
油菜菌核病研究进展
2015-01-07 07:16丁一娟万华方梅家琴
西北农林科技大学学报(自然科学版) 2015年10期
汪 雷,刘 瑶,丁一娟,王 雨,万华方,梅家琴,钱 伟
(西南大学 农学与生物科技学院,重庆市油菜工程技术研究中心,重庆 北碚400715)
油菜菌核病研究进展
汪 雷,刘 瑶,丁一娟,王 雨,万华方,梅家琴,钱 伟
(西南大学 农学与生物科技学院,重庆市油菜工程技术研究中心,重庆 北碚400715)
菌核病是危害油菜生产的主要病害之一。结合已有文献与本研究室的研究结果,文章就核盘菌的致病过程和致病机理、油菜菌核病抗病资源筛选、抗病育种现状、抗性遗传规律、抗病QTL定位及抗性基因发掘等方面进行了总结和展望。
油菜;菌核病;致病机理;分子机制;抗病育种
油菜是重要的油料作物,在食品、冶金、机械等多种行业应用广泛,经济价值高。核盘菌(Sclerotiniasclerotiorum)是一种广谱性病原真菌,可寄生于70多科400多种植物,由其引起的菌核病是油菜的一大主要病害[1]。在中国,菌核病可引起油菜减产10%~80%[2];在澳大利亚,菌核病导致油菜年均减产24%[3];在北美双低油菜主要种植区域[4]以及欧洲油菜种植区[5-7]该病的危害也日趋严重,加之全球气候变化,更可能加速该病害的流行[8]。化学杀菌剂的使用对该病害有一定防控效果,但效率却依赖于施用时间、植株发育阶段及环境等条件[9],且提高了生产成本[4],还会引发环境污染等问题。因此,选择和培育抗病品种是控制油菜菌核病最经济有效的途径[10-11]。本文参照国内外及本课题组的研究成果,综述了核盘菌的致病机理,并从抗源筛选、抗性遗传规律探索、抗病分子机制解析等方面总结了油菜菌核病目前的研究状况,以期为油菜菌核病的抗性改良提供参考。
1 核盘菌的致病过程及致病机理
1.1 致病过程
核盘菌经典的侵染循环路径主要是菌核在土壤、染病茎杆或种子中越冬、越夏,当外界环境适宜时,菌核可萌发产生菌丝或子囊孢子。子囊孢子成熟后从子囊中弹出,借气流传播,飘落到植株的叶片或花瓣上,在营养和环境条件适宜时萌发形成菌丝,侵入植物体,使植物受侵部位的细胞、组织坏死,形成水渍状病斑并腐烂,再漫延到其他健康植株,最后在患病部位尤其是茎杆中形成菌核。对油菜而言,其菌核病的发病符合这个过程(图1)。核盘菌在油菜花期来临前萌发,通过产生子囊孢子附着在油菜花瓣,继而以花瓣为营养基质生长萌发产生菌丝,菌丝再随着花瓣的凋零飘落到油菜的叶片或茎杆上,开始实现对油菜的侵染[12]。
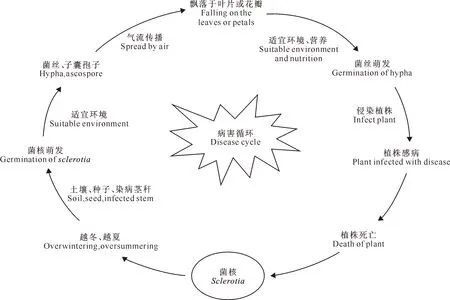
图1 油菜菌核病的病害循环
1.2 致病机理
目前,对核盘菌的致病机理尚缺乏系统全面的阐述,但研究表明细胞壁降解酶和草酸的分泌对核盘菌的致病有十分重要的作用[13-18]。核盘菌能分泌多种作用于寄主表皮的水解酶,如多聚半乳糖醛酸酶(Polygalacturonase,PG)、纤维素酶(Cellulase)、半纤维素酶(Hemicellulase)、果胶酶(Pectinase)、蛋白酶(Protease)、角质酶(Cutinase)、葡聚糖酶(Dextranase)等酶类,攻破寄主表层防护,降解细胞壁,侵染组织,为病菌提供营养,便于菌丝入侵和病菌繁殖[13-16,19]。核盘菌分泌的草酸是其关键致病因子之一[17]。研究发现,草酸通过以下方式在核盘菌的致病过程中发挥作用:①酸化、腐败寄主组织[20];②与细胞壁中胶层的Ca2+结合形成草酸钙结晶损坏细胞壁结构[21-22],与Mg2+螯合使叶绿素合成受阻和核糖体功能退化[23],与二价金属离子结合改变膜透性,导致电解质外渗,使细胞生理机能退化[24-25];③影响寄主氧化还原环境,促进寄主的氧化爆发,破坏寄主体内平衡,诱发细胞程序性死亡[26-31];④为病菌分泌水解酶营造适宜的pH环境[15,24-25],抑制与寄主抗病性相关的酶[15,32];⑤刺激保卫细胞对K+的吸收及对淀粉的降解,影响其细胞膜的渗透性,使保卫细胞功能失调,促使气孔张开,引发叶片发生脱水等症状,利于病菌入侵[33]。然而,有学者发现了分泌草酸毒素能力强,但致病力却极弱的核盘菌菌株[34-35],表明草酸并非核盘菌的惟一致病因子,核盘菌存在更复杂的致病机理,尚待进行更深入和系统的研究。
2 油菜菌核病抗性资源的筛选及抗病育种
2.1 抗病资源的筛选与鉴定
优良的抗源对作物抗病育种有至关重要的作用,因此学者们对甘蓝型油菜资源进行了系统而全面的菌核病抗性鉴定,发现甘蓝型油菜对菌核病的抗性存在差异,但并未找到免疫或者高抗材料[36-39],因此学者们尝试在甘蓝型油菜的近缘物种中发掘优良抗源。早在20世纪末,中国农业科学院油料作物研究所等对2 000余份芸薹属材料进行了鉴定,发现芥菜型油菜抗性最强,并选出约6%的高抗种质[37]; Li等[38]和Barbetti等[39]报道了在芥菜型油菜中存在较好的抗病资源,如澳大利亚的BrassicajunceaJM06018、BrassicajunceaJM06006和中国的Brassicajuncea2等材料。也有学者报道芸薹属中的埃芥对菌核病具有很强的抗性[40]。芸薹属中的甘蓝,尤其是野生甘蓝拥有很强的菌核病抗性[41-42];此外,还有学者在十字花科植物中寻找抗病资源,发现Erucastrumcardaminoides、Diplotaxistenuisiliqua和Erucastrumabyssinicum等野生型十字花科植物具有高抗菌核病的性能[43]。这些研究尽管均未找到完全免疫的材料,但明确了不同芸薹属物种对菌核病的抗性存在明显差异,甘蓝型油菜的近缘物种中存在抗性较强的资源,这对甘蓝型油菜的抗菌核病育种具有较大的应用价值。
2.2 抗菌核病育种
受限于油菜中抗病资源的缺乏,国内育种工作者通过传统育种方式选育出了一批具有部分抗性的甘蓝型油菜品种,如中双2号[44]、中油821[45]、中双4号[46],中双9号[47]、湘油11号和湘油15号[48]、中双11号[49]等;针对北美油菜种植,先锋育种公司也培育出了对菌核病抗性显著提高的杂交春油菜品种Pioneer Brand®45S51和Pioneer Brand®45S52[50]。虽然有学者曾采用无花瓣育种[51]、诱变育种[52]和基因工程育种[53-55]等方式来培育抗菌核病的油菜,但目前尚没有通过此类途径获得抗菌核病品种的报道。由此看来,要培育高抗菌核病的油菜必须依赖于优良抗源的发掘和先进育种技术的应用。
3 油菜菌核病的抗性遗传规律
由于没有找到高抗菌核病的油菜资源,人们只能采用具有部分抗性的油菜来研究菌核病的抗性遗传规律。早期研究发现,甘蓝型油菜BC1F1与BC2F1代具有明显偏亲本较强抗性的特性,其抗性表现为部分显性[56]。随后有学者发现,油菜菌核病抗性主要受核基因控制,由多个基因共同作用,抗性遗传由基因的加性效应和非加性效应共同控制,且加性遗传方差大于显性遗传方差[57-59]。何昆燕等[60]利用六世代分析模型研究了甘蓝型油菜的菌核病抗性遗传规律,发现抗性受2对主基因的控制,并受多基因修饰,基因效应及其互作随病情发展呈加性至显性的动态变化。近年来对油菜抗菌核病QTL定位的研究结果表明,油菜的菌核病抗性为数量性状,受多基因共同作用,各抗病位点间以加性效应为主,存在一定的上位性,且可能存在与环境的互作[61]。本研究小组在对甘蓝的抗病位点定位研究中也发现了同样的规律[62-63]。综上可知,各研究得出的结论不尽相同,这预示着油菜菌核病抗性遗传机制的复杂性,但可以确定的是,油菜菌核病抗性为数量性状,其抗性遗传受多基因控制,并倾向于认为加性效应在油菜菌核病的抗性遗传中起着主要作用。
除了缺乏完全免疫或高抗资源外,在分子水平上对油菜菌核病抗性的不完全阐述也是限制相关研究的一大因素。为此,学者们从DNA、RNA以及蛋白质水平上对油菜的抗菌核病基因进行了发掘,以期解析油菜抗菌核病的分子机制。
3.1 抗病QTL的定位
在DNA水平上,学者们主要采取QTL定位途径来寻找抗病相关基因(表1)。刘春林等[64]采用RAPD标记构建图谱与F2单株苗期叶片抗性结合的方式检测到3个抗病主效QTL,但未能提供该图谱与甘蓝型油菜各连锁群的对应关系,导致其所鉴定的抗病QTL难以与其他类似研究进行比较。何昆燕[65]和Yin等[66]各自利用3种不同的鉴定方法对同一个甘蓝型油菜DH群体进行多年、多环境下的抗病性鉴定,分别检测到18和21个抗病QTL,但可能由于鉴定方法的不完全相同,两者鉴定出的QTL重叠较少。Zhao等[2]在甘蓝型油菜F2∶3家系中鉴定出3个苗期和3个成株期抗病QTL,但苗期与成株期抗病QTL之间未检测到重叠,随后其又利用叶柄接种法在2个DH群体中分别获得11和2个抗病位点,单个QTL能解释的最高表型变异度为22%,并在N12和N02连锁群上的QTL区域检测到同源区段[67]。本研究室利用甘蓝型油菜DH群体在N12和N02上鉴定出抗病QTL,并发现2个QTL区域之间存在同源性[68]。近来,一些研究者通过不同的抗性鉴定方法在不同的群体中也鉴定出了一些抗病QTL,这些QTL多数分布在甘蓝型油菜的A亚基因组上,少数存在于C亚基因组,其中单个QTL能解释的最大变异度达到42%[67,69-72]。在最近的一篇报道中,研究者将重要的QTL位点锁定在了甘蓝型油菜的A9和C6染色体上,通过生物信息学手段并结合RT-PCR验证的方法,确定了一个吲哚硫代甲基转移酶(Indole glucosinolate methyltransferase,IGMT)编码基因(BnIGMT5.a),该基因为C6上QTL区间中的候选抗病基因[61]。

表1 甘蓝型油菜的抗菌核病QTLTable 1 Resistant QTLs against Sclerotinia sclerotiorum of Brassica napuse
注:R.抗病,S.感病;A.苗期离体叶片接种法,B.成株期田间牙签接种法,C.成株期田间菌丝琼脂块贴附法,D.活体叶柄接种法,E.花瓣接种法,F.草酸浸根法;括号中的编号代表该连锁群在Yin等[66]图谱中所对应的染色体情况;“-”表示数据缺失。
Note:R.Represents resistant,S.Represents susceptible;A.Represents detached leaf inoculation,B.Represents toothpick inoculation,C.Represents mycelial plug inoculation,D.Represents petiole inoculation,E.Represents petal inoculation,F.Represents oxalic-acid soak root inoculation;Numbers in parentheses represent the linkage group in the map of the corresponding chromosomal condition in Yin et al[66];“-” indicates missing data.
3.2 基因表达层面对抗病基因的挖掘
在RNA水平上,多数研究者采用cDNA芯片技术来研究油菜受核盘菌侵染后诱导表达的基因,或不同抗性材料在核盘菌侵染后基因的表达差异。研究发现,受核盘菌诱导的基因主要包括:①几丁质酶(Chitinase)、葡聚糖酶、渗调蛋白(Osmotins)和凝集素(Agglutinin)等病程相关蛋白(Pathogenesis-related proteins,PRs)编码基因;②细胞色素P450(Cytochrome P450,CYP450)相关蛋白基因以及谷胱甘肽转移酶(Glutathione transferase)基因;③茉莉酸(Jasmonic acid,JA)生物合成和信号传递、活性氧代谢(Reactive oxygen metabolism)及氧爆反应(Oxidative burst)相关蛋白基因;④细胞壁结构与功能相关基因;⑤蛋白激酶(Protein kinases)、分子转运蛋白和细胞发育相关基因等;⑥转录因子(锌指、WRKY、AP2和MYB家族)、次生代谢物合成基因等[73-75]。其中,初步确定的候选抗病相关基因包括:Bn-29、Bn-30和Bn-10(三者一起正调控油菜菌核病的抗性)[76],促分裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK)基因[77-78]、BnMAPK7-1、BnMAPK7-2和BnMAPK7-3[79],正调控抗性的乙烯信号转导途径(Ethylene-signaling pathway)关键调节因子BnEIN3[80],乙烯应答转录因子(Ethylene-responsive transcriptional factors,ERFs)BnERF104和BnERF50基因[81-82]及多聚半乳糖醛酸酶抑制蛋白(Polygalacturonase-inhibiting protein,PGIP)基因Pgip1和Pgip2[83-84]等。尽管众多学者都致力于揭示油菜抗病分子机制,但受cDNA芯片技术本身的限制,这些研究无法在整个转录组水平上对油菜接种核盘菌后基因的表达变化进行系统而全面地跟踪,因而这些研究类似于一幅拼图中的少数几个小单元,只有用更多的单元来补充和完善整幅图片后,才能明确油菜抗核盘菌的分子机制。
3.3 蛋白质层面对抗病基因的挖掘
在蛋白质水平上,早期学者们主要是通过检测少量与植物抗逆/抗病相关蛋白在接种核盘菌前后的变化情况来判断其是否与菌核病抗性相关,如王汉中等[85]研究了苯丙氨酸解氨酶(Phenylalanine ammonia-lyase,PAL)、几丁质酶、β-1,3-葡聚糖酶(β-1,3-dextranase)、过氧化物酶(Peroxidase,POD)和多酚氧化酶(Polyphenol oxidase,PPO)等油菜中重要的防御酶类在抗、感材料接种前后的活性变化,发现PAL、几丁质酶和β-1,3-葡聚糖酶、POD、PPO与油菜对菌核病的抗性密切相关;张志元等[86]发现,超氧物歧化酶(Superoxide dismutase,SOD)与油菜对菌核病的抗性有关。后来学者们开始采用蛋白质组学来研究油菜抗菌核病的机制和抗病基因。皇甫海燕等[84]对油菜抗菌核病近等基因系及其感病亲本在核盘菌侵染后的蛋白质组进行了分析,发现组成线粒体ATP合酶F1的重要亚基F1β、糖酵解关键酶磷酸甘油酸激酶(Phosphoglycerate kinase,PGK)、参与合成维生素B1的辅酶噻唑(thiazole)生物合成酶THI1和1,5-二磷酸核酮糖羧化酶/加氧酶(Ribulose-1,5-bisphosphate carboxylase/oxygenas,RuBisCO)在油菜对菌核病的抗性反应中差异表达。Wen等[87]对甘蓝型油菜抗、感品种接种核盘菌前后进行了比较蛋白组学分析,发现与抗性相关的蛋白主要在病原抗性、抗氧化作用和转录调节这几个部分发生作用,并发现一些作用于活性氧清除的胰蛋白酶抑制蛋白(Trypsin inhibitor protein,TIP)、富含甘氨酸的细胞壁蛋白(Glycine rich protein,GRP)、热激蛋白(Heat shock protein,HSP)和硫醇甲基转移酶(Thiol methyltransferase,TMT)在抗性品种中呈上调或特异表达。
4 油菜菌核病的研究展望
尽管科学家们对油菜抗菌核病育种及相关研究投入了较大精力,也在取得了一定成效,但还存在一些重要问题需要解决:一是油菜中缺乏可利用的优良抗源,导致抗性位点定位的重复性差,抗病基因鉴定工作进展缓慢,其研究结果难以应用于育种实践,油菜菌核病抗性的改良难以获得突破性进展;二是寄主与核盘菌的互作机制复杂,病原菌的致病机理、寄主的抗病机制至今仍不清晰,从而限制了抗病基因的鉴定和油菜抗菌核病的分子设计育种;三是研究手段尚需改进,当前对抗病基因的鉴定研究多采用QTL定位或通过检验少数基因对核盘菌的响应情况来推测其与抗性的关系,难以准确鉴定出抗病基因,更无法了解寄主的抗病机制。
鉴于上述问题,笔者认为油菜菌核病研究的首要内容仍然是广泛搜寻可利用的优良抗病资源,包括油菜的近缘物种乃至十字花科其他植物。在获得优良抗源的基础上,通过远缘杂交或传统育种方式将其抗病成分转入油菜,利用其进行抗病位点鉴定的准确性必将得到有效提高,抗病机制的分析难度也将降低。随着生物学和生物信息学研究技术的快速发展,以及甘蓝型油菜、甘蓝和白菜全基因组测序的进行和完成,结合全基因组水平、转录组水平和代谢组水平的分析手段,可快速、全面、准确地解析油菜的抗菌核病机制并发掘抗病基因,这对于油菜抗菌核病的分子设计育种具有重要的实践意义。
[1] Bolton M D,Thomma B P,Nelson B D.Sclerotiniasclerotiorum(Lib.) de Bary:Biology and molecular traits of a cosmopolitan pathogen [J].Molecular Plant Pathology,2006,7(1):1-16.
[2] Zhao J,Meng J.Genetic analysis of loci associated with partial resistance toSclerotiniasclerotiorumin rapeseed (BrassicanapusL.) [J].Theoretical and Applied Genetics,2003,106(4):759-764.
[3] Garg H,Sivasithamparam K,Banga S S,et al.Cotyledon assay as a rapid and reliable method of screening for resistance againstSclerotiniasclerotioruminBrassicanapusgenotypes [J].Australasian Plant Pathology,2008,37(2):106-111.
[4] Del Río L,Bradley C,Henson R,et al.Impact ofSclerotiniastem rot on yield of canola [J].Plant Disease,2007,91(2):191-194.
[5] Koch S,Dunker S,Kleinhenz B,et al.A crop loss-related forecasting model forSclerotiniastem rot in winter oilseed rape [J].Phytopathology,2007,97(9):1186-1194.
[8] Siebold M,Tiedemann A V.Potential effects of global warming on oilseed rape pathogens in Northern Germany [J].Fungal Ecology,2012,5(1):62-72.
[9] Turkington T,Morrall R.Use of petal infestation to forecastSclerotiniastem rot of canola:The influence of inoculum variation over the flowering period and canopy density [J].Phytopathology,1993,83(6):682-689.
[10] Zhao J,Peltier A,Meng J,et al.Evaluation ofSclerotiniastem rot resistance in oilseedBrassicanapususing a petiole inoculation technique under greenhouse conditions [J].Plant Disease,2004,88(9):1033-1039.
[11] Li C,Li H,Sivasithamparam K,et al.Expression of field resistance under Western Australian conditions toSclerotiniasclerotiorumin Chinese and AustralianBrassicanapusandBrassicajunceagermplasm and its relation with stem diameter [J].Crop and Pasture Science,2006,57(10):1131-1135.
[12] 张信扬,彭连英.油菜菌核病发生规律研究 [J].云南农业大学学报,2000,15(3):289-290.
Zhang X Y,Peng L Y.Studies on the occurrence of sclerotium blight of rape oil [J].Journal of Yunnan Agricultural University,2000,15(3):289-290.(in Chinese)
[13] Chahed H,Ezzine A,Mlouka A B,et al.Biochemical caracterization,molecular cloning,and structural modeling of an interesting β-1,4-glucanase fromSclerotiniasclerotiorum[J].Molecular Biotechnology,2014,55(4):340-350.
[14] Bashi Z D,Rimmer S R,Khachatourians G G,et al.Factors governing the regulation ofSclerotiniasclerotiorumcutinase A and polygalacturonase 1 during different stages of infection [J].Canadian Journal of Microbiology,2012,58(5):605-616.
[15] 毛 玮,侯英敏,刘志文.核盘菌和草酸诱导下的油菜几种酶活力的变化分析 [J].大连工业大学学报,2011,30(1):39-42.
Mao W,Hou Y M,Liu Z W.Analysis of several enzymes activity induced bySclerotiniasclerotiorumand oxalic acid in rapeseed [J].Journal of Dalian Polytechnic University,2011,30(1):39-42.(in Chinese)
[16] 刘小燕,高智谋,汪世军,等.油菜菌核病菌致病力与纤维素酶活性关系的初步研究 [J].现代农业科学,2009,16(6):1-3.
Liu X Y,Gao Z M,Wang S J,et al.Preliminary study on the relationship between pathogenicity and cellulase activity ofSclerotiniasclerotiorumisolates [J].Modem Agricultural Sciences,2009,16(6):1-3.(in Chinese)
[17] Zhou T,Boland G J.Mycelial growth and production of oxalic acid by virulent and hypovirulent isolates ofSclerotiniasclerotiorum[J].Canadian Journal of Plant Pathology,1999,21(1):93-99.
[18] Godoy G,Steadman J,Dickman M,et al.Use of mutants to demonstrate the role of oxalic acid in pathogenicity ofSclerotiniasclerotiorumonPhaseolusvulgaris[J].Physiological and Molecular Plant Pathology,1990,37(3):179-191.
[19] Riou C,Freyssinet G,Fevre M.Production of cell wall-degrading enzymes by the phytopathogenic fungusSclerotiniasclerotiorum[J].Applied and Environmental Microbiology,1991,57(5):1478-1484.
[20] Punja Z K,Huang J S,Jenkins S F.Relationship of mycelial growth and production of oxalic acid and cell wall degrading enzymes to virulence inSclerotiumrolfsii[J].Canadian Journal of Plant Pathology,1985,7(2):109-117.
[21] Durman S B,Menendez A B,Godeas A M.Variation in oxalic acid production and mycelial compatibility within field populations ofSclerotiniasclerotiorum[J].Soil Biology and Biochemistry,2005,37(12):2180-2184.
[22] Heller A,Witt G T.Oxalic acid has an additional,detoxifying function inSclerotiniasclerotiorumpathogenesis [J].PLOS One,2013,8(8):e72292.
[23] Dutton M V,Evans C S.Oxalate production by fungi:Its role in pathogenicity and ecology in the soil environment [J].Canadian Journal of Microbiology,1996,42(9):881-895.
[25] 李玉芳,官春云.油菜菌核病菌侵染的组织病理学、致病及抗病机制的研究 [J].作物研究,2005(5):327-331.
Li Y F,Guan C Y.The research of the histopathology, pathogenicity and disease mechanism of the infection aboutSclerotiniasclerotiorum[J].Crop Research,2005(5):327-331.(in Chinese)
[26] Cessna S G,Sears V E,Dickman M B,et al.Oxalic acid,a pathogenicity factor forSclerotiniasclerotiorum,suppresses the oxidative burst of the host plant [J].Plant Cell,2000,12(11):2191-2199.
[27] Maeno E,Ishizaki Y,Kanaseki T,et al.Normotonic cell shrin-kage because of disordered volume regulation is an early prerequisite to apoptosis [J].Proceedings of the National Academy of Sciences,2000,97(17):9487-9492.
[28] Guo X,Stotz H U.Defense againstSclerotiniasclerotioruminArabidopsisis dependent on jasmonic acid,salicylic acid,and ethylene signaling [J].Molecular Plant-Microbe Interactions,2007,20(11):1384-1395.
[29] Errakhi R,Meimoun P,Lehner A,et al.Anion channel activity is necessary to induce ethylene synthesis and programmed cell death in response to oxalic acid [J].Journal of Experimental Botany,2008,59(11):3121-3129.
[30] Kim K S,Min J-Y,Dickman M B.Oxalic acid is an elicitor of plant programmed cell death duringSclerotiniasclerotiorumdisease development [J].Molecular Plant-Microbe Interactions,2008,21(5):605-612.
[31] Williams B,Kabbage M,Kim H J,et al.Tipping the balance:Sclerotiniasclerotiorumsecreted oxalic acid suppresses host defenses by manipulating the host redox environment [J].PLoS Pathogens,2011,7(6):e1002107.
[32] 刘胜毅,潘家荣.油菜对毒素草酸的吸收代谢与抗性机理 [J].植物病理学报,1998,28(1):33-37.
Liu S Y,Pan J R.Oilseed rape’s absorption metabolism and resistance mechanism for oxalate [J].Acta Phytopathologica Sinica,1998,28(1):33-37.(in Chinese)
[33] Guimarães R L,Stotz H U.Oxalate production bySclerotiniasclerotiorumderegulates guard cells during infection [J].Plant Physiology,2004,136(3):3703-3711.
[34] 李国庆.作物菌核病病源:核盘菌的多样性研究 [D].武汉:华中农业大学,1996.
Li G Q.Polymorphisms ofSclerotiniasclerotioruma kind economically-important pathogen [D].Wuhan: Huazhong Agricultural University,1996.(in Chinese)
[35] 李国庆,王道本,姜道宏,等.不同核盘菌及其近缘物种的RAPD分析 [J].植物病理学报,2000,30(2):166-170.
Li G Q,Wang D B,Jiang D H,et al.RAPD assay of strains ofSclerotiniasclerotiorumand its related species [J].Acta Phytopathologica Sinica,2000,30(2):166-170.(in Chinese)
[36] Mei J,Qian L,Disi J,et al.Identification of resistant sources againstSclerotiniasclerotioruminBrassicaspecies with emphasis onB.oleracea[J].Euphytica,2011,177(3):393-399.
[37] 周乐聪,刘胜毅,周必文.油菜品种资源对菌核病的抗性鉴定 [J].中国油料作物学报,1994(4):69-71.
Zhou Y C,Liu S Y,Zhou B W.Rapeseed germplasm for resistance toSclerotiniaidentification [J].Chinese Journal of Oil Crop Sciences,1994(4):69-71.(in Chinese)
[38] Li C X,Liu S Y,Sivasithamparam K,et al.New sources of resistance toSclerotiniastem rot caused bySclerotiniasclerotiorumin Chinese and AustralianBrassicanapusandB.junceagermplasm screened under Western Australian conditions [J].Australasian Plant Pathology,2009,38(2):149-152.

[40] 张洁夫,张玉卿,伍贻美.油菜种质资源抗 (耐) 菌核病性筛选与鉴定 [J].江苏农业科学,1994(2):26-28.
Zhang J F,Zhang Y Q,Wu Y M.Rapeseed germplasm screening and identification of resistance toSclerotinia[J].Jiangsu Agricultural Sciences,1994(2):26-28.(in Chinese)
[41] 梅家琴.甘蓝及其野生种种质资源评价 [D].重庆:西南大学,2008.
Mei J Q.Evaluation of germplasm inBrassicaoleraceaand the wild species [D].Chongqing:Southwest University,2008.(in Chinese)
[42] 陈碧云,伍晓明,陆光远,等.欧洲野生甘蓝资源的菌核病抗性鉴定与评价 [C]//中国作物学会油料作物专业委员会.中国作物学会油料作物专业委员会第六次代表大会暨学术年会论文集.武汉:科学出版社,2008:393-396.
Chen B Y,Wu X M,Lu G Y,et al.The identification and evaluation of resistance toSclerotiniaof the european wild cabbage resources [C]//Crop Science Society of Professional Committee of China Oil Crops.The sixth congress and annual conference of crop science society of professional committee of China oil crops proceedings.Wuhan:Science Press,2008:393-396.(in Chinese)
[43] Garg H,Atri C,Sandhu P S,et al.High level of resistance toSclerotiniasclerotiorumin introgression lines derived from hybridization between wild crucifers and the cropBrassicaspeciesB.napusandB.juncea[J].Field Crops Research,2010,117(1):51-58.
[44] 刘澄清,黄有菊,邹崇顺.中双二号双低高产多抗 (耐) 甘蓝型油菜新品种选育 [J].中国油料作物学报,1990(4):59-63.
Liu C Q,Huang Y J,Zou C S.New breeding of the double low yielding and the multi-resistant varieties about Zhongshuang 2 ofBrassicanapus[J].Chinese Journal of Oil Crop Sciences,1990(4):59-63.(in Chinese)
[45] 王宗礼.中油 821 [J].中国农村科技,1997(1):24.
Wang Z L.Zhongyou 821 [J].China Rural Science & Technology,1997(1):24.(in Chinese)
[46] 钟敏秀.全国油菜双低之冠:中双4号 [J].中国农技推广,1998(1):42-43.
Zhong M X.National championship of double low rapeseed-Zhongshuang 4 [J].China Agricultural Technology Extension,1998(1):42-43.(in Chinese)
[47] Wang H,Liu G,Zheng Y,et al.Breeding of theBrassicanapuscultivar Zhongshuang 9 with high-resistance toSclerotiniasclerotiorumand dynamics of its important defense enzyme activity [J].Scientia Agricultura Sinica,2004,37(1):23-28.
[48] 官春云,李方球,李 构,等.芸芥(ErucasativaMill.)对菌核病的抗性研究 [J].中国农业科学,2004,37(8):1138-1143.
Guan C Y,Li F Q,Li G,et al.Resistance of rocket salad (ErucasativaMill.) to stem rot (Sclerotiniasclerotiorum) [J].Scientia Agricultura Sinica,2004,37(8):1138-1143.(in Chinese)
[49] 屠建明.超高含油量油菜新品种中双11号特征特性及栽培技术 [J].中国农技推广,2011,27(10):19-20.
Tu J M.Characteristics and cultivation techniques of the new ultrahigh oil content of rapeseed variety Zhongshuang 11 [J].China Agricultural Technology Extension,2011,27(10):19-20.(in Chinese)
[51] 陈玉卿,张洁夫,吕忠进.甘蓝型无花瓣油菜对菌核病抗(耐)性研究 [J].江苏农业科学,1991(6):10-13.
Chen Y Q,Zhang J F,Lü Z J.The research of theSclerotiniaresistantenceforBrassicanapuswithout petals [J].Jiangsu Agricultural Sciences,1991(6):10-13.(in Chinese)
[52] Liu S,Wang H,Zhang J,et al.Invitromutation and selection of doubled-haploidBrassicanapuslines with improved resistance toSclerotiniasclerotiorum[J].Plant Cell Reports,2005,24(3):133-144.
[53] Dong X,Ji R,Guo X,et al.Expressing a gene encoding wheat oxalate oxidase enhances resistance toSclerotiniasclerotiorumin oilseed rape (Brassicanapus) [J].Planta,2008,228(2):331-340.
[54] Liu H,Guo X,Naeem M S,et al.TransgenicBrassicanapusL.lines carrying a two gene construct demonstrate enhanced resistance againstPlutellaxylostellaandSclerotiniasclerotiorum[J].Plant Cell Tissue and Organ Culture,2011,106(1):143-151.
[55] Fu X,Wen M,Wang F,et al.Overexpression of an nsLTPs-like antimicrobial protein gene (LJAMP2) from motherwort (Leonurusjaponicus) enhances resistance toSclerotiniasclerotiorumin oilseed rape (Brassicanapus) [J].Physiological and Molecular Plant Pathology,2013,82:81-87.
[56] 刘澄清,杜德志,黄有菊,等.甘蓝型油菜的抗耐病性及其遗传效应研究 [J].中国农业科学,1991,24(3):43-49.
Liu C Q,Du D Z,Huang Y J,et al.Study of disease resistance and genetic effects ofBrassicanapus[J].Scientia Agricultura Sinica,1991,24(3):43-49.(in Chinese)
[57] 黄永菊,陈 军.甘蓝型油菜菌核病抗 (耐) 性的遗传研究:Ⅰ.抗性遗传属性与配合力分析 [J].中国油料作物学报,2000,22(4):1-5.
Huang Y J,Chen J.Genetic studies on resistance (tolerence) toSclerotiniasclerotiorumin rapeseed (B.napusL.):Ⅰ.Genetic nature and analysis of combining abilities of resistance [J].Chinese Journal of Oil Crop Sciences,2000,22(4):1-5.(in Chinese)
[58] 杨 伟.甘蓝型油菜品种多态性及抗菌核病分子标记 [D].北京:中国农业科学院,2001.
Yang W.Genotypes polymorphism and identification of RAPD markers linked toSclerotinasclerotiorumresistance inBrassicanapus[D].Beijing:Chinese Academy of Agricultural Sciences,2001.(in Chinese)
[59] 刘胜毅,张建坤,许泽永,等.甘蓝型油菜对核盘菌及其毒素的抗性遗传分析 [J].植物保护学报,2005,32(1):43-47.
Liu S Y,Zhang J K,Xu Z Y,et al.Genetic analysis of resistance toSclerotinasclerotiorumand its toxin oxalic acid inBrassicanapus[J].Acta Phytophylacica Sinica,2005,32(1):43-47.(in Chinese)
[60] 何昆燕,易 斌,傅廷栋,等.甘蓝型油菜菌核病抗性的遗传分析 [J].作物学报,2005,31(11):1495-1499.
He K Y,Yi B,Fu T D,et al.Genetic analysis of resistance toSclerotinasclerotioruminBrassicanapusL. [J].Acta Agronomica Sinica,2005,31(11):1495-1499.(in Chinese)
[61] Wu J,Cai G,Tu J,et al.Identification of QTLs for resistance toSclerotiniastem rot and BnaC.IGMT5.a as a candidate gene of the major resistant QTL SRC6 inBrassicanapus[J].PLoS One,2013,8(7):e67740.
[62] Mei J,Ding Y,Lu K,et al.Identification of genomic regions involved in resistance againstSclerotiniasclerotiorumfrom wildBrassicaoleracea[J].Theoretical and Applied Genetics,2013,126(2):549-556.
[63] Disi J O,Mei J,Wei D,et al.Inheritance of leaf and stem resistance toSclerotiniasclerotiorumin a cross betweenBrassicaoleraceavar.alboglabra[J].The Journal of Agricultural Science,2014,152(1):146-152.
[64] 刘春林,官春云,李 栒,等.油菜分子标记图谱构建及抗菌核病性状的QTL定位 [J].遗传学报,2000,27(10):918-924.
Liu C L,Guan C Y,Li X,et al.Construction of linkage map and mapping resistance gene ofSclerotinasclerotioruminBrassicanapus[J].Journal of Genetics and Genomics,2000,27(10):918-924.(in Chinese)
[65] 何昆燕.利用DH(Doubled Haploid)群体对甘蓝型油菜菌核病抗性基因的QTL定位和分析 [D].武汉:华中农业大学,2003.
He K Y.Mapping and analysis of QTLs associated with disease resistance toSclerotinasclerotioruminBrassicanapusin using doubled haploid [D].Wuhan:Huazhong Agricultural University,2003.(in Chinese)
[66] Yin X,Yi B,Chen W,et al.Mapping of QTLs detected in aBrassicanapusDH population for resistance toSclerotiniasclerotiorumin multiple environments [J].Euphytica,2010,173(1):25-35.
[67] Zhao J,Udall J A,Quijada P A,et al.Quantitative trait loci for resistance toSclerotiniasclerotiorumand its association with a homeologous non-reciprocal transposition inBrassicanapusL. [J].Theoretical and Applied Genetics,2006,112(3):509-516.
[68] Wei D,Mei J,Fu Y,et al.Quantitative trait loci analyses for resistance toSclerotiniasclerotiorumand flowering time inBrassicanapus[J].Molecular Breeding,2014,34(4):1797-1804.


[72] 涂江颖.甘蓝型油菜抗菌核病的遗传分析及其基因定位 [D].武汉:华中农业大学,2011.
Tu J Y.Genetic analysis and QTL mapping of resistanee toSclerotiniaselerotiorumin rapeseed (BrassicanapusL.) [D].Wuhan: Huazhong Agricultural University,2011.(in Chinese)
[73] 韦淑亚.甘蓝型油菜菌核病抗性相关基因 eDNA 文库构建及评价 [D].武汉:华中农业大学,2004.
Wei S Y.Construction and evaluation of eDNA libray related to the defense genes induced bySclerotinasclerotioruminBrassicanapus[D].Wuhan:Huazhong Agricultural University,2004.(in Chinese)
[74] Zhao J,Wang J,An L,et al.Analysis of gene expression profiles in response toSclerotiniasclerotioruminBrassicanapus[J].Planta,2007,227(1):13-24.
[75] Zhao J,Buchwaldt L,Rimmer S R,et al.Patterns of differential gene expression inBrassicanapuscultivars infected withSclerotiniasclerotiorum[J].Molecular Plant Pathology,2009,10(5):635-649.
[76] 任秋红.油菜菌核病抗性相关基因的功能研究 [D].郑州:郑州大学,2010.
Ren Q H.Funetion analysis of resistance-related genesagainstSclerotinasclerotioruminBrassicanapus[D].Zhengzhou:Zhengzhou University,2010.(in Chinese)
[77] Wang Z,Mao H,Dong C,et al.Overexpression ofBrassicanapusMPK4 enhances resistance toSclerotiniasclerotiorumin oilseed rape [J].Molecular Plant-Microbe Interactions,2009,22(3):235-244.
[78] 王 政.甘蓝型油菜 MPK4 抗菌核病功能研究 [D].北京:中国农业科学院,2008.
Wang Z.Function ofBrassicanapusMPK4 in resistence toSclerotinasclerotiorum[D].Beijing:Chinese Academy of Agricultural Sciences,2008.(in Chinese)
[79] 朱 斌,陆俊杏,彭 茜,等.甘蓝型油菜MAPK7基因家族及其启动子的克隆与表达分析 [J].作物学报,2013,39(5):789-805.
Zhu B,Lu J X,Peng Q,et al.Cloning and analysis ofMAPK7 gene family and their promoters fromBrassicanapus[J].Acta Agronomica Sinica,2013,39(5):789-805.(in Chinese)
[80] 许李明.油菜菌核病抗性相关基因EIN3及3A06和6C02的功能研究 [D].武汉:中南民族大学,2009.
Xu L M.Function analysis of resistance-related genesEIN3 and 3A06 and 6C02 againstSclerotiniasclerotioruminBrassicanapus[D].Wuhan:South-Central University for Nationalitie,2009.(in Chinese)
[81] 汪承刚,蔡 丽,董彩华,等.甘蓝型油菜BnERF104 超表达增强了转基因拟南芥对核盘菌的抗性 [J].中国油料作物学报,2011,33(4):325-330.
Wang C G,Cai L,Dong C H,et al.Over-expression ofBnERF104 fromBrassicanapusenhanced transgenicArabidopsisresistance toSclerotiniasclerotiorum[J].Chinese Journal of Oil Crop Sciences,2011,33(4):325-330.(in Chinese)
[82] 汪承刚,李 丹,许李明,等.甘蓝型油菜BnERF50的超表达增强了转基因拟南芥对核盘菌的抗性 [J].中国油料作物学报,2011,33(5):427-432.
Wang C G,Li D,Xu L M.Over-expression ofBnERF50 fromBrassicanapusenhanced transgenicArabidopsisresistance toSclerotiniasclerotiorum[J].Chinese Journal of Oil Crop Sciences,2011,33(5):427-432.(in Chinese)
[83] Hegedus D D,Li R,Buchwaldt L,et al.Brassicanapuspossesses an expanded set of polygalacturonase inhibitor protein genes that are differentially regulated in response toSclerotiniasclerotioruminfection,wounding and defense hormone treatment [J].Planta,2008,228(2):241-253.
[84] 皇甫海燕,官春云.甘蓝型油菜抗菌核病近等基因系和感病亲本蛋白差异的初步研究 [J].中国农业科学,2010,43(10):2000-2007.
Huangfu H Y,Guan C Y.A preliminary analysis of differential proteins betweenBrassicanapusresistance toSclerotiniasclerotiorumnear-isogenic lines and susceptible parent [J].Scientia Agricultura Sinica,2010,43(10):2000-2007.(in Chinese)
[85] 王汉中,刘贵华,郑元本,等.抗菌核病双低油菜新品种中双9号选育及其重要防御酶活性变化规律的研究 [J].中国农业科学,2004,37(1):23-28.
Wang H Z,Liu G H,Zheng Y B,et al.Breeding of theBrassicanapuscultivar Zhongshuang 9 with high-resistance toSclerotiniasclerotiorumand dynamics of its important defence enzyme activity [J].Scientia Agricultura Sinica,2004,37(1):23-28.(in Chinese)
[86] 张志元,张 翼,罗永兰,等.油菜抗菌核病的部分生理生化机制 [J].中国油料作物学报,2007,29(2):189-194.
Zhang Z Y,Zhang Y,Luo Y L,et al.Partial biochemical and physiological mechanisms ofBrassicanapusmediating resistance toSclerotiniasclerotiorum[J].Chinese Journal of Oil Crop Sciences,2007,29(2):189-194.(in Chinese)
[87] Wen L,Tan T,Shu J,et al.Using proteomic analysis to find the proteins involved in resistance againstSclerotiniasclerotiorumin adultBrassicanapus[J].European Journal of Plant Pathology,2013,137(3):505-523.
Advance inSclerotiniastem rot of rapeseed
WANG Lei,LIU Yao,DING Yi-juan,WANG Yu,WAN Hua-fang,MEI Jia-qin,QIAN Wei
(SchoolofAgronomyandBiotechnology,SouthwestUniversity,ChongqingRapeseedEngineering&TechnologyResearchCenter,Beibei,Chongqing400715,China)
Sclerotiniastem rot caused by fungal pathogenSclerotiniasclerotiorumis one of the most devastating diseases in rapeseed production.Combining with the existing research literature and the findings of our lab,the review summarized and prospected the researches in infection process and pathogenesis mechanism ofS.sclerotiorum,disease-resistant resources,breeding progress,inheritance of resistance,resistant QTL loci,and resistance genes in rapeseed.
rapeseed;Sclerotiniastem rot; pathogenesis;molecular mechanism;resistance breeding
时间:2015-09-09 15:41
10.13207/j.cnki.jnwafu.2015.10.012
2014-03-05
中央高校基本业务费项目(SWU113106);国家自然科学基金项目(31171585)
汪 雷(1991-),女,重庆铜梁人 ,硕士,主要从事作物遗传育种研究。E-mail:wangleicqtl@163.com
梅家琴(1982-),女,四川彭州人,副教授,主要从事植物分子育种及芸薹属物种菌核病抗性研究。 E-mail:jiaqinmay@163.com
S332.2
A
1671-9387(2015)10-0085-09
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20150909.1541.024.html
猜你喜欢
今日农业(2022年4期)2022-06-01
四川蚕业(2021年3期)2021-02-12
西藏农业科技(2019年3期)2019-11-04
西藏农业科技(2019年1期)2019-07-25
西藏农业科技(2018年4期)2018-04-25
浙江农业学报(2017年1期)2017-05-17
上海农业学报(2017年3期)2017-04-10
蚕桑通报(2015年1期)2015-12-23
作物研究(2014年6期)2014-03-01
植物营养与肥料学报(2012年3期)2012-10-26
