
基于地理种源的刨花楠苗木比叶面积与叶片化学计量学关系
2015-01-19 07:06徐朝斌钟全林程栋梁胡松竹伍伯妍孙晓媚张佩生
生态学报 2015年19期
徐朝斌, 钟全林,*, 程栋梁, 胡松竹, 胡 波, 伍伯妍, 孙晓媚, 张佩生
1 福建师范大学地理科学学院, 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 2 江西农业大学林学院, 南昌 330045
基于地理种源的刨花楠苗木比叶面积与叶片化学计量学关系
徐朝斌1, 钟全林1,*, 程栋梁1, 胡松竹2, 胡 波1, 伍伯妍1, 孙晓媚1, 张佩生1
1 福建师范大学地理科学学院, 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 2 江西农业大学林学院, 南昌 330045
探讨植物比叶面积(SLA)与叶片碳(C)、氮(N)、磷(P)化学计量学关系,能够反映植物为获取最大光合生产所采取的内部调控机制,共同体现植物的适应策略。利用生长于同一土壤与气候环境中培育的刨花楠(Machiluspauhoi)1年生苗木,对其SLA与叶片C、N、P含量进行测定,并对SLA与叶片C、N、P化学计量学特征及其与种源地环境因子的关系进行分析。结果表明:(1)叶片养分含量的变异系数大小排序为C < N < P;SLA与叶片N、P含量呈显著的正相关,与叶片C∶N及C∶P呈极显著的负相关。(2)SLA与经度、年均温、年降水量呈显著负相关;叶片C、N、P含量也受种源地环境因子影响,其中以海拔最为重要。研究结果有助于理解刨花楠苗木的生存适应对策,对探究刨花楠对养分的资源利用效率等具有重要意义。
比叶面积; 叶片碳氮磷含量; 化学计量学; 刨花楠
植物的叶片性状(包括生态和生理性状)能够反应植物适应环境变化的生存对策[1-2],例如,净光合速率、比叶面积(叶片面积和质量的比值,SLA)、叶寿命、叶片碳(C)、氮(N)和磷(P)含量等,都是植物为获得最大光合生产所形成的适应策略[3-6]。其中,SLA与植物的生长和生存对策有紧密的联系,能反映植物对不同生境的适应特征[7-10];C、N、P元素(主要生源要素)是植物生长所必需的大量元素[11-14],C元素是结构性物质[15],N、P元素是植物生长的限制元素[16]。探讨植物SLA与叶片C、N、P化学计量学关系,能够反映植物为获取最大C收获所采取的内部调控机制,具有重要的生态学和生物进化意义[17]。
刨花楠(Machiluspauhoi)为亚热带常绿乔木阔叶树种,是珍贵的用材树种和优美的庭园观赏树种[18]。鉴于市场对该树种资源的需求量日益增加,对其研究也越来越多,研究内容主要集中在其生长特性[19-20]、木材特性[21]、生物量[22-23]、人工扦插繁殖及培育技术[24-26]、光响应特性[27]、外界因素对生长的影响[28-29]等方面,但暂未对其SLA与叶片C、N、P化学计量学的关系进行研究。为此,本文通过对刨花楠SLA及其C、N、P含量的测定与分析,阐明其SLA与叶片C、N、P化学计量学关系,并对其生态适应机制进行分析,研究结果将有助于理解刨花楠生存适应策略,对探究刨花楠对养分的资源利用效率等也具有重要意义。
1 材料与方法
1.1 研究地概况
试验样地位于江西省南昌市江西农业大学苗圃园(115°49′59″E,28°45′35″ N),海拔约44 m。实验地属中亚热带湿润季风气候,气候湿润温和,日照充足,一年中夏冬季长,春秋季短。夏天非常炎热,冬天较寒冷。年平均气温17—17.7 ℃,极端最高气温40.9 ℃,极端最低气温-15.2 ℃,≥10 ℃的年积温为4480—4590 ℃。年降雨量1600—1700 mm,降水日为147—157 d,年平均暴雨日5.6 d,年平均相对湿度为78.5%。年日照时间1723—1820 h,日照率为40%。年无霜期251—272 d。冬季多偏北风,夏季多偏南风。试验地地势平坦,光照充足,排水通畅。
1.2 供试材料
试验材料为1年生刨花楠苗木,其种子分别产自江西崇义(CY)、吉安(JA)、遂川(SC)、泰和(TH)和永新(YX)5个县。5个县区的试验材料集中于试验地生长时间为1a,苗木株行距为5cm×20 cm,能够满足苗木的正常生长。对试验地苗木实行统一的常规管理,所施肥料为尿素 (CO(NH2)2)与五氧化二磷(P2O5),其中尿素施用量为300 kg/hm2,P2O5为64.12 kg/hm2(即N∶P比为5∶1),于生长季3—6 月期间每20 d施1次。于2010年9月对试验地(土壤为红壤)幼苗生长情况进行调查。随机选取生长状况中等的刨花楠幼苗66 株(分别种源选取,具体样本数见表2),测量其冠幅、株高、地径,地径用游标卡尺测量,株高用钢卷尺测量,冠幅(分南北和东西方向测量,取平均值)。样木平均地径为(0.397±0.009) cm,平均株高为(26.29±0.68) cm,平均冠幅为(22.26±0.50) cm。对所取样木各选取3片完全伸展、没有病虫害、大小中等的叶片,用LI-3100测量其叶面积(分别测量5次取平均值),用奥豪斯公司生产的电子天平测量其叶重(先在烘箱内经105℃杀青30 min,然后再在70 ℃下烘干至恒重后称,精确到0.001 g)。为防止用于测定叶片C、N、P含量的样品质量不足,对每1株苗木另采摘大小中等、无病虫害的相同叶位的完全伸展叶片2 片,按上述同样方法烘干至恒重。将烘干的样叶研磨并过1 mm筛,用于叶片C、N、P含量的测定。
1.3 测定方法
1.3.1 碳氮磷含量的测定
利用CHNOS元素分析仪(Elemental Analyzer Vario EL III, Germany。)分析得出C、N值,采用H2SO4-H2O2消煮-钼锑抗比色法(参见《土壤农化分析》[30]测定P值。
1.3.2 比叶面积的测定
利用便携式叶面积仪(LI-3100,America。)测量其叶面积,并测定其干质量为叶干重, SLA按公式计算,即:
SLA(m2/kg)=叶面积(m2)/叶干重(kg)
1.4 数据处理
所有统计分析与作图均采用Excel 2003 和SPSS 17.0 软件完成。
2 结果与分析
2.1 刨花楠苗木叶片SLA和养分(C、N、P)化学计量学特点
SLA与植物的生长对策及植物利用资源的能力紧密相关,能够提供植物生长和适应环境的重要信息,且具有易测定的特点[3]。叶片经济学谱(Leaf Economic Spectrum)认为高的SLA表明叶片薄,N含量高,光合速率高,叶寿命短;反之,厚的叶片N含量较低,光合速率较低,而叶寿命长[3]。植物C、N、P生态化学计量学(Ecological stoichiometry)理论为研究植物的养分利用状况提供了重要的手段,植物能够主动地调整养分需求以适应环境变化[31]。C、N、P 三者之间的化学计量比能够反映植物的C积累状态和N、P 养分限制格局,并成为植物生长、生理过程发生变化的内在驱动因素之一[32]。由表1可知,苗木SLA具有较大变异性,变化范围在11.93—35.49 m2/kg之间。叶片C、N、P 含量、C∶N、C∶P和N∶P的变异系数较小。
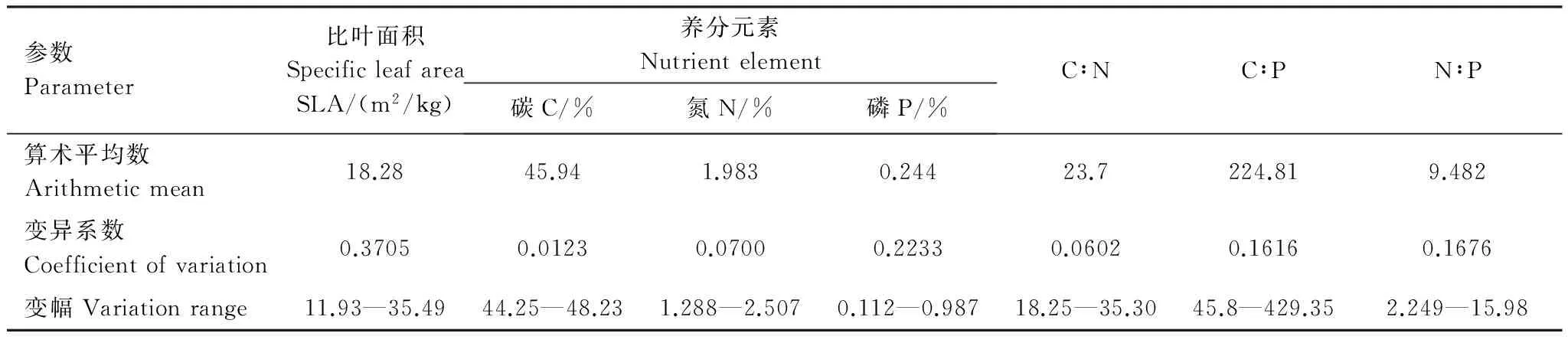
表1 观测样木叶片性状的统计特征(n=66)
2.2 叶片C、N、P含量的相关性
叶片中C、N、P3种元素含量的相关性分析表明,N、P含量间呈极显著的正相关(P<0.01)(图1),表明刨花楠叶片N含量和P含量具有一定的相互促进作用,但其相关系数R2较小,可能是由于样本种子来源地较多,不同种源地的局部地理环境(如土壤养分、温度、降水等等)差异对其有重要影响。
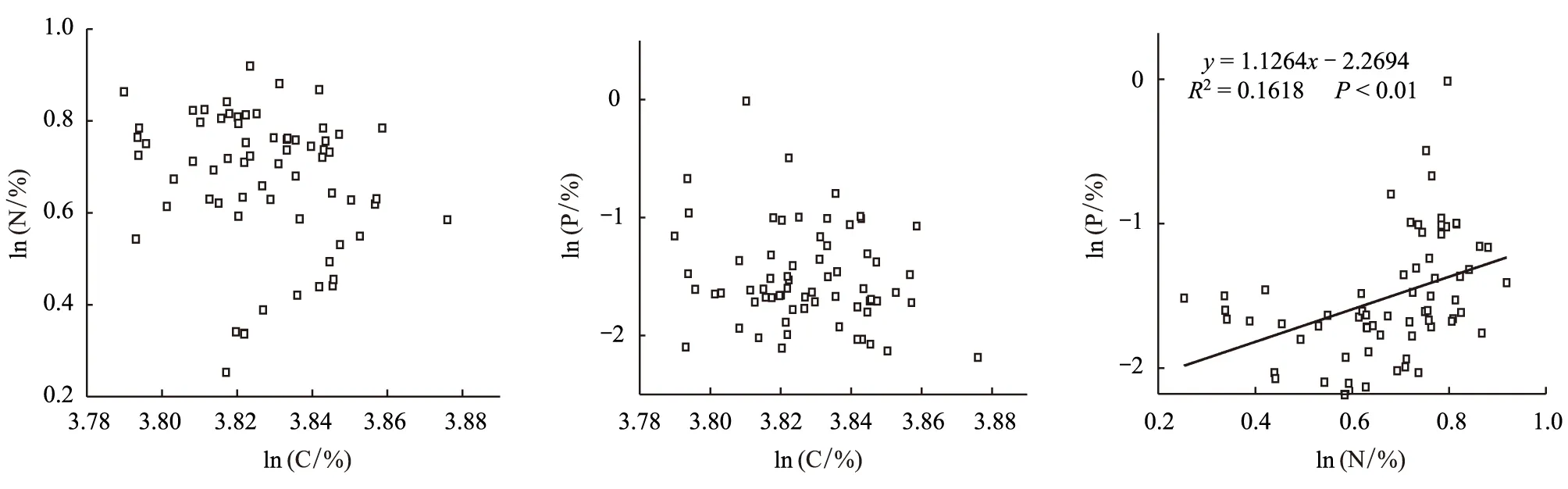
图1 刨花楠苗木叶片碳、氮、磷含量的关系(n=66)Fig.1 Correlation coefficients among Leaf C, N, P concentration of Machilus pauhoi
2.3 叶片SLA与C、N、P含量的相关性
植物对生源要素(C、N、P)具有稳定的生理代谢需求,可以通过叶片性状(如SLA)与外界环境进行物质交换,进而反馈于自身的化学计量特征[33],SLA与植物的生理代谢和生长策略密切相关[15],是连接植物生理和生长环境的重要纽带。对各不同种源刨花楠SLA与C、N、P含量进行相关性分析发现,仅CY种源SLA与C含量呈极显著负相关(P<0.01)、SC种源SLA与N含量呈极显著正相关(P<0.01)(表2),分析原因,可能是受种源内采种母树间相隔距离远近所影响,距离较远的各母树种子所发育形成的苗木叶片间的叶面积、叶片质量与养分含量由于受局部土壤养分含量差异影响而相差较大,这也反映了其对环境适应的生长策略。
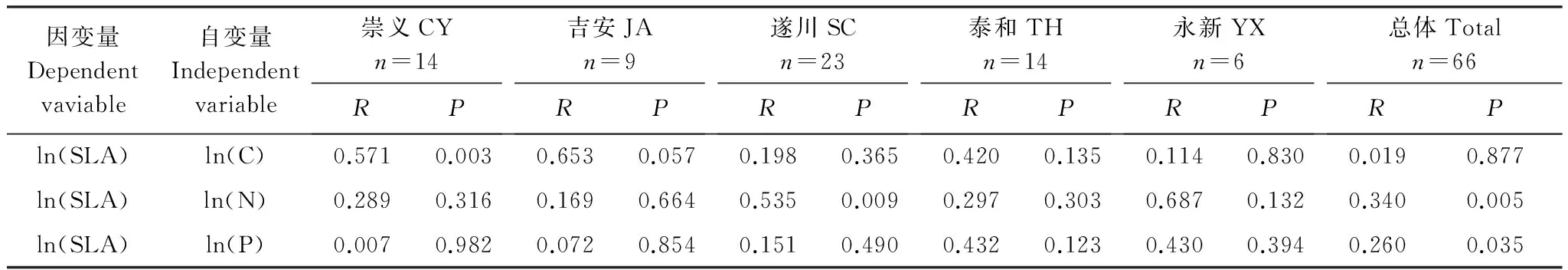
表2 不同种源刨花楠比叶面积和叶片C、N、P含量的线性相关量列表
CY, JA, SC, TH, YX mean Chongyi, Ji′an, Suichuan, Taihe, Yongxin county of Jiangxi province, respectively
图2可看出总体刨花楠苗木SLA与N、P含量呈正相关,其中SLA与N含量之间呈极显著相关性(P<0.01),与P含量之间呈显著相关性(P<0.05),但与叶片C含量之间的相关性均未达到显著水平(P>0.05)(表2)。随着SLA的增大,叶片N、P含量呈现出上升的趋势。从两模型方程的指数看,N的变化要小于P,因为幼苗时期的刨花楠核糖体快速地合成蛋白质支持其快速生长,导致分配到rRNA的P增加[34]。生长速率假说认为生物体的生长受到RNA含量以及依靠RNA合成的蛋白质的双重限制, 已有研究证明SLA值大的植物叶片N含量高[3]。
2.4 叶片SLA与C∶N、C∶P、N∶P的相关性
植物体按一定的比例关系吸收利用各种必需元素(C、N、P), 并在体内相互耦合保持相对平衡[35]。其C、N、P化学计量比与叶片性状(如SLA等)形成了叶片稳态机制[36]。由表3可知,5种源刨花楠中仅SC种源SLA与叶片C∶N呈极显著正相关(P<0.01)。从图3可看出,刨花楠苗木SLA与叶片C∶N、C∶P的呈显著负相关(P<0.05),但与N∶P的相关性不显著(P>0.05)(表3)。
2.5 叶片SLA及C、N、P化学计量学与种源地环境因子的关系
了解种源差异与地理气候的关系, 有助于揭示影响地理变异的主导自然因子。SLA与经度、年均温、年降水量呈显著负相关(P<0.05)。表明刨花楠SLA呈经向地带性规律,随温度与降水量增加而减小。
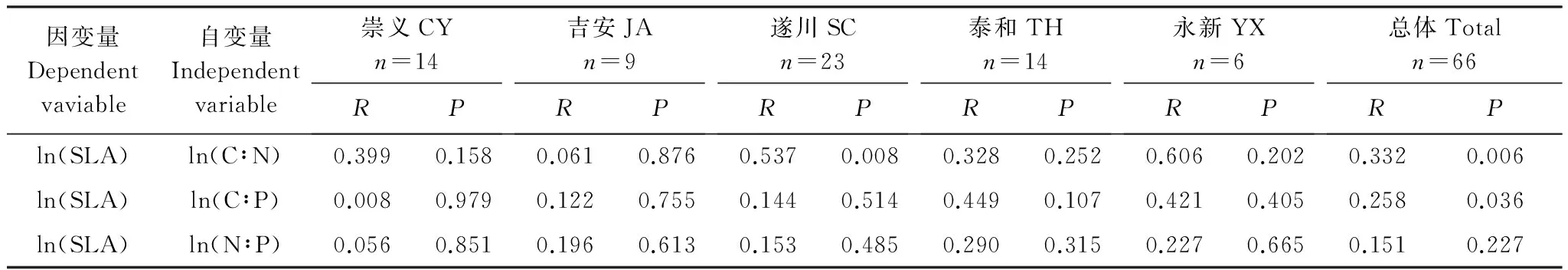
表3 不同种源刨花楠比叶面积和叶片C∶N、C∶P与N∶P比的线性相关量列表
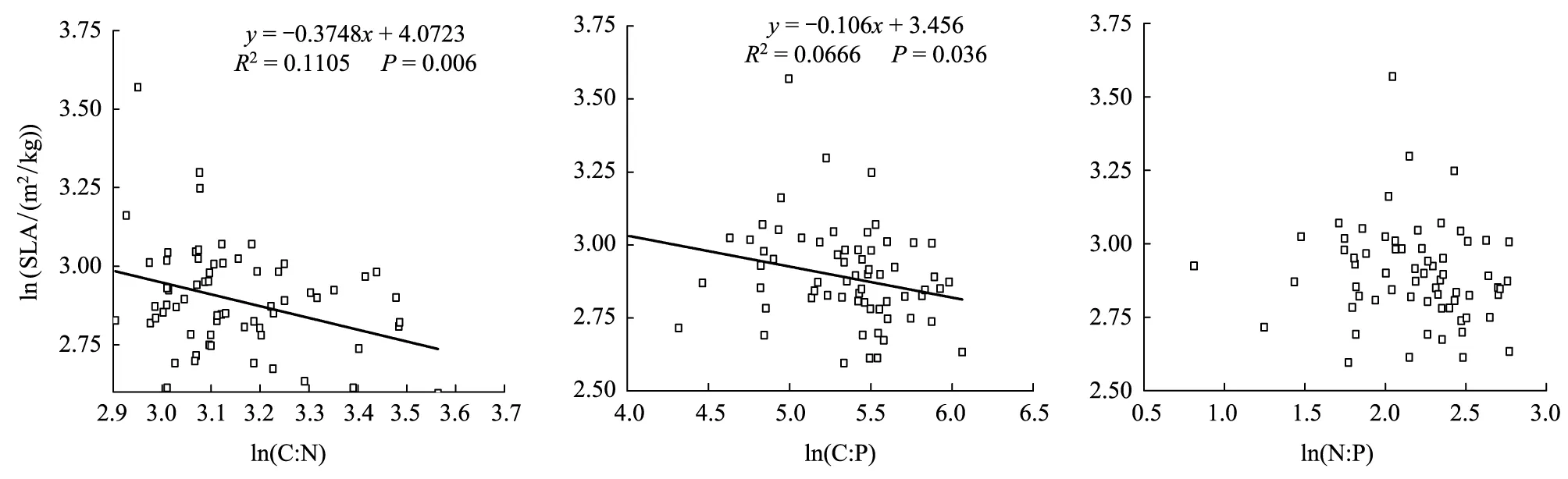
图3 刨花楠苗木SLA和叶片C∶N、C∶P与N∶P比的关系(n=66)Fig.3 Correlation coefficients among SLA and Leaf C∶N, C∶P, N∶P of Machilus pauhoi
叶片C含量与经度、纬度及年无霜期呈显著正相关(P<0.05),与海拔呈极显著负相关(P<0.01)。叶片N、P含量与海拔呈显著正相关(P<0.05),与经度、年无霜期呈显著负相关(P<0.05);另叶片P含量与纬度、年均温呈显著负相关(P<0.05)。
叶片C∶N、C∶P与经度及年无霜期呈显著正相关(P<0.05),与海拔呈显著负相关(P<0.05),另C∶P与纬度、年均温也呈显著正相关(P<0.05)。叶片N∶P与经度、纬度、年均温呈显著正相关(P<0.05)。
3 讨论与结论
植物叶片性状能反映植物适应环境的生长策略和利用资源能力,其中植物SLA是生态学研究中重要的叶片性状指标[1]。从表1可看出,不同种源地的刨花楠种子在同一生长环境下生长1a后,其SLA及C、N、P含量等主要性状指标以SLA变异为大,可能是刨花楠个体的SLA受种源地环境影响相对较为敏感,产生了资源利用效率差异所致[1],反映了其生长策略和生活史的多样性。
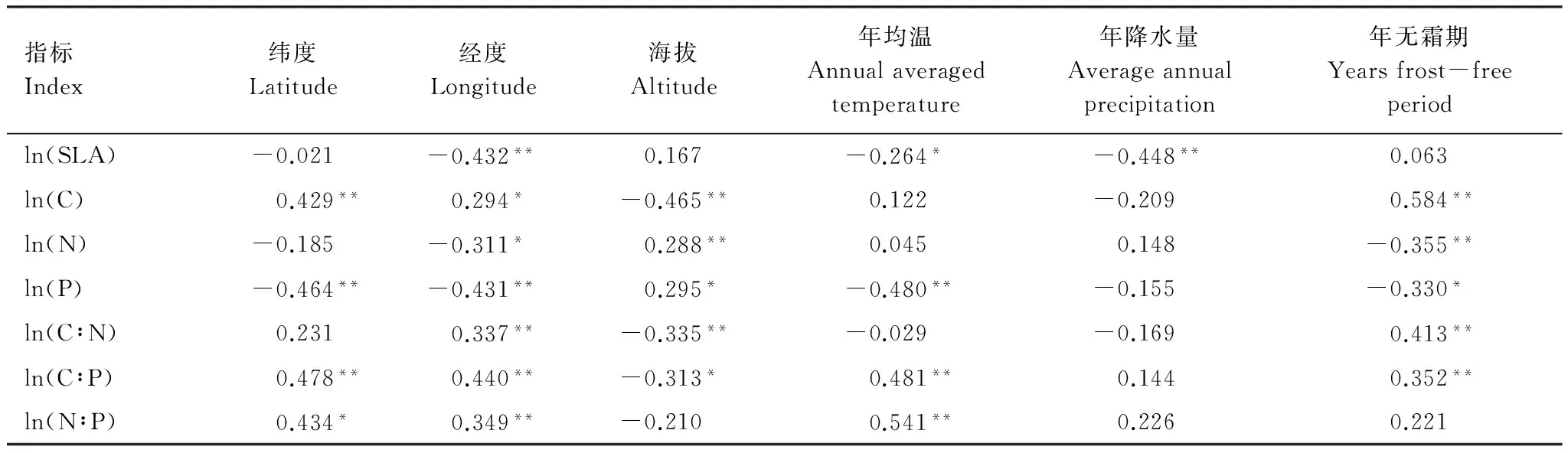
表4 叶片性状与种源地环境因子的相关性(n=66)
植物叶片对环境变化敏感且可塑性大,其营养元素组成反映植物的生态策略,也是适应生境条件的一种表征[37]。叶片养分含量的变异系数大小排序为C < N < P(表1),这是因为C元素是较稳定的结构性物质,N、P元素是植物生长的限制元素。研究还发现,刨花楠苗木叶片N含量和P含量具有一定的相互促进作用,因为N可以合成蛋白质,有利于植物根系生长,故对植物吸收利用P有利;P可以促进植物蛋白质的合成,故有利于植物对N的吸收利用[38]。三者以叶片P含量为其营养组成(主要生源要素)最主要的变动因素,高生长速率的个体需要更多的rRNA投入以生产生长所需的蛋白质,而rRNA是植物的一个主要P库,rRNA含量增加将导致植物细胞中P浓度增加[39],但叶片生态化学计量学特征也会随着季节更替和生长阶段而不断变化,一次性采样分析不能反映其生态化学计量学特征变化动态, 全面的、跨越时空的采样更加科学、合理,有待后续深入研究[40-41]。
植物叶片性状因遗传特性和环境条件不同而异,即使在相同生境内的同一物种也会因植物个体生存策略和资源利用效率不同表现出差异性。因此通过研究刨花楠SLA和C、N、P含量关系来讨论刨花楠的生态适应机制。一般认为,植物SLA能够反映植物对C的获取与利用的平衡关系[42],但本研究发现刨花楠SLA与叶片C含量呈微弱的负相关(图2,表2),导致这种现象的可能原因:一是与幼苗的呼吸作用密切相关,幼苗时期大部分通过光合作用合成的碳水化合物,为了给蛋白质合成过程提供能量,被呼吸作用消耗,致使幼苗叶片C含量减少;二是土壤中的N、P元素影响和限制着植物的生长状况和生产力[43]。本研究中刨花楠SLA和N、P含量呈显著正相关(图2,表2),说明具有较高养分含量的叶片,其光合生产能力较强,导致其SLA较大[3],与韦兰英和上官周平[44]、Wright等[45]、Reich等[46]的研究结果一致。另外,SLA越大的叶片,其厚度或密度相对较小[47-48],便于光强度到达叶片内部叶绿体,减少了CO2在叶肉组织中的传导阻力,缩短了气体在叶内的传导距离[49],可能提高叶片N向光合器官的分配比例[50-52]。综合来看,N和P对刨花楠SLA的影响要大于C对刨花楠SLA的影响。
随着对单个营养元素研究的深入,越来越多学者开始研究元素的耦合关系。其中C、N、P化学计量比为研究C、N、P耦合过程提供了很好的思路。三者之间较为稳定的化学计量比是植物维持其生长和生理过程井然有序的基本前提[40-41,53]。植物叶片C∶N和C∶P反映的是植物叶片C与N、P相对协调能力,预示着植物吸收营养所能同化C的能力[40],植物体中的N∶P化学计量特征可以体现N和P两种营养元素的供给状况相对有效性[54]。刨花楠苗木SLA和叶片C∶N、C∶P、N∶P化学计量比呈负相关(图3),且与叶片C∶N、C∶P的相关性达到显著性水平(P<0.05),这与本研究结果(SLA和N、P含量呈显著正相关)相一致,这是由于幼苗时期刨花楠对各营养元素吸收不同步[55-58],C相对较稳定[15],N、P是受环境影响较大的生长元素[16],N和P对SLA的影响要大于C。本研究表明SLA与叶片养分结构特征存在较为稳定的关系,这是刨花楠对环境适应策略的体现。
种源地地理气候的差异可能是刨花楠SLA和养分组成发生变化的重要原因,但调控植物叶性特征的机制较为复杂。SLA呈经向地带性规律,随温度与降水量增加而减小,反映了对热量负荷、湿度的适应策略。叶片C、N、P化学计量学与各地理气候因子的关系密切,表明太阳辐射、相对湿度、平均温度和蒸发量等气候因子是影响刨花楠种子的重要因素。为进一步了解各气候因子对叶片性状影响的权重,经主成分分析发现海拔为第一个主成分,本研究中叶片N、P含量与海拔呈显著正相关(P<0.05),这可能是高海拔植物叶片对其生存环境的一种适应方式[59],即在高海拔地区土壤有效N含量降低,植物为了适应“低N”的生存环境,其用于光合生产的N反而会增加,是一种遗传适应性的表现。高海拔的植物为了适应高海拔的不良环境,其体内含有更高含量的蛋白质、脂肪、淀粉[60]和黄酮类物质[61],叶片紫外吸收物质含量也随海拔增加[62],导致其生理功能的强化和物质代谢的改善,即植物的特异性适应性变异。由于刨花楠种子存在结实不稳、结实率低、且通常2—3a结果1次等特点,当年仅搜集到江西5种源地的种子,未搜集到浙江、湖南与广东等省份的种源种子,因此关于刨花楠SLA和养分组成对种源地地理气候的生态适应对策仍需进行深入研究。本文也仅限于刨花楠苗木的研究,随着其个体的生长发育,其SLA与叶片C、N、P化学计量学关系可能会发生变化。
[1] Vendramini F, Diaz S, Gurvich D E, Wilson P J, Thompson K, Hodgson J G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytologist, 2002, 154(1): 147-157.
[2] Westoby M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil, 1998, 199(2): 213-227.
[3] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T L, Lee W, Lusk C, Midgley J J, Navas M, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821-827.
[4] 黄海侠, 杨晓东, 孙宝伟, 张志浩, 阎恩荣. 浙江天童常绿植物当年生与往年生叶片性状的变异与关联. 植物生态学报, 2013, 37(10): 912-921.
[5] 何春霞, 李吉跃, 孟平, 张劲松. 4种高大树木的叶片性状及WUE随树高的变化. 生态学报, 2013, 33(18): 5644-5654.
[6] 王丽芳, 王德轩, 上官周平. 大穗型小麦叶片性状、养分含量及氮素分配特征. 生态学报, 2013, 33(17): 5219-5227.
[7] 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 2005, 25(2): 304-311.
[8] Meziane D, Shipley B. Interacting determinants of specific leaf area in 22 herbaceous species: effects of irradiance and nutrient availability. Plant, Cell & Environment, 1999, 22(5): 447-459.
[9] Poorter H, de Jong R. A comparison of specific leaf area, chemical composition and leaf const ruction costs of field plants from 15 habitats differing in productivity. New Phytologist, 1999, 143(1): 163-176.
[10] Garnier E, Shipley B, Roumet C, Laurent G. A standardized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology, 2001, 15(5): 688-695.
[11] 杨佳佳, 张向茹, 马露莎, 陈亚南, 党廷辉, 安韶山. 黄土高原刺槐林不同组分生态化学计量关系研究. 土壤学报, 2014, 51(1): 133-142.
[12] 陈军强, 张蕊, 侯尧宸, 马丽娜, 丁路明, 龙瑞军, 尚占环. 亚高山草甸植物群落物种多样性与群落C、N、P生态化学计量的关系. 植物生态学报, 2013, 37(11): 979-987.
[13] 洪江涛, 吴建波, 王小丹. 全球气候变化对陆地植物碳氮磷生态化学计量学特征的影响. 应用生态学报, 2013, 24(9): 2658-2665.
[14] 刘兴华, 陈为峰, 段存国, 王凯, 王海翠. 黄河三角洲未利用地开发对植物与土壤碳、氮、磷化学计量特征的影响. 水土保持学报, 2013, 27(2): 204-208.
[15] Agren G I. Stoichiometry and nutrition of plant growth in nature communities. Annual Review of Ecology, Evolution, and Systematics, 2008, 39: 153-170.
[16] Elser J J, Bracken M E S, Cleland E E, Gruner D S, Harpole W S, Hillebrand H, Ngai G T, Seabloom E W, Shurin J B, Smith J E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 2007, 10(12): 1135-1142.
[17] 刘福德, 王中生, 张明, 王文进, 安树青, 郑建伟, 杨文杰, 张世挺. 海南岛热带山地雨林幼苗幼树光合与叶氮、叶磷及比叶面积的关系. 生态学报, 2007, 27(11): 4651-4661.
[18] 徐朝斌, 钟全林, 程栋梁, 胡松竹, 常云妮, 张治, 胡波. 基于C2H和D2H的刨花楠幼苗生物量回归模型比较. 华南农业大学学报, 2013, 34(4): 543-547.
[19] 钟全林, 胡松竹, 黄志强, 肖水清, 程建华. 刨花楠生长特性及其生态因子影响分析. 林业科学, 2002, 38(2): 165-168.
[20] 廖龙泉. 刨花楠生长规律的初步研究. 江苏林业科技, 1997, 24(1): 39-41.
[21] 郭晓敏, 牛德奎, 孙科辉. 优良阔叶树种-刨花楠木材构造性质及用途的研究. 江西农业大学学报, 1999, 21(3): 391-394.
[22] 钟全林, 张振瀛, 张春华, 周海林, 黄志强. 刨花楠生物量及其结构动态分析. 江西农业大学学报, 2001, 23(4): 533-536.
[23] 邓仕坚, 廖利平, 汪思龙, 高洪, 林柏. 湖南会同红栲-青冈-刨花楠群落生物生产力的研究. 应用生态学报, 2000, 11(5): 651-654.
[24] 胡松竹, 钟全林, 黄志强, 肖水清, 程建华. 刨花楠人工栽培技术初探. 江西农业大学学报, 2001, 23(3): 332-335.
[25] 何贵平, 黄海泳, 骆文坚, 季新良, 冯建国, 冯建民. 刨花楠、花梨木、乐东拟单性木兰嫩枝扦插繁殖试验. 浙江林业科技, 2004, 24(3): 30-32.
[26] 徐奎源, 徐永星, 徐裕良. 红楠等4种楠木树种的栽培试验. 江苏林业科技, 2005, 32(2): 26-27.
[27] 钟全林, 程栋梁, 胡松竹, 贺利中, 唐承财, 文雅香, 邱剑飞, 李秀花. 刨花楠和华东润楠叶绿素含量分异特征及与净光合速率的关系. 应用生态学报, 2009, 20(2): 271-276.
[28] 鲁美娟, 江洪, 李巍, 余树全, 曾波, 蒋馥蔚, 李佳, 金清. 模拟酸雨对刨花楠幼苗生长和光合生理的影响. 生态学报, 2009, 29(11): 5986-5994.
[29] 汪炎明. 杉木萌芽林中套中刨花楠经营技术及生长效应. 林业科技开发, 2009, 23(5): 119-121.
[30] 史瑞和. 土壤农化分析 (第二版). 北京: 农业出版社, 1992.
[31] 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 2005, 29(6): 1007-1019.
[32] 羊留冬, 杨燕, 王根绪, 郭剑英, 杨阳. 短期增温对贡嘎山峨眉冷杉幼苗生长及其C N P 化学计量学特征的影响. 生态学报, 2011, 31(13): 3668-3676.
[33] 闫帮国, 何光熊, 李纪潮, 纪中华. 金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系. 植物生态学报, 2012, 36(11): 1136-1144.
[34] 银晓瑞, 梁存柱, 王立新, 王炜, 刘钟龄, 刘小平. 内蒙古典型草原不同恢复演替阶段植物养分化学计量学. 植物生态学报, 2010, 34(1): 39-47.
[35] 秦海, 李俊祥, 高三平, 李铖, 李蓉, 沈兴华. 中国660种陆生植物叶片8种元素含量特征. 生态学报, 2010, 30(5): 1247-1257.
[36] Sterner R W, Elser J J. Ecological Stoichiometry: the Biology of Elements from Molecules to the Biosphere. Princeton: Princeton University Press, 2002: 906-907.
[37] 邓蕾, 王鸿喆, 上官周平, 刘广全. 水蚀风蚀交错区柠条锦鸡儿叶片比叶面积和营养元素变化动态. 生态学报, 2010, 30(18): 4889-4897.
[38] 蔡艳, 张毅, 刘辉, 张锡洲. 峨眉山常绿阔叶林常绿和落叶物种叶片C、N、P研究. 浙江林业科技, 2009, 29(3): 9-13.
[39] Gorokhova E, Kyle M. Analysis of nucleic acids in daphnia: development of methods and ontogenetic variations in RNA-DNA content. Journal of Plankton Research, 2002, 24(5): 511-522.
[40] 吴统贵, 陈步峰, 肖以华, 潘勇军, 陈勇, 萧江华. 珠江三角洲3 种典型森林类型乔木叶片生态化学计量学. 植物生态学报, 2010, 34(1): 58-63.
[41] 吴统贵, 吴明, 刘丽, 萧江华. 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化. 植物生态学报, 2010, 34(1): 23-28.
[42] Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 1999, 143(1): 155-162.
[43] 宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692-3703.
[44] 韦兰英, 上官周平. 黄土高原不同退耕年限坡地植物比叶面积与养分含量的关系. 生态学报, 2008, 28(6): 2526-2535.
[45] Wright I J, Reich P B, Westoby M. Strategy shifts in leaf physiology, structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats. Functional Ecology, 2001, 15(4): 423-434.
[46] Reich P B, Uhl C, Walters M B, Ellsworth D S. Leaf lifespan as a determinant of leaf structure and function among 23 Amazonian tree species. Oecologia, 1991, 86(1): 16-24.
[47] Niinemets Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology, 2001, 82(2): 453-469.
[48] 刘金环, 曾德慧, Lee D K. 科尔沁沙地东南部地区主要植物叶片性状及其相互关系. 生态学杂志, 2006, 25(8): 921-925.
[49] Parkhurst D F. Diffusion of CO2and other gases inside leaves. New Phytologist, 1994, 126(3): 449-479.
[50] Poorter H, Evans J R. Photosynthetic nitrogen-use efficiency of species that differ inherently in specific leaf area. Oecologia, 1998, 116(1/2): 26-37.
[51] Feng Y L, Fu G L, Zheng Y L. Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation and use efficiencies between invasive and noninvasive alien congeners. Planta, 2008, 228(3): 383-390.
[52] Feng Y L, Lei Y B, Wang R F, Callaway R M, Valiente-Banuet A, Inderjit, Li Y P, Zheng Y L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(6): 1853-1856.
[53] Schimel D S. All life is chemical. BioScience, 2003, 53(5): 521-524.
[54] 邬畏, 何兴东, 周启星. 生态系统氮磷比化学计量特征研究进展. 中国沙漠, 2010, 30(2): 196-301.
[55] Fichtner K, Schulze E D. The effect of nitrogen nutrition on growth and biomass partitioning of annual plants originating from habitat s of different nitrogen availability. Oecologia, 1992, 92(2): 236-241.
[56] McDonald A J S, Lohammar T, Ingestad T. Net assimilation rate and shoot area development in birch (BetulapendulaRoth.) at different steady-state values of nutrition and photon flux density. Trees, 1992, 6(1): 1-6.
[57] 李凯, 项文化. 湘中丘陵区12个树种比叶面积、SPAD值和种子干质量的比较. 中南林业科技大学学报, 2011, 31(5): 213-218.
[58] 黄菊莹, 袁志友, 李凌浩. 羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化. 植物生态学报, 2009, 33(3): 442-448.
[59] Hultine K R, Marshall J D. Altitude trends in conifer leaf morphology and stable carbon isotope composition. Oecologia, 2000, 123(1): 32-40.
[60] 韩发, 贲桂英, 师生波. 青藏高原不同海拔矮嵩草蛋白质、脂肪和淀粉含量的变异. 植物生态学报, 1997, 21(2): 97-104.
[61] 汪海峰, 鞠兴荣, 何广斌, 靳晓秋, 陈剑. 不同海拔高度和生长季节对银杏叶中黄酮苷含量的影响. 林产化学与工业, 2002, 22(4): 47-50.
[62] 彭慧超, 韩发, 王环, 师生波, 沈建伟, 周党卫. 3种高山植物紫外吸收物质含量随海拔与季节的变化特征. 西北植物学报, 2010, 30(6): 1197-1203.
Variation in relationships between SLA and Leaf C, N, P stoichiometry inMachiluspauhoiamong locations
XU Chaobin1, ZHONG Quanlin1,*, CHENG Dongliang1, HU Songzhu2, HU Bo1, WU Boyan1, SUN Xiaomei1, ZHANG Peisheng1
1StateKeyLaboratoryBreedingBaseofHumidSubtropicalMountainEcology,CollegeofGeographicalSciences,FujianNormalUniversity,Fuzhou350007,China2CollegeofForestry,JiangxiAgriculturalUniversity,Nanchang330045,China
Machiluspauhoiis a native subtropical forest hardwood species. Due to its ecological and economic importance and its growing market demand, it is necessary to explore the survival strategies ofM.pauhoi. Specific leaf area (SLA), leaf nitrogen, and phosphorus concentration are closely associated with plant performance. For example, SLA is closely related to plant growth and survival strategies. The elements C, N, and P are mainly biogenic elements in plants and relatively stable leaf C, N, and P stoichiometry is fundamental to plant growth and physiological processes. C is a structural material, while N and P are the limiting elements for the plant growth. Therefore, the relationships between leaf nutrients (i.e., C, N, P concentration) and SLA are of considerable interest to researchers attempting to understand nutrient resource use efficiency, as well as to those interested in plant ecological adaption and survival strategies. To understand the mechanisms underlying variation in leaf nutrients and SLA among geographic locations, one-year old seedlings ofM.pauhoiwere collected from five counties in Jiangxi Province. Leaf C, N, P stoichiometry, and SLA were compared and the relationships between these traits and associated environmental factors analyzed. The results indicated that across the fiveM.pauhoipopulations, mean leaf C, N, and P concentrations were 45.94%, 1.983% and 0.244% respectively. Leaf C concentration had the lowest coefficient of variation (0.0123), whereas leaf P concentration had the highest coefficient of variation (0.2233). Mean values of C/N, C/P, and N/P were 23.7, 224.81, and 9.482, respectively, across the fiveM.pauhoipopulations. SLA was positively correlated with leaf N and P and negatively related to leaf C∶N and C∶P ratios. However, SLA showed no significant relationship with leaf C concentration, which might result from the relatively constant leaf C concentration among the five populations. Furthermore, SLA showed negative relationships with longitude, mean annual temperature and mean annual precipitation, indicating that SLA was sensitive to environmental factors. Likewise, leaf C, N, and P concentrations were also influenced by spatial variation in environmental factors. Altitude was the most important environmental factor influencing leaf nutrient variation among the fiveM.pauhoipopulations. Our study suggests that it is useful to explore the efficiency of nutrient utilization in plants, and will aid understanding of the survival strategies ofM.pauhoiseedlings. However, further studies are needed to understand the relationships between SLA and stoichiometry of leaf C, N, P which may change in later growth stages.
specific leaf area(SLA); leaf carbon, nitrogen and phosphorus concentration; stoichiometry;Machiluspauhoi
国家自然科学基金项目(31170596, 31170374, 31370589); 国家农业科技成果转化资金项目(2011GB2C400005); 福建省科技厅重大项目(2014N5008); 福建省种业创新项目(FJZY2014-01)
2014-02-07; < class="emphasis_bold">网络出版日期:
日期:2014-12-04
10.5846/stxb201402070217
*通讯作者Corresponding author.E-mail: qlzhong@126.com
徐朝斌, 钟全林, 程栋梁, 胡松竹, 胡波, 伍伯妍, 孙晓媚, 张佩生.基于地理种源的刨花楠苗木比叶面积与叶片化学计量学关系.生态学报,2015,35(19):6507-6515.
Xu C B, Zhong Q L, Cheng D L, Hu S Z, Hu B, Wu B Y, Sun X M, Zhang P S.Variation in relationships between SLA and Leaf C, N, P stoichiometry inMachiluspauhoiamong locations.Acta Ecologica Sinica,2015,35(19):6507-6515.
猜你喜欢
林产工业(2022年4期)2022-04-20
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
世界科学技术-中医药现代化(2021年12期)2021-04-19
绿色科技(2020年1期)2020-11-29
农家科技中旬版(2020年4期)2020-05-25
绿色科技(2020年1期)2020-04-19
新疆农业科学(2020年1期)2020-02-14
中国麻业科学(2015年5期)2015-12-28
创新科技(2015年9期)2015-12-15
云南中医学院学报(2014年4期)2014-07-31
