometeorology.Journal of Climate Applied Meteorology, 1984, 23(2): 201-213.
[22] Biondi F, Waikul K. DendroClim2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers and Geosciences, 2004, 30(3): 303-311.
[23] 王亚军, 勾晓华, 刘普幸,陈发虎. 甘肃榆中200年来春季气温的变化. 中国沙漠, 2006, 26(2): 283-285.
[24] 王劲松, 李金豹, 陈发虎, 勾晓华, 彭剑峰, 刘普幸,靳立亚. 树轮宽度记录的天山东段近200a干湿变化. 冰川冻土, 2007, 29(2): 209-216.
[25] 彭剑峰, 勾晓华, 陈发虎, 张永香, 刘普幸,张永, 方克艳. 阿尼玛卿山地祁连圆柏径向生长对气候的响应. 地理学报, 2007, 62(7): 742-752.
[26] 朱海峰, 王丽丽, 邵雪梅, 方修琦. 雪岭云杉树轮宽度对气候变化的响应. 地理学报, 2004, 59(6): 863-870.
[27] 易亮, 刘禹, 宋慧明, 李强, 蔡秋芳, 杨银科, 孙军艳. 山西芦芽山地区树木年轮记录的1676AD以来5-7月温度变化. 冰川冻土, 2006, 28(3): 330-336.
Responses of radial growth inLarixprincipis-rupprechtiito climate change along an elevation gradient on the southern slope of Luya Mountain
ZHANG Wentao1,3,4, JIANG Yuan1,2,*, WANG Mingchang1, ZHANG Lingnan1, DONG Manyu1
1CollegeofResourcesScienceandTechnology,BeijingNormalUniversity,Beijing100875,China2StateKeyLaboratoryofEarthSurfaceProcessandResourceEcology,BeijingNormalUniversity,Beijing100875,China3CollegeofLifeSciences,BeijingNormalUniversity,Beijing100875,China4BeijingMunicipalResearchInstituteofEnvironmentalProtection,Beijing100037,China
Many studies have addressed the effects of climatic factors on the radial growth of trees in different habitats. However, changes in climate can modify the relationships between tree growth and climate, and this, in turn, can modify the annual growth patterns of trees. Luya Mountain, located in the north of central China, has experienced a clear increase in air temperature over the last several decades according to the findings of the Intergovernmental Panel on Climate Change. The Wuzhai County Meteorology Station has also recorded an increasing trend in the annual mean air temperature and a slight decrease in total annual precipitation from 1996 to 2007. Trees at three different elevations on the southern slopes of Luya Mountain (low, 2330 m; middle, 2440 m; and high, 2540 m) were selected for study, and tree-ring chronologies were created to study variation in radial growth response ofLarixprincipis-rupprechtiitrees to climate change along an elevation gradient. A total of 92 cores from 46 trees were collected, and ultimately three residual chronologies were created, one for each of the three elevations. Statistical analysis of the chronologies revealed the expressed population signals of all three chronologies exceeded 0.85, the minimum threshold indicating a strong climatic signal in tree ring chronologies. This implies that the three chronologies were all suitable for growth-climate relationship studies. Pearson′s correlations were calculated between the tree-ring chronologies and monthly climatic factors (monthly means of temperature, maximum temperature, and minimum temperature as well as total monthly precipitation) to investigate the climatic responses of trees. Moving correlation analysis was also applied to the tree-ring data and these monthly climatic factors. This allowed further analysis of the stability of the relationships between each pair of chronologies, and the relationships between tree growth and climate. The annual growth rhythms ofLarixprincipis-rupprechtiiand their relationships with climatic factors varied over time and among the three elevations. Similar annual growth rhythms were found at the mid-elevation (2440 m) and high elevation sites (2540 m), but these differed from those of trees at the low elevation site (2330 m). As for the relationship between tree growth and climate, growth at low elevation showed significant negative correlations with both monthly mean temperature in April and total monthly precipitation in the previous November, whereas the trees at middle and high elevations exhibited significant negative correlations with monthly mean temperature in the previous October and total monthly precipitation in June. However, none of these relationships between tree growth and climatic factors at the three elevations were stable based on the moving correlation analysis. The correlations between tree growth and the main climatic factors became significant in 1996, when the annual mean air temperature showed an abrupt increase. Similarly, the elevational variation in the relationships between tree growth and climate, as well as in the growth rhythms, both of which also are first apparent in 1996, suggest climatic warming drives elevational variation in growth-climate relationships and growth rhythms. The findings of this study build on our knowledge of elevational variation in the influence of climate change on vegetation.
climate change; tree rings; elevation gradient; Luya Mountain;Larixprincipis-rupprechtii
国家自然科学基金项目(41171067)
2014-03-14; < class="emphasis_bold">网络出版日期:
日期:2014-12-04
10.5846/stxb201403140452
*通讯作者Corresponding author.E-mail: jiangy@bnu.edu.cn
张文涛, 江源, 王明昌, 张凌楠, 董满宇.芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应.生态学报,2015,35(19):6481-6488.
Zhang W T, Jiang Y, Wang M C, Zhang L N, Dong M Y.Responses of radial growth inLarixprincipis-rupprechtiito climate change along an elevation gradient on the southern slope of Luya Mountain.Acta Ecologica Sinica,2015,35(19):6481-6488.
猜你喜欢
中国农业科技导报(2024年1期)2024-04-29 00:42:04
世界建筑(2022年10期)2022-10-25 08:26:48
今日农业(2021年10期)2021-11-27 09:45:24
景德镇陶瓷(2021年1期)2021-03-24 05:48:02
艺术品鉴(2019年12期)2020-01-18 08:47:30
山东林业科技(2019年2期)2019-06-03 10:10:54
现代营销(创富信息版)(2018年2期)2018-08-15 00:45:27
西南农业学报(2016年6期)2016-04-16 05:13:01
湖北农业科学(2014年21期)2014-12-22 11:42:23
自然资源遥感(2014年3期)2014-02-27 11:56:35
