
天目山针阔混交林林木碳储量树种分布特征
2015-01-03 05:12范佩佩韦新良郭如意
西北林学院学报 2015年5期
范佩佩,韦新良,郭如意
(1.浙江农林大学 环境与资源学院,浙江 临安311300;2.浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安311300)
森林碳储量是反映森林资源及其生态环境效能的重要指标,越来越受到各国政府部门的重视,许多学者也对此进行了大量的研究[1]。准确估计森林生态系统的碳储量,评价森林的碳汇功能意义重大[2-3]。碳储量的测定如生物量模型的建立,碳密度的估算等方面有较多的研究,为探讨针阔混交林碳储量的分布提供了相应的方法。碳储量分布的规律研究为我们了解森林生态系统的碳平衡,改善和维护区域生态环境的功能提供科学的理论基础和实践依据[4-7]。我国对森林碳储量的研究起步较晚,也是以森林生态系统的生物量测定为开端。对森林植被生物量进行估算是当前生态学研究的重要内容,也是研究陆地生态系统碳动态及碳循环的基础[8-9]。胸径与树高是反映林木状况的最基本因子,有关胸径与树高的生长模型很多[10-12],可以反映林分的平均状态,也是检验森林经营效果、对林分提出合理经营措施的主要依据[13]。树冠是树木进行光合作用的重要场所,它决定树木的生活力和生产力,冠幅是可视化的重要参数[14]。
本研究的目的在于对天目山针阔混交林林分的碳储量树种分布特性进行研究,以便掌握针阔混交林林分树种碳储量随直径、树高和冠幅分布的形态和构成比例,为检验森林经营效果、制定合理的经营措施提供依据。
1 研究区概况
天目山位于浙江省西北部,其主体分别由东天目和西天目组成。地理坐标为191°24′11″-119°28′21″E、30°18′30″-30°24′55″N。区域内年平均气温为14.8~8.8℃,年均降水量可达1 390~1 870 mm,形成浙江省西北部的多雨中心[15]。天目山地处中亚热带气候带北缘,具有中亚热带向北亚热带气候、丘陵向平原气候过渡的特征。区域内气候温和、地质古老、生物资源极其丰富。这些独特的环境条件构成了天目山植物区系的古老性,复杂性和种类丰富性,共计有苔类植物70种、藓类植物240种、蕨类植物110种、种子植物1 570种。其中国家重点保护植物25种,以天目山命名的植物24种,珍稀孑遗植物银杏的野生种也分布于此,具有重要的科研价值[16]。
2 材料与方法
2.1 材料
2011年,在天目山区选择没有人为干扰且具有典型性和代表性、树种类型多样的天然针阔混交林林地,设置15个30m×30m的样地,对木本植物个体进行种类识别、挂牌等。在样地设置时,踏查了全林分,了解林分界线和特点。采用全面调查法,调查样地内所有胸径≥5cm的林木特征值,包括树种、胸径、树高、枝下高、冠幅等,同时用全站仪测定并记录每棵林木的具体坐标。样地中主要树种有:马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)、枫香(Liquidambar formosana)、短柄枹(Quercus glandulifera var.brevipetiolata)、白栎(Quercus fabri)、苦槠(Castanopsis sclerophylla)、山 矾 (Symplocos caudata)、锥 栗 (Castanea henryi)、麻栎(Quercus acutissima)、化香(Platycarya strobilacea)、木 荷 (Schima superba)、青 冈(Cyclobalanopsis glauca)、檵 木 (Loropetalum chinense)、山合欢(Albizia kalkora)等。
2.2 单株林木碳储量测算
2.2.1 林木生物量测算 测算林木碳储量的方法有许多种,以生物量法较为常见。依据样地调查所得数据,选取适合浙江省各类树种的单株生物量模型计算出各类林木的单株生物量。鉴于样地内树种繁多,根据资料将样地内所有树种分为松类、杉木、Ⅰ类硬阔、Ⅱ类硬阔、软阔类等五大类树种进行生物量估算。其中,松类主要包括马尾松、金钱松;杉木类主要是杉木;Ⅰ类硬阔的树干木材密度一般<0.7 g/cm3,主要包括香樟、木荷、华东楠等树种;Ⅱ类硬阔的树干木材密度一般>0.7g/cm3,主要包括冬青、栎类、苦槠等树种;软阔类树种主要包括枫香、枫杨、檫木等树种。计算单株林木生物量模型公式为[17-18]:

式中:W 为单株林木生物量,W1为树干部分生物量,W2为树冠部分生物量,W3为树根部分生物量(表1)。
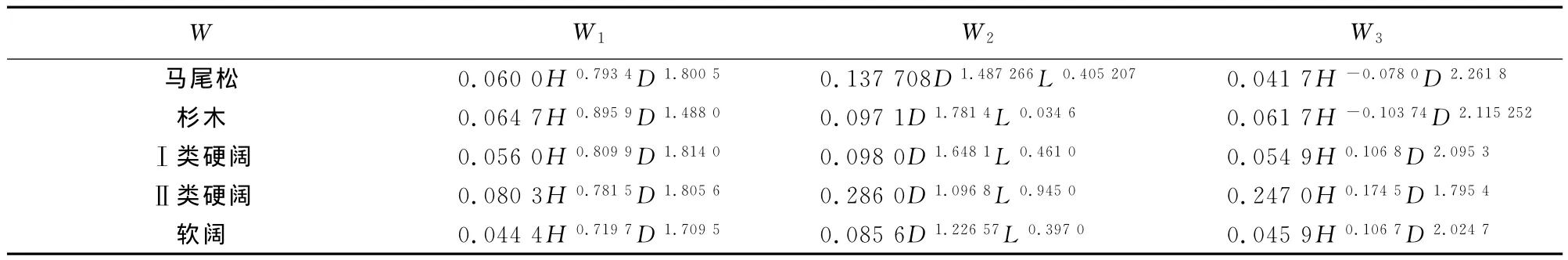
表1 树种单株生物量模型Table 1 Single plant biomass model of tree species
2.2.2 林木碳储量测算 计算出各类树种单株生物量后,根据公式计算出单株林木的碳储量。具体公式如下:某一森林类型(树种)碳储量=某一森林类型(树种)生物量×森林类型(树种)含碳系数
本文所采用的含碳系数[19]见表2。
2.3 分树种林木碳储量统计
计算出各树种单株林木的碳储量之后,将各样地所有林木按不同树种进行碳储量求和,并且分树种求出碳储量的平均值。即可得出各树种的总碳储 量及平均碳储量。

表2 树种含碳系数Table 2 Carbon coefficient of species
3 结果与分析
3.1 林木碳储量的树种结构特性
采用重要值法计算,按照重要值大小排列得出13种优势树种分别为马尾松、杉木、枫香、短柄枹、白栎、苦槠、山矾、锥栗、麻栎、黄连木、木荷、化香、山合欢[20]。计算各优势树种碳储量得到图1,可以看出马尾松碳储量最高,约占林分碳储量的3/5,且远远超出其他树种。杉木、白栎、枫香、短柄枹碳储量相对较高。由图2可以看出,林木平均单株碳储量中黄连木最高,其次是马尾松和白栎,且其平均单株碳储量均远远超过其他树种,其余树种单株碳储量差别不大。由此可知,马尾松、白栎是天目山针阔混交林林分中主要的固碳树种,针叶树种的碳汇效应优于阔叶树种。
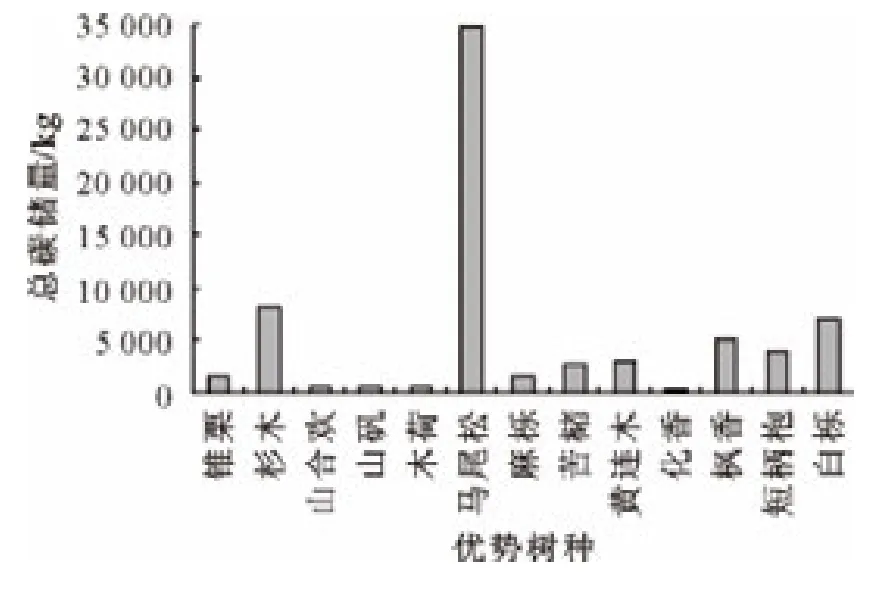
图1 各优势树种总碳储量Fig.1 Total carbon storage of every dominant species
3.2 主要树种林木碳储量的直径结构特性
生物量在林木各径级中的分配比例是群落演替过程中的重要参数之一,是反映森林生态系统动态平衡的标志,碳储量随径阶分布主要受个体数量和胸径大小2个因素影响[21]。以4cm划分径级,整体上看针阔混交林碳储量的径级分布呈“中间大两头小”的规律(图3),即各优势树种碳储量在小径级和大径阶内出现最小值,中等径阶出现最大值。马尾松碳储量在32~40cm出现高峰值,锥栗、山合欢、木荷、苦槠、化香、短柄枹和山矾的碳储量在6~14cm出现高峰值,这7种阔叶树种都是在较小径阶内达到碳储量的高峰值,说明锥栗、山合欢、木荷、苦槠、化香、短柄枹和山矾株数在小径阶内占有非常大的比例。杉木、麻栎、黄连木、枫香、和白栎都存在多峰现象,均在较大、较小径阶处出现碳储量的高峰值。从总体来看,小径级和中径级株数占林分的绝大部分,48cm径阶以上的较大林木株数较少。
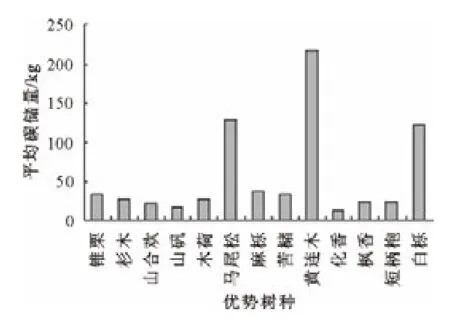
图2 各优势树种平均碳储量Fig.2 Average carbon storage of every dominant species
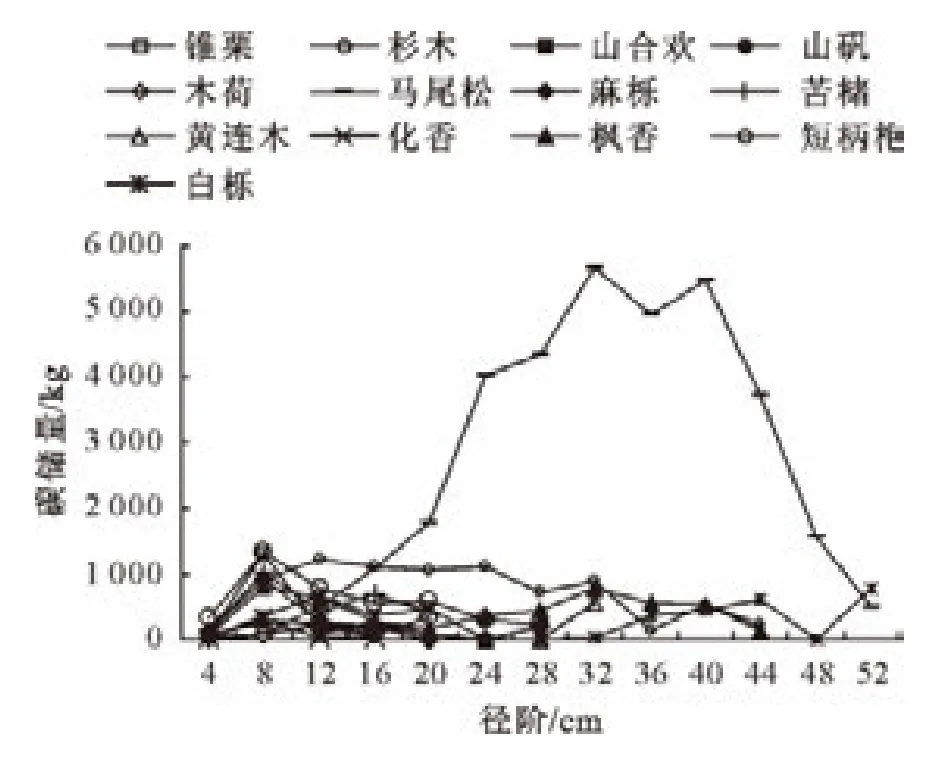
图3 各优势树种碳储量径级分布Fig.3 Carbon storage allocations in each DBH class of dominant species
3.3 主要树种林木碳储量的树高结构特性
以4m划分树高级,伴随着树高高度的增加各优势树种碳储量都存在着明显的变化趋势(图4),大致表现为碳储量的分布沿树高级的增加而增长,至1个峰值后,又随高度的增加而下降,整个变化趋势呈现1个近似正态的峰状曲线。大多数优势树种均在6~10m处达到碳储量高峰。在碳储量达到高峰后,碳储量曲线伴随高度增加而呈现的下降趋势,主要是因为随高度增加树种株数减少的缘故。白栎碳储量随树高增加而增加,尚未出现递减趋势,虽然它受株数随树高等级增加而随之减小的影响,但因其有较大的胸径,致使出现此结果。预计在未来数年内,它仍将出现同其他优势树种一样的分布。

图4 各优势树种碳储量树高级分布Fig.4 Carbon storage allocations in each tree height class of dominant species
3.4 主要树种林木碳储量的冠幅结构特性
以2m划分冠幅级,伴随着冠幅的增加各优势树种碳储量变化趋势表现为碳储量的分布沿冠幅级的增加而增长(图5),至1个峰值后,又随高度的增加而下降,整个变化趋势呈现1个近似正态的峰状曲线,与树高级分布相似。大多数优势树种均在2~6m处达到碳储量高峰。碳储量曲线伴随冠幅增加而呈现的下降趋势,主要是因为随冠幅增加树种株数减少的缘故。
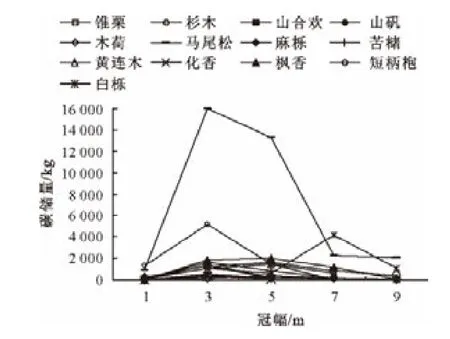
图5 各优势树种碳储量冠幅级分布Fig.5 Carbon storage allocations in each crown width class of dominant species
4 结论
天目山针阔混交林林分中主要的固碳树种为马尾松和白栎,针叶树种的碳汇效应优于阔叶树种。林分中各树种碳储量的径级分布总体呈“中间大两头小”的特征,锥栗、山合欢、木荷、苦槠、化香、短柄枹和山矾等树种的株数在小径阶处占有较大比例。从总体来看,小径级和中径级株数占林分的绝大部分,48cm径阶以上的较大林木株数较少。各树种的碳储量在不同树高级和冠幅级处变化较为相似,均为各优势树种碳储量的分布随着度量级的增加而增长,至1个峰值后,又随度量级的增加而下降,整个变化趋势呈现近似正态的峰状曲线。各树种碳储量在达到高峰值后,碳储量曲线伴随高度和冠幅增加而呈现的下降趋势,主要是因为随高度和冠幅增加树种株数减少的缘故。
针阔混交林是浙江省重要的森林类型,充分发挥针阔混交林的碳汇功能,对改善浙江的生态环境及社会经济的可持续发展至关重要。天目山针阔混交林林木碳储量的树种分布及其随径级、树高级、冠幅级变化的特性研究结果表明,林分中碳储量分布总体处于比较合理的结构状态,碳汇效应较高。在针阔混交林,树种种类虽然较多,但只要进行合理的空间和时间秩序配置,各树种均能处于比较良好的生长状态,各自形成良好的径级、树高级、冠幅级结构,并有良好的生长发育态势。营造针阔混交林应该可以成为碳汇林建设的主要森林类型之一。
[1] 庞宏东,王晓荣,张家来,等.湖北省不同森林类型和不同地区间林下灌木层生物量和碳密度特征[J].西北林学院学报,2014,29(6):46-51.PANG H D,WANG X R,ZHANG J L,et al.Characteristics of shrub layer biomass and carbon density in different forest types and different regions in Hubei Province[J].Journal of Northwest Forestry University,2014,29(6):46-51.(in Chinese)
[2] 闫平,冯晓川.原始阔叶红松林碳素储量及空间分布[J].东北林业大学学报,2006,34(5):23-25.YAN P,FENG X C.Spatial distribution and carbon storage in primitive broad leaved Korean pine forests[J].Journal of Northwest Forestry University,2006,34(5):23-25.(in Chinese)
[3] 王棣,佘雕,张帆,等.森林生态系统碳储量研究进展[J].西北林学院学报.2014,29(2):85-91.WANG D,SHE D,ZHANG F,et al.Advances in the researches carbon storage of forest ecosystems[J].Journal of Northwest Forestry University,2014,29(2):85-91.(in Chinese)
[4] 李轩然,刘琪璟,胡理乐,等.不同方法计算湿地松林生物量的比较[J].生态学杂志,2006,25(12):1594-1598.LI X R,LIU Q J,HU L L,et al.Calculation of Pinus elliotii biomass:a comparison of different methods[J].Chinese Journal of Ecology,2006,25(12):1594-1598.(in Chinese)
[5] 姜鹏,任佳佳,任仙,等.典型针阔混交林中山杨生物量和碳储量研究[J].西北林学院学报,2014,29(1):15-20.JIANG P,REN J J,REN X,et al.Biomass and carbon storage of Populus davidianain typical theropencedrymion[J].Journal of Northwest Forestry University,2014,29(1):15-20.(in Chinese)
[6] 李佳,邵全琴,刘纪远.江西省兴国县森林碳储量动态变化特征[J].西北林学院学报,2012,27(2):163-168.LI J,SHAO Q Q,LIU J Y.Characteristics of spatio-temporal dynamic changes of the carbon storage of forest vegetation in Xingguo county[J].Journal of Northwest Forestry University,2012,27(2):163-168.(in Chinese)
[7] 吴鹏飞,朱波,刘世荣,等.不同林龄桤-柏混交林生态系统的碳储量及其分配[J].应用生态学报,2008,19(7):1419-1424.WU P F,ZHU B,LIU S R,et al.Carbon storage and its allocation in mixed alder-cypress plantations at different age stages[J].Journal of Applied Ecology,2008,19(7):1419-1424.(in Chinese)
[8] 叶功富,吴锡麟,张清海.沿海防护林生态系统不同群落生物量和能量的研究[J].林业科学,2003,39(1):9-14.YE G F,WU X L,ZHANG Q H.Study on biomass and energy of different communities on the coastal forest ecosystem[J].Scientia Silvae Sinicae,2003,39(1):9-14.(in Chinese)
[9] 唐守正,张会儒,胥辉.相容性生物量模型的建立及其估计方法研究[J].林业科学,1999,12(1):53-59.TANG S Z,ZHANG H R,XU H.Study on establish and estimate method of compatible biomass model[J].Scientia Silvae Sinicae,1999,12(1):53-59.(in Chinese)
[10] 韦善华,覃静,朱贤良,等.南宁地区灰木莲人工林生长规律研究[J].西北林学院学报,2011,26(5):174-178.WEI S H,QIN J,ZHU X L,et al.Growth regularity of Manglietia glauce plantation growing in Nanning[J].Journal of Northwest Forestry University,2011,26(5):174-178.(in Chinese)
[11] 刘丽颖,张绍轩,任佳佳,等.30年生不同生长势杉木的胸径和树高的生长动态[J].西北林学院学报,2014,29(6):171-175.LIU L Y,ZHANG S X,REN J J,et al.DBH and tree height growth dynamics of Cunninghamia lanceolata of different growth degree[J].Journal of Northwest Forestry University,2014,29(6):171-175.(in Chinese)
[12] 张连金,惠刚盈,孙长忠.天然红松阔叶林径级结构模拟[J].西北林学院学报,2014,29(6):157-163.ZHANG L J,HUI G Y,SUN C Z.Modeling diameter distribution of natural broadleaved-korean pine mixed forest[J].Journal of Northwest Forestry University,2014,29(6):157-163.(in Chinese)
[13] 赵俊卉,亢新刚,张慧东,等.长白山主要针叶树种胸径和树高变异系数与竞争因子的关系[J].应用生态学报,2009,20(8):1832-1837.ZHAO J H,KANG X G,ZHANG H D,et al.Relationships between coefficient of variation of diameter and height and competition index of main coniferous trees in Changbai Mountains[J].Journal of Applied Ecology,2009,20(8):1832-1837.(in Chinese)
[14] 符利勇,孙华,张会儒,等.不同郁闭度下胸高直径对杉木冠幅特征因子的影响[J].生态学报,2013,33(8):2434-2443.FU L Y,SUN H,ZHANG H R,et al.Effects of diameter at breast height on crown characteristics of Chinese fir under different canopy density conditions[J].Acta Ecologica Sinica,2013,33(8):2434-2443.(in Chinese)
[15] 汤孟平,徐文兵,陈永刚,等.天目山近自然毛竹林空间结构与生物量的关系[J].林业科学,2011,47(8):1-6.TANG M P,XU W B,CHEN Y G,et al.Relationship between spatial structure and biomass of a close-to-nature Phyllostachys edulis stand in Tianmu Mountain[J].Scientia Silvae Sinicae,2011,47(8):1-6.(in Chinese)
[16] 汤孟平,周国模,施拥军,等.天目山常绿阔叶林优势种群及其空间分布格局[J].植物生态学报,2006,30(5):743-752.TANG M P,ZHOU G M,SHI Y J,et al.Study of dominant plant populations and their spatial patterns in evergreen broadleaved forest in Tianmu Mountain[J].Journal of Plant E-cology,2006,30(5):743-752.(in Chinese)
[17] 袁位高,江波,葛永金,等.浙江省重点公益林生物量模型研究[J].浙江林业科技,2009,29(2):1-5.YUAN W G,JING B,GE Y J,et al.Study on biomass model of key ecological forest in Zhejiang Province[J].Zhejiang Forestry Science and Technology,2009,29(2):1-5.(in Chinese)
[18] 应宝根,袁位高,葛永金,等.浙江省重点公益林松类生物量模型研究[J].浙江林业科技,2008,28(2):1-5.YING B G,YUAN W G,GE Y J,et al.Establishment of the biomass model for pine in key ecological forest in Zhejiang[J].Zhejiang Forestry Science and Technology,2008,28(2):1-5.(in Chinese)
[19] 王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究[J].应用生态学报,2001,12(1):13-16.WANG X K,FENG Z W,OUYANG Z Y.Vegetation carbon storage and density of forest ecosystems in China[J].Journal of Applied Ecology,2001,12(1):13-16.(in Chinese)
[20] 王敬,韦新良,徐建,等.天目山区针阔混交林树种测树因子特征研究[J].华东森林经理,2014,28(1):35-40.WANG J,WEI X L,XU J,et al.Survey of species measurement factor in Tianmu Mountain on coniferous-broadleaved mixed forest[J].East China Forest Management,2014,28(1):35-40.(in Chinese)
[21] 姜慧泉,张会儒,亢新刚.长白山阔叶次生林主要乔木地上碳储量分布[J].林业资源管理,2009(5):58-63.JIANG H Q,ZHANG H R,KANG X G.Above-ground carbon storage allocations of main tree species in broad-leaved natural secondary forest of Changbai Mountains[J].Forest Resources Management,2009(5):58-63.(in Chinese)
猜你喜欢
广西林业科学(2022年4期)2022-11-07
安徽农业科学(2022年19期)2022-10-29
林业调查规划(2022年3期)2022-06-13
绿色天府(2022年2期)2022-03-16
林业科技情报(2021年3期)2021-09-01
园艺与种苗(2021年3期)2021-05-26
上海文化(文化研究)(2021年2期)2021-05-08
文化交流(2018年1期)2018-01-09
中南林业调查规划(2017年1期)2017-12-19
现代农业科技(2016年24期)2017-04-19
