
黄土高原丘陵区沙棘人工林土壤微生物群落演变特征研究
2015-01-03 05:12刘金良张顺祥于泽群
西北林学院学报 2015年5期
余 旋,刘 旭,刘金良,张顺祥,于泽群,赵 忠*
(1.西北农林科技大学 林学院,陕西 杨陵712100;2.教育部西部环境与生态重点实验室,陕西 杨陵712100;3.西北农林科技大学 葡萄酒学院,陕西 杨陵712100)
土壤微生物是森林土壤生态系统的重要组成部分之一。它们通过分泌胞外酶,对林木凋落物分解、腐殖质合成及养分循环起着重要调节作用[1-2]。土壤微生物是评价土壤质量的重要指标之一[3],其种群结构的变化不仅能够反映出各个土壤因素对微生物生化特性、生态分布以及功能的影响和作用,还可以反映出微生物对土壤形成演化、土壤肥力维持、植物生长、物质循环以及能量转化的影响和作用。另外,微生物对所生存环境十分敏感,也被公认为是土壤生态系统变化的预警及敏感指标[4-5]。
沙棘是黄土高原丘陵区生态建设的重要树种之一,在保持水土、防风固沙和固氮改土等方面具有重要的生态、经济和社会意义。近年来,我国北方部分地区的沙棘人工林出现了大面积衰败或死亡现象,给当地的生态环境和区域经济发展造成了巨大损失。目前,已有许多学者从水分生理特征[6-7]、病虫害[8]、生态地理分布[9]等方面对此现象进行了研究,尚未得出统一结论。仅有少数学者从土壤生态环境的角度对此进行研究。李玉新[10]等采用Biolog-Eco微平板法对沙棘人工林土壤微生物代谢多样性进行研究,结果表明不同林龄沙棘人工林土壤微生物利用碳源底物的能力存在一定差异。陈金泉[11]等对沙棘人工林土壤氮矿化速率进行研究,认为温度是影响土壤氮矿化的主要因素,季节的影响不明显。因此,拟从沙棘人工林生长发育进程的角度研究其对土壤微生物种群结构的影响,探讨林木生长与土壤环境之间的关系以及沙棘人工林地土壤肥力的影响机理,旨在为黄土高原丘陵区沙棘人工林的健康评价和科学经营提供依据。
1 材料与方法
1.1 研究区概况
试验地点位于陕西省永寿县渡马乡马莲滩流域,107°56′-108°20′E、34°29′-34°59′N,海 拔1 116~1 276m。该区属于温带大陆性季风气候,年均气温10.8℃,年均降雨量601.6mm,年均无霜期210d。地带性土壤为褐土、黄墡土和黑垆土,还有少量的红土、潮土、淤土、石渣土等。本区植被属暖温带落叶阔叶林,森林植被破坏殆尽,残存有少量辽东栎(Quercus liaotungensis)、山杨(Populus davidiana)、白桦(Betula platyphylla)、侧柏(Platycladus orientalis)等天然次生林。人工林以刺槐(Robinia pseucdoacacia)、油松(Pinus tabulaeformis)、侧柏(Platycladus orientalis)、沙棘(Hippophae rhamnoides)为主。
1.2 研究方法
1.2.1 样地设置与样品采集 本试验于2013年7月在陕西省永寿县马莲滩流域峁顶立地条件相同的地段,选取不同发育阶段(8、13、18a)的沙棘人工林作为样地(表1)。每个样地设置3块20m×20m的样方,共计9块样方。在每个样方内按“S”形线路五点法采取表层土壤(0~10cm),并充分混匀,装入无菌样品袋,用冰盒带回实验室。一部分风干过筛后进行土壤理化性质测定,另一部分过2mm筛后放在4℃冰箱,用于土壤微生物磷脂脂肪酸测定。

表1 不同林龄沙棘人工林样地基本概况Table 1 Basic information of the sampling sites
1.2.2 土壤理化性质测定[12]土壤pH采用电极电位法;有机碳采用重铬酸钾氧化-外加热法;全氮用凯氏定氮法;全磷用NaOH熔融-钼锑抗比色法;全钾采用NaOH熔融-原子吸收法;碱解氮采用扩散法;速效磷用碳酸氢钠浸提-钼锑抗比色法;速效钾用乙酸铵浸提-原子吸收法。
1.2.3 磷脂脂肪酸的分离与气象色谱检测 土壤微生物群落结构分析采用磷脂脂肪酸(Phospholipid fatty acid,PLFA)生物标记法。PLFA的提取与测定采用 White[13]的方法。土样用体积比为1∶2∶0.8的氯仿∶甲醇∶柠檬酸缓冲液振荡提取总脂类,SPE硅胶柱分离得到磷脂脂肪酸,碱性甲酯化后用Agilent6850N气相色谱仪分析PLFA成分,内标为正十九烷酸甲酯(19∶0)。色谱条件为:ULTRA-2柱(25.0m×200μm×0.33μm),进样量1μL,分流比10∶1,载气为 N2,流速0.8mL·min-1。二阶程序升高柱温:170℃起始,5℃·min-1升至260℃,而后40℃·min-1升至310℃,维持1.5min。根据Frostagard命名法,以总碳数:双键数和双键距离分子末端位置命名,即:X:YωZ(c/t)。后缀c(cis)和t(trans)分别表示顺式和反式脂肪酸,后缀i(iso)和a(anteiso)分别表示异式支链脂肪酸和反异式支链脂肪酸,cy表示环丙烷脂肪酸。通过查找参考文献,在本论文中采用14:0i,15:0i,15:0a,16:0i,17:0i,17:0a表示革兰氏阳性菌[14];14:1w5c,15:1w6c,16:1w7c,cy17:0,cy19:0,15:03OH,16:12OH,16:1w9c,18:1w5c,cy18:0表示革兰氏阴性菌[14];14:0,15:0,16:0,18:0表示不知名细菌[15];18:1w9c,18:3w6c表示真菌[16-17];16:1w5c表示菌根[18]。
1.3 数据处理
试验数据采用SPSS19.0软件进行方差分析、主成分分析和相关性分析。其中,方差分析采用LSD法检验不同林龄沙棘人工林土壤理化特性、微生物种群结构之间的差异,相关性分析采用Pearson相关系数表示理化特性与土壤微生物种群结构之间的相关性,主成分分析用以解释林龄与土壤微生物种群结构的相互关系。
2 结果与分析
2.1 不同林龄沙棘人工林土壤养分特征
由表2可知,土壤有机碳、碱解氮、速效磷含量随着林龄增加呈显著上升趋势,并于18年生林地中达到最高值,分别较8年生林地高出192.89%、68.06%、85.38%。土壤全氮和速效钾含量在8年生林地较低,分别为1.19g·kg-1和108.75mg·kg-1。随着林分年龄的增加,在13年生林地有显著上升,分别为1.74g·kg-1、104.64mg·kg-1。但是在由13年过渡到18年生的生长发育过程中增长趋缓,且与13年生林地无显著差异。土壤pH值和全钾含量在沙棘人工林发育进程中无显著差异。土壤全磷含量在林分生长发育初期增长缓慢,而在由中龄林过渡到成熟林的进程中增长迅速,18年生林地较13年生高出106.1%。
2.2 不同林龄沙棘人工林土壤微生物群落结构特征
由表3可知,本研究共检测到40种PLFA,其中有17种细菌,2种真菌。总体上来看,随着林龄的增加所测得的PLFA种类和总PLFA量逐渐增加,其中以18年生沙棘人工林测得的PLFA种类最多,为40种,总PLFA量最高,为18.0nmol·g-1干土,显著高于13年和8年生沙棘人工林。细菌总量、革兰氏阳性菌总量和菌根总量在沙棘生长的初期含量较低。而在13年生沙棘林地,三者的量较8年生林地有显著增加。在18年生林地,三者含量虽继续增加,但与13年生林地无显著差异。另外,林分生长发育的进程对革兰氏阴性菌含量有较大影响。革兰氏阴性菌含量随着林龄增加而增加,13年生林地和18年生林地的含量显著高于8年生林。真菌的变化趋势则表现为13年生沙棘人工林真菌的含量最高,为1.41nmol·g-1干土,显著高于8年生沙棘人工林。18年生沙棘人工林的真菌含量虽然有所下降,但与13年生林无显著差异。
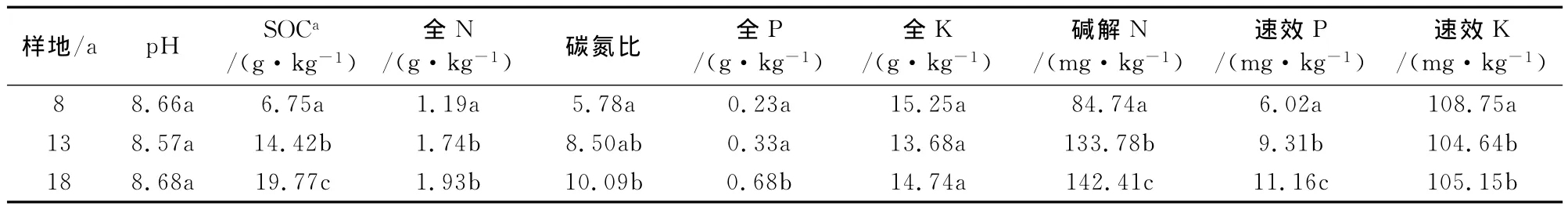
表2 各样地土壤理化特性Table 2 Soil chemical characteristics in different sampling sites

表3 不同林龄沙棘人工林土壤微生物群落特征Table 3 Soil microbial community structure by indicator lipids(nmol·g-1 dry soil)from the sampling sites
由图1可知,主成分分析能区分不同林龄的土壤微生物群落组成。第1主成分解释了微生物群落结构总变异的68.93%,其中单一脂肪酸种类20:1w9c,17:0,18:12OH,18:0,14:0,18:1w5c,16:0i,19:0i,18:1w7c,15:0,10me18:0,10me17:0,19:0cyclow8c,17:0cyclo,17:0i,15:0i,15:0a的载荷值较高。第2主成分解释了微生物群落组成总变异的19.11%,其中单一脂肪酸16:1w5c,14:1w5c,15:1isoG,16:1isoG,18:1w9c,16:12OH 的载荷值较高。这表明林木生长发育进程对土壤微生物群落结构组成影响较大。
2.3 土壤微生物群落结构特征与土壤养分特性的相关性
由表4可知,土壤微生物群落结构的组成与土壤pH无显著相关性。土壤微生物总PLFA量、土壤细菌PLFA量与土壤有机碳、全氮、碳氮比、全磷、碱解氮、速效磷呈显著正相关系。革兰氏阳性菌PLFA量与土壤有机碳、土壤碳氮比、碱解氮、速效磷含量呈显著正相关关系,而与土壤速效钾呈显著负相关。革兰氏阴性菌与所测土壤理化指标(pH、全钾除外)均表现出显著相关。真菌PLFA量与土壤全氮、碱解氮呈显著正相关关系,与土壤速效钾呈极显著负相关关系。菌根真菌PLFA量与土壤有机碳、全氮、碳氮比、碱解氮、速效磷含量呈显著正相关关系,而与土壤全钾、速效钾呈显著和极显著负相关关系。
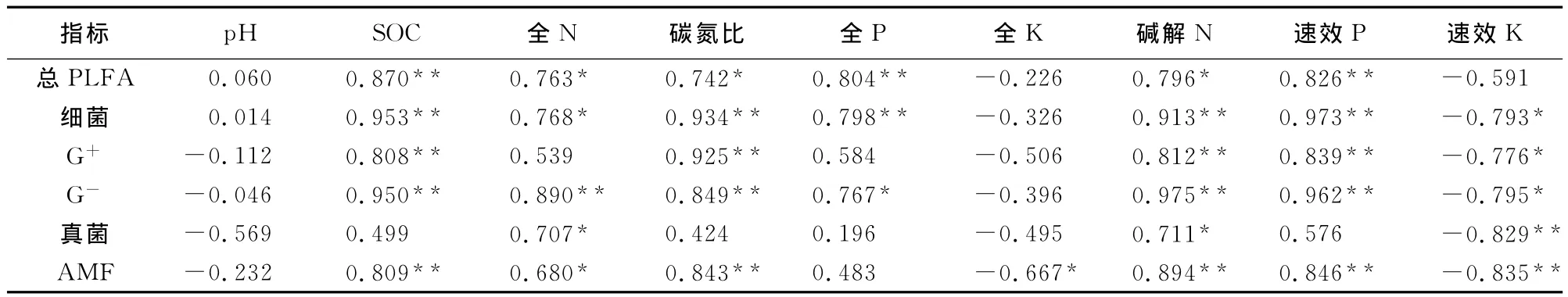
表4 各样地土壤理化特性与微生物群落特征的相关性分析Table 4 Correlation coefficient between the biomass of PLFA groups and soil chemical properties

图1 磷脂脂肪酸主成分分析Fig.1 Principle components analysis of fatty acids
3 结论与讨论
3.1 沙棘人工林的生长发育对理化特性的影响
林木生长与土壤养分相互作用,相互联系。植物通过根系分泌物和残体向土壤提供碳、氮,从而影响土壤有机质的输入,进而显著影响土壤的养分特性[19]。土壤养分作为林木营养的主要来源,直接影响林木的生长发育。本研究结果表明土壤的养分特性随着沙棘人工林的生长发育进程呈现出一定的规律,主要表现为土壤有机碳、全氮、全磷、碱解氮和速效磷显著上升,这可能是由于随着林木生长发育年限的加长,林分枯枝落叶的储量会增加,土壤碳、氮的输入量随之增加,从而影响土壤的其它养分特性。由此可见,人工林的营造有助于恢复和维持土壤肥力,这与刘杰[20]、安韶山[21]、易海燕[22]等的研究结果一致。但也有研究报道表明人工林的土壤容易退化和形成干层[23-24]。这说明人工林的土壤肥力除了受到林分植被这一主要因素的影响外,还受到立地因子,如坡向、坡位、海拔等因素的影响。因此,在研究林分植被对土壤肥力的影响时,还需要研究多因子对土壤的综合作用。
3.2 沙棘人工林土壤微生物种群结构与植被类型
森林植被对土壤微生物种群结构有着显著影响,这是因为植被通过凋落物和根系分泌物向土壤输入碳、氮物质,而土壤碳、氮含量是土壤微生物生长获得营养的重要来源。因此,植被凋落物和根系分泌物的数量和质量决定着土壤微生物种群结构的组成[25]。本研究发现土壤微生物总PLFA量随着林龄的增加而出现递增趋势,且在成熟林时达到最大值。这可能是由于在沙棘幼龄期植物生物量较低,归还到土壤的枯枝落叶和营养物质较少。而此时沙棘生长迅速需要大量的营养物质,具有较强的营养竞争能力,从而抑制土壤微生物的生长,土壤微生物总PLFA量呈现出较低的水平。随着沙棘林的生长,逐步进入中龄林和成熟林期,覆盖度增大,林下植被多样性增加,归还到土壤的养分显著增加,不仅能满足沙棘自身生长发育需要的养分,还能为土壤微生物的生长提供丰富的营养物质,土壤微生物总PLFA量则表现出增加的趋势。
此外,研究结果还发现,不同林龄沙棘人工林土壤微生物种群结构差异很大,且与土壤理化特性,如有机碳含量、全氮含量、碳氮比呈显著正相关关系,这说明土壤理化特性对土壤微生物种群有显著的影响。这与前人的研究结果相一致[26-27]。然而,S.J.Grayston[28]等的研究结果表明土壤有机碳含量、全氮含量与土壤微生物种群结构呈负相关关系。这可能是由于研究所采用的样地土壤类型不一样,其土壤的酸碱度、湿度以及阳离子交换量等物理特性不一样,从而影响土壤养分循环,导致土壤微生物种群出现差异,对碳、氮营养的需求不一致。另外,土壤pH值对土壤肥力,微生物活动、有机质分解以及其他营养元素的释放也起着重要的作用[20]。从前人的研究结果来看,土壤酸碱度对土壤微生物种群结构有显著影响[29]。R.D.Bardgett[30]等研究发现增加土壤酸度有助于提高土壤真菌丰富度,土壤细菌PLFA量随着土壤pH值升高而增加。而本文的研究结果表明,土壤酸碱度对土壤微生物种群直接影响较小。这说明除土壤酸碱度外还有其他环境变量影响土壤微生物种群,土壤酸碱度对微生物种群的影响主要表现为间接影响。
3.3 细菌与土壤环境的关系
在利用PLFA分析土壤微生物种群结构时,单个磷脂脂肪酸物质可以灵敏的反映土壤环境的改变[25]。本研究结果表明,土壤真菌和菌根真菌的含量随着林龄的增加而增加,均在中龄林时期达到最高,这与土壤有机碳的变化趋势一致。一般来说,土壤中真菌含量较高,对应的有机碳含量也较高,这与J.Six[31]等、Q.Priha[32]等的研究结果一致。这可能是由于真菌较细菌可以固定更多土壤中的有机碳,且在真菌居多的土壤中,土壤有机碳的流转较慢。另外,土壤真菌是纤维素和木质素的主要分解者[33]。本研究中土壤真菌逐渐增多,说明随着沙棘生长发育进程的延长,凋落物中难以分解的木质素和纤维素逐渐增多,因此真菌通过庞大的菌丝体,分解不易分解的物质,从而为土壤提供养分。在微生物多样性分析中,细菌与真菌的比值通常被用作描述土壤细菌、真菌在整个土壤微生物群落中所占的相对比例。本试验结果发现,随着沙棘林龄的增长,土壤中细菌与真菌的比值逐渐增大,且在成熟林时达到最高值,说明在沙棘人工林土壤生态系统中,细菌占主导地位,这与Y.S.Cao[25]等在南方巨桉人工林生态系统中所得出的结论一致。Cao[25]等认为这与南方亚热带季风气候有关,高湿高热的环境加速了森林土壤凋落物的分解,从而为细菌的生长繁殖提供有利条件。而在黄土高原丘陵区干旱半干旱气候条件下沙棘人工林土壤生态系统中仍以细菌占主导地位,有待于进一步研究。
[1] 裴雪霞,党建友,张定一,等.不同耕作方式对石灰性褐土磷脂脂肪酸及酶活性的影响[J].应用生态学报,2014,25(8):2275-2280.PEI X X,DANG J Y,ZHANG D Y,et al.Effects of different tillage methods on phospholipid fatty acids and enzyme activities in calcareous cinnamon soil[J].Chinese Journal of Applied Ecology,2014,25(8):2275-2280.(in Chinese)
[2] 余彬彬,金则新,李钧敏.常绿阔叶林次生演替系列群落土壤微生物生物量及酶活性[J].西北林学院学报,2008,23(5):30-33.YU B B,JIN Z X,LI J M.Soil microbial biomass and enzyme activity of soils in different secondary succession stages of evergreen broad-leaved forest[J].Journal of Northwest Forestry University,2008,23(5):30-33.(in Chinese)
[3] 马瑞丰,刘金福,吴则焰,等.格式栲林土壤微生物结构的多样性特征研究[J].西南林业大学学报,2014,34(4):14-19.MA R F,LIU J F,WU Z Y,et al.Soil microbial community structure diversity in Castanopsis kawakamii forest[J].Journal of Southwest Forestry University,2014,34(4):14-19.(in Chinese)
[4] ZELLES L.Fatty acid patterns of phospholipids and lipopolysaccharides in the characterization of microbial communities in soil:a review[J].Biology and Fertility of Soils,1999,29:111-129.
[5] 鹿士杨,彭晚霞,宋同清,等.喀斯特峰丛洼地不同退耕还林还草模式的土壤微生物特性[J].生态学报,2012,32(8):2390-2399.LU S Y,PENG W X,SONG T Q,et al.Soil microbial properties under different grain-for-green patterns in depressions between karst hills[J].Acta Ecologica Sinica,2012,32(8):2390-2399.(in Chinese)
[6] 李孙玲,李甜江,李根前,等.毛乌苏沙地中国沙棘存活及生长对灌水和密度的响应[J].西北林学院学报,2011,26(3):107-111.LI S L,LI T J,LI G Q,et al.The growth and survival of Hippophae rhamniodes subsp.sinensis in response to irrigation intensity and initial density in Mu Us sand-land[J].Journal of Northwest Forestry University,2011,26(3):107-111.(in Chinese)
[7] 代光辉,李根前,李甜江,等.水分条件对中国沙棘生长及种群稳定性的影响[J].西北林学院学报,2011,26(2):1-8.DAI G H,LI G Q,LI T J,et al.Effects of water conditions on the growth and population stability of Hippophae rhamnoides subsp.sinensis[J].Journal of Northwest Forestry University,2011,26(2):1-8.(in Chinese)
[8] 刘永华,章一巧,阎雄飞,等.栎黄枯叶蛾对沙棘林的危害及生物学特性[J].植物保护,2013,39(2):147-151.LIU Y H,ZHANG Y Q,YAN X F,et al.Damage of Trabala vishnou gigantina (Lepidoptera:Lasiocampidae)to the seabuckthorn forest and its biological characteristic[J].Plant Protection,2013,39(2):147-151.(in Chinese)
[9] 李秀寨,李根前,韦宇,等.中国沙棘大面积死亡原因的探讨[J].沙棘,2005,18(1):24-28.LI X Z,LI G Q,WEI Y,et al.Cause of seabuckthorn death in large acreage in China[J].Hippophae,2005,18 (1):24-28.(in Chinese)
[10] 李玉新,赵忠,陈金泉,等.沙棘林土壤微生物多样性研究[J].西北农林科技大学学报:自然科学版,2010,38(8):67-82.LI Y X,ZHAO Z,CHEN J Q,et al.Study on soil microbial diversity of seabuckthorn forest[J].Journal of Northwest A&F University:Nat.Sci.Edi.,2010,38(8):67-82.(in Chinese)
[11] 陈金泉,赵忠,李玉新.不同类型沙棘人工林对土壤氮矿化速率的影响[J].土壤通报,2011,42(3):616-621.CHEN J Q,ZHAO Z,LI Y X.The research of nitrogen mineralization rate of different Hippophae rhamnoides plantation[J].Chinese Journal of Soil Science,2011,42(3):616-621.(in Chinese)
[12] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999.
[13] WHITE D C,DAVIS W M,NICKLES J S,et al.Determination of the sedimentary microbial biomass by extractible lipid phosphate[J].Oecologia,1979,40:51-62.
[14] FROSTEGÅRDÅ,BÅÅTH E.The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil[J].Biology and Fertility of Soils,1996,22:59-65.
[15] BOSSIO D A,FLECK J A,SCOW K M,et al.Alteration of soil microbial communities and water quality in restored wetlands[J].Soil Biology&Biochemistry,2006,38:1223-1233.
[16] YARDWOOD S A,MYROLD D D,Hogberg M N.Termination of belowground C allocation by trees alters soil fungal and bacterial communities in a boreal forest[J].FEMS Microbiology Ecology 2009,70:151-162.
[17] MYERS R T,ZAK D R,WHITE D C,et al.Landscape-level patterns of microbial community composition and substrate use in upland forest ecosystems[J].Soil Science Society of A-merica Journal,2001,65:359-367.
[18] FROSTEGÅRDÅ,TUNLID A,BÅÅTH E.Use and misuse of PLFA measurements in soils[J].Soil Biology &Biochemistry,2011,43:1621-1625.
[19] RUTIGLIANO F A,ASCOLI R D,DESANTO A V.Soil microbial metabolism and nutrient status in a Mediterranean area as affected by plant cover[J].Soil Biology &Biochemistry,2004,36:1719-1729.
[20] 刘杰,马履一,贾忠奎,等.不同林龄华北落叶松林下土壤理化性质及微生物学指标评价[J].水土保持通报,2013,33(6):88-93.LIU J,MA L Y,JIA Z K,et al.Assess of soil physicochemical properties and microbiology indicators in Larix principisrupprechtii plantations[J].Bulletin of Soil and Water Conservation,2013,33(6):88-93.(in Chinese)
[21] 安韶山,黄懿梅.黄土丘陵区柠条林改良土壤作用的研究[J].林业科学,2006,42(1):70-74.AN S S,HUANG Y M.Study on the ameliorate benefits of Caragana korshinskii shrubwood to soil properties in Loess hilly area[J].Scientia Silvae Sinicae,2006,42(1):70-74.(in Chinese)
[22] 易海燕,宫渊波,陈林武,等.岷江上游山地森林/干旱河谷交错带退耕还林后土壤养分变化和微生物分布特征[J].水土保持通报,2010,17(2):130-135.YI H Y,GONG Y B,CHEN L W,et al.Soil nutrient status and microbiological properties in the mountain forests/arid valley ecotone in the upper reaches of Minjiang River after converting farmland to forest[J].Research of Soil and Water Conservation,2010,17(2):130-135.(in Chinese)
[23] 薛萐,刘国彬,戴全厚,等.侵蚀环境生态恢复过程中人工刺槐林(Robinia pseudoacacia)土壤微生物量演变特征[J].生态学报,2007,27(3):909-917.XUE S,LIU G B,DAI Q H,et al.Evolution of soil micro-bial biomass in the restoration process of artificial Robinia pseudoacacia under erosion environment[J].Acta Ecologica Sinica,2007,27(3):909-917.(in Chinese)
[24] 刘占锋,刘国华,傅伯杰,等.人工油松林(Pinus tabulaeformis)恢复过程中土壤微生物量C、N的变化特征[J].生态学报,2007,27(3):1011-1018.LIU Z F,LIU G H,FU B J,et al.Dynamics of soil microbial biomass C,N along restoration chronosequences in pine plantations[J].Acta Ecologica Sinica,2007,27(3):1011-1018.(in Chinese)
[25] CAO Y S,FU S L,ZOU X M,et al.Soil microbial community composition under eucalyptus plantations of different age in subtropical China[J].European Journal of Soil Biology,2010,46:128-135.
[26] DING M M,YI W M,LIAO L Y,et al.Effect of afforestation on microbial biomass and activity in soils of tropical China[J].Soil Biology &Biochemistry,1992,24:865-872.
[27] MENDHAM D S,SANKARAN K V,O'CONNELL A M,et al.Eucalyptus globules harvest residue management effects on soil carbon and microbial biomass at 1and 5years after plantation establishment[J].Soil Biology &Biochemistry,2002,34:1903-1912.
[28] GRAYSTON S J,CAMPBELL C D,BARDGETT R D,et al.Assessing shifts in microbial community structure across a range of grasslands of differing management intensity using CLPP,PLFA and community DNA techniques[J].Applied Soil Ecology,2004,25:63-84.
[29] LUCAS-BORJA M E,CANDEL D,JINDO K,et al.Soil microbial community structure and activity in monospecific and mixed forest stands,under Mediterranean humid conditions[J].Plant and Soil,2012,354:359-370.
[30] BARDGETT R D,HOBBS P J,FROSTEGÅRDÅ.Changes in soil fungal:bacterial biomass ratios following reductions in the intensity of management of an upland grassland[J].Biology and Fertility of Soils,1996,22:261-264.
[31] SIX J,FREY S D,THIET R K,et al.Bacterial and fungal contributions to carbon sequestration in agroecosystems[J].Soil Science Society of America Journal,2006,70:555-569.
[32] PRIHA O,GRAYSTON S J,PENNANEN T,et al.Microbial activities related to C and N cycling and microbial community structure in the rhizospheres of Pinus sylvestris,Picea abies and Betula pendulaseedlings in an organic and mineral soil[J].FEMS Microbiology Ecology,1999,30:187-199.
[33] YANNIKOS N,LEINWEBER P,HELGASON B L,et al.Impact of Populus trees on the composition of organic matter and the soil microbial community in Orthic Gray Luvisols in Saskatchewan(Canada)[J].Soil Biology &Biochemistry,2014,70:5-11.
猜你喜欢
今日农业(2022年13期)2022-09-15
防护林科技(2022年5期)2022-08-22
中国水土保持(2022年3期)2022-03-24
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
西藏农业科技(2019年1期)2019-07-25
中国水土保持科学(2019年6期)2019-04-26
知识经济·中国直销(2018年11期)2018-11-26
现代园艺(2017年19期)2018-01-19
现代园艺(2017年21期)2018-01-03
