
福建三明常绿阔叶次生林物种多样性及群落演替
2015-01-03 05:12:16王俊峰郑小贤
西北林学院学报 2015年5期
王俊峰,郑小贤
(1.北京林业大学 森林资源与环境管理国家林业局重点实验室,北京100083;2.西南林业大学,云南 昆明650224)
常绿阔叶林是我国亚热带地区的地带性植被[1],对维护该地区的生态稳定有重要的作用,但由于长期受到人为活动的干扰,现存的常绿阔叶林退化严重,多为次生林[2]。同时,由于干扰程度的差异以及自然恢复的时间不同,造成了常绿阔叶次生林具有明显的演替动态特征。研究次生群落的演替规律有助于生态系统的恢复与重建[3-4],物种多样性的恢复发展是常绿阔叶林次生林生态系统恢复的关键[5]。而物种的多样性与群落演替动态之间又密切相关[6-7],从多样性的角度研究植物群落的动态或稳定性是生态学研究的一个重要方向[8-11]。群落演替的动态特征可以通过物种多样性这一重要群落信息反映出来,对群落物种多样性的研究有助于更好地认识群落的组成、结构、功能和动态,掌握群落演替的一般规律[12]。
目前,研究常绿阔叶林的演替规律一般是按照草本植物群落-灌草丛-灌丛-针叶林-针阔叶混交林-常绿阔叶林等6个演替阶段进行[13],或者是其中的部分演替阶段[14-17],针对恢复到常绿阔叶林后的演替规律的研究还比较少。通过对不同类型阔叶次生林的群落特征及演替规律进行研究,分析不同森林类型生物多样性之间的差异,及多样性指数与群落演替动态的关系,揭示其演替规律,为研究地区的常绿阔叶次生林的恢复与重建提供参考依据。同时,将研究地区常绿阔叶林划分成不同的演替阶段,可为下一步的森林多功能经营提供依据,并对探索研究地区常绿阔叶林的性质有一定意义。
1 研究区概况
研究区位于福建省三明地区将乐林场,地处武夷山支脉,属低山丘陵地带,位于26°26′-27°04′N、117°05′-117°40′E之间,平均海拔在400~800m左右。属中亚热带季风性山地气候,气候温润,年均温为18.7℃,雨季时间长,雨量充沛但年度分配不均,年均降雨量为1 703.7mm。将乐林场土地总面积为7 113.6hm2,森林覆盖率94.8%,有林地面积6 568.9hm2,其中阔叶林仅13.09%,以生态公益林为主。研究区的阔叶林多为1958年皆伐后自然恢复的林分,火灾、气象灾害等干扰少见,干扰主要来自人为砍伐,现阶段林场阔叶林的经营措施主要是以封山育林为主。
2 材料与方法
2.1 样地调查
2012年7-8月,在研究区内设置20m×30m的样地16块,样地基本情况见表1。以6cm为起测径阶,径阶大小2cm,进行每木检尺,并调查每株树的树种、树高、枝下高、冠幅等。在样地内按照10 m×10m的大小将样地划分成小样方,用皮尺测定每株树木的空间坐标。灌木和草本调查分别在样地内四角和中心位置设置5个小样方,其中灌木样方为5m×5m,草本样方为1m×1m,分别调查样方内每株灌木与草本的物种名称、直径、高度等,并分物种统计总株树,记录每个物种的平均盖度等。
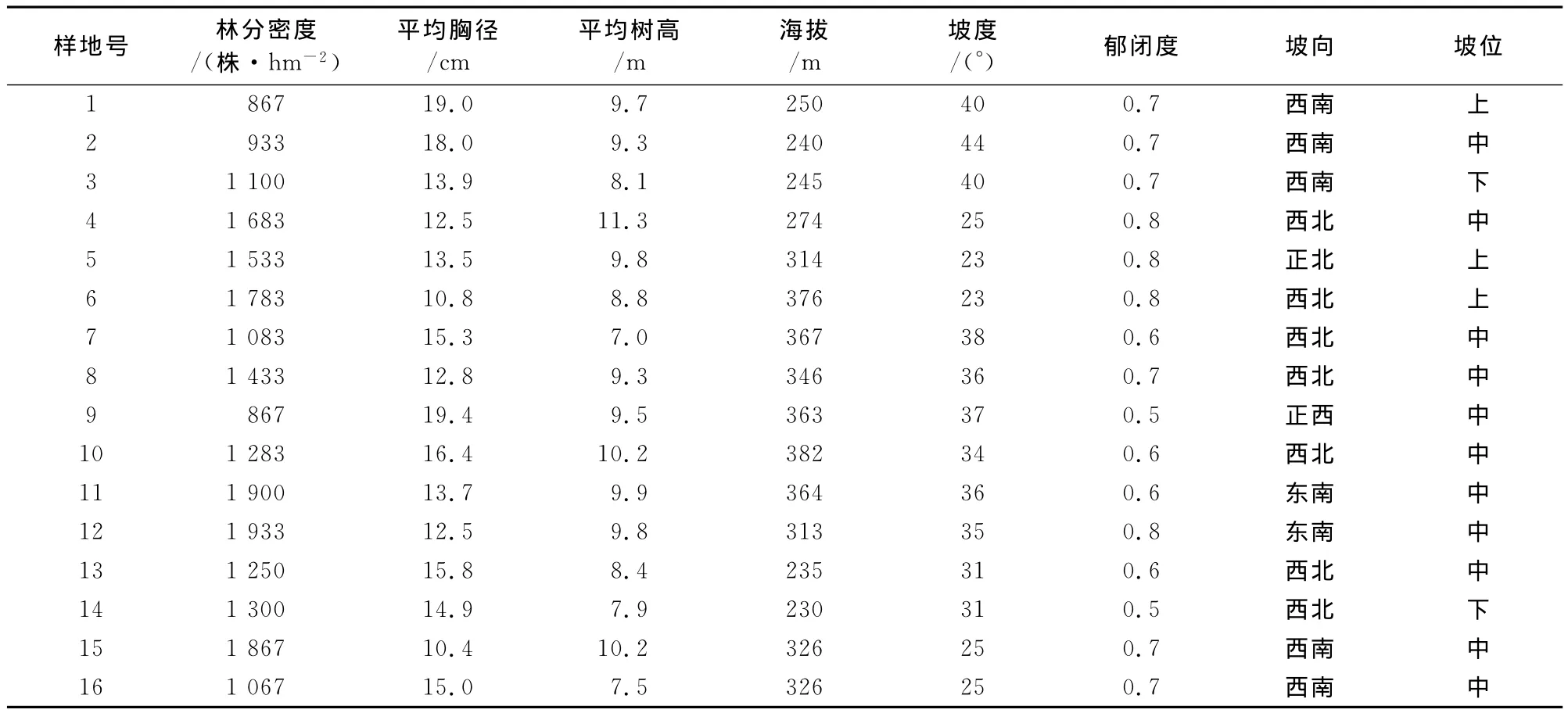
表1 样地基本情况Table 1 Basic conditions of the sample stands
2.2 重要值
重要值(importance value)是以综合数值来表示群落中各植物种的相对重要性,它由相对多度(或相对密度)、相对频度、相对优势度3项特征指标数值综合量化而成,即IV=(相对多度+相对频度+相对优势度)/3。多度用株数表示,频度是某个物种在全部样方中出现的频率,优势度用相对胸高断面积表示。灌木和草本的多度和频度的计算方法与乔木一致,优势度用盖度表示。
2.3 物种多样性
本研究从物种的丰富度指数、多样性指数和均匀度指数3个方面来阐述,其中丰富度指数选取Gleason指数,多样性指数选取辛普森多样性指数(Simpson's diversity index)和 香 农-威 纳 指 数(Shannon-Weiner index),均匀度指数则选取Pielou均匀度指数。
1)物种丰富度(S)
物种丰富度S=出现在样方内物种数量
2)多样性指数
Gleason指数:D=S/lnA
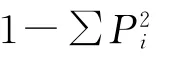
香农-威纳生物多样性指数(Shannon-Weiner diversity index):H′=-∑PilnPi
Pielou均匀度指数:E=H′/lnSmax
式中:A为单位面积或样地面积(m2);Pi为物种i的相对优势度,即Pi=Ni/N,即种i的个体数占群落中总个体数的比例,其中Ni为物种i在群落中的个体数,N为群落中各层的物种个体总数。
3 结果与分析
3.1 群落物种组成
由表2可知,所有调查样地共有植物物种169种,分属57科109属,其中,乔木72种,灌木73种,草本29种,其中灌木层中包含部分乔木的幼树。只含有1种的科有28个,含有5种以上的科有9个:樟科7属15种,壳斗科5属13种,茜草科10属11种,山茶科3属11种,蝶形花科5属10种,紫金牛科4属6种,冬青科1属6种,大戟科4属5种,漆树科3属5种。

表2 样地物种组成Table 2 Species composition of all plots
在乔木层中,苦槠(Castanopsis sclerophylla)为所有样地的共有种,其他分布较广的树种有栲树(Castanopsis fargesii)、拟赤杨(Alniphyllum for-tunei)、青冈(Cyclobalanopsis glauca Oersted)和木荷(Schima superba)。其中,栲树出现在在14个样地中,拟赤杨、青冈出现在13个样地内,木荷出现在12个样地内。乔木层共有72种植物,分属25科46属。其中,只含1种的科有11个,含有3种以上的科有9个:壳斗科5属13种,樟科6属11种,山茶科3属5种,漆树科3属4种,金缕梅科3属3种,大戟科2属3种,冬青科1属3种,山矾科1属3种,柿树科1属3科。
灌木层在样地之间没有共有种,分布较广的物种有杜茎山(Maesa japonica)、毛冬青(Ilex pubescens)分布在15个样地内,山血丹(Ardisia punctata)、网脉酸藤子(Embelia rudis)出现在14个样地内。灌木层共有73种植物,其中部分是乔木的幼苗幼树,分属32科54属。只含有1种的科有18个,含有3种及以上的科有9个:茜草科8属9种,蝶形花科5属7种,山茶科2属6种,樟科4属5种,紫金牛科4属6种。忍冬科2属3种,越桔科1属3种,冬青科1属3种,杜鹃花科1属3种。
草本层在样地之间也没有共有种,分布最广的为狗脊蕨(Woodwardia japonica),在10个样地内出现,其次为芒萁(Dicranopteris dichotoma),在9个样地内出现。草本层共有29种植物,分属16科,23属。其中,只含有1种的科有10个,含有3种以上的科有3个:莎草科2属4种,禾本科含3属3种,鳞毛蕨科2属3种。
3.2 森林类型划分
由表3可知,16块样地的乔木树种以米槠(Castanopsis carlesii)、苦 槠 (Castanopsis sclerophylla)、拟赤 杨 (Alniphyllum fortunei)、黄 樟(Cinnamomum porrectum )、栲 树 (Castanopsis fargesii)、枫香(Liquidambar formosana)为主,以重要值最大的乔木树种作为优势树种,将16块样地划分为6个群落类型,分别为米槠林(样地1),苦槠林(样地2、5),拟赤杨林(样地3、4、6、13、15),栲树林(样地7、9、10、11、12、16),黄樟林(样地8)和枫香林(样地14)。主要灌木种类为小刚竹(Phyllostachys sulphurea)、毛冬青(Ilex pubescens)、杜茎山(Maesa japonica)、箬叶竹(Indocalamus tessellatus),主要草本种类为狗脊蕨(Woodwardia japonica)、华南毛蕨(Cyclosorus parasiticus)、芒萁(Dicranopteris dichotoma)、华 南 紫 萁 (Osmunda vachellii)等。
3.3 物种多样性分析
3.3.1 不同森林类型物种多样性 根据森林类型划分的结果,不同类型包含的样地数量不一致,为便于比较不同森林群落类型多样性之间的差异,将各多样性指数换算到600m2单位面积。不同森林群落类型的多样性结果(表4)表明,各森林类型的物种丰富度和多样性指数都比较高,Simpson指数都在0.9以上。其中,各森林类型的总体Simpson指数排序为栲树林>枫香林>拟赤杨林>苦槠林>黄樟林>米槠林,栲树林的Simpson指数最高,达0.960 2,米槠林的Simpson指数最低,为0.915 3;枫香林的物种丰富度最高为51种,米槠林最低,只有24种;Gleason指数枫香林最高,达到7.972 6,最低的为米槠林。综合各项多样性指数,栲树林、枫香林和拟赤杨林的多样性最为丰富,苦槠林和黄樟林次之,米槠林的多样性最低。各森林类型之间的物种多样性差异并不明显,不同生长环境(坡向、海拔)对多样性指数的影响也不显著。

表3 各样地主要物种及重要值Table 3 Main species and important values of all plots

表4 不同森林类型各层次多样性指数Table 4 Diversity index of different forest types
在各个层次中,乔木层的Simpson指数排序为栲树林>苦槠林>拟赤杨林>枫香林>米槠林>黄樟林,灌木层Simpson指数为栲树林>黄樟林>枫香林>拟赤杨林>苦槠林>米槠林,草本层Simpson指数为拟赤杨林>枫香林>黄樟林>栲树林>米槠>苦槠林。不同森林类型的各个层次的多样性指数顺序差异较大,各多样性指数间的顺序也不完全一致。
3.3.2 不同层次物种多样性 按照乔木、灌木、草本将植物群落划分为3个层次,利用t检验比较各层次间的多样性差异(表5)。由表5可以看出,乔木层与灌木层的物种多样性比较接近,各多样性指数的差异均不显著。米槠林、苦槠林、栲树林的Simpson指数为乔木层>灌木层>草本层,拟赤杨林、黄樟林和枫香林的Simpson指数为灌木层>乔木层>草本层(表4),只有拟赤杨林、黄樟林和枫香林的灌木层的多样性指数是大于乔木层的。草本层在各森林群落的多样性指数都是最低的,且与乔木层、灌木层的差异显著,主要是由于在常绿阔叶林中,林分郁闭度都比较大,林下光照不足,草本层生长空间有限,造成草本植物种类较少且分布不均的情况,导致草本层的物种丰富度及多样性指数都较低。

表5 物种多样性差异性检验(t检验)Table 5 Difference test of species diversity(t-test)
3.4 与其他地区常绿阔叶林物种多样性的比较
Gleason指数不仅能够消除野外调查时样地面积的差异性,描述各植物群落物种多样性的相对大小,还可进行不同地区间的比较。常绿阔叶林的物种多样性随着从北往南纬度的逐渐降低,有逐渐增高的趋势。而研究区的Gleason指数(5.96)却比其更偏北的古田山(6.91)、缙云山(8.69)、乌岩岭(10.30)3个地区[18-19]的常绿阔叶林要小,这3个地区的常绿阔叶林都位于自然保护区内,多样性保护较好,同时,也说明研究区的阔叶林遭到严重的干扰,多样性指数低于其正常水平。
3.5 多样性与群落演替动态的关系
由于上述划分的6种森林类型的多样性指数各指标排序差异较大,不利于分析多样性与演替之间的关系,有必要根据树种生物学特性对6种森林类型进一步归并。在6种森林类型中,米槠林、苦槠林和栲树林3种森林群落都属于壳斗科锥属,属亚热带最为典型的常绿阔叶林类型——栲类林[1];拟赤杨为阳性树种,由于其适应性较强,生长迅速,属于先锋树种;黄樟和枫香为较耐阴树种,属后迁入树种。因此,可以将6种森林类型进一步划分为栲类林、拟赤杨林和其他(黄樟林、枫香林)等3种森林类型。
3种森林类型各层次的多样性指数见表6,各个多样性指数排序一致,都为其他类型(黄樟林、枫香林)的最大,栲类林次之,拟赤杨林最小。在演替的早期阶段,以拟赤杨等为代表的阳性树种在群落中占有主要地位,生物多样性较低,随着演替的进行,耐阴树种(枫香、黄樟、栲类树种等)不断侵入定居,物种不断增加,多样性逐渐升高,当阳性树种和较耐阴的树种同时出现时,群落的多样性达到最大。在演替后期阶段,由于群落中的种间竞争越来越激烈,林分逐渐郁闭,阳性树种生长受到抑制,导致群落的多样性会降低。多数的研究结果也表明从演替早期到后期,群落物种多样性呈现先增加后降低的趋势。根据多样性的变化趋势,结合各森林类型主要树种的生物学特性,可以将3种森林类型划分为3个演替阶段:拟赤杨林处于演替的较早阶段,其他类型(黄樟林、枫香林)处于中间阶段,栲类林处于演替的后期阶段,3个演替阶段的物种多样性呈现先增加后降低的趋势。
草本层多样性的变化对划分演替阶段是也具有重要指示作用,随着演替的进行,林分逐步郁闭,林下光照不足,草本层的多样性会有逐步减小的趋势。由表6中草本层的Simpson指数可以看出,拟赤杨林的草本层的多样性最丰富,其次为枫香林和黄樟林,栲类林的最小。因此,根据草本层的多样性变化趋势也可以得出与上述相同的3个演替阶段。
由于在演替后期多样性指数有下降的趋势,在栲类林的3种森林类型中(表4),栲树林的各层次的多样性都较米槠林、苦槠林两种类型更丰富,可以认为在这3种森林类型中米槠林和苦槠林处于更顶级的演替阶段,米槠林为研究地区的最顶级演替阶段。

表6 3种森林类型各层次多样性指数Table 6 Diversity index of three forest types
4 结论与讨论
根据各样地主要树种组成将研究对象划分为米槠林、苦槠林、拟赤杨林、黄樟林、栲树林和枫香林6种森林类型。其中,拟赤杨与枫香都属于落叶树种,这2种森林类型中,也分布很多壳斗科、樟科等常绿阔叶林的典型树种,是研究区常绿阔叶林的演替中间阶段。因此,本文也把这2种森林类型同归为常绿阔叶次生林。
常绿阔叶林的次生演替一般可划分为5~7个阶段,包括从裸地、灌丛、针叶林到针阔混交林,最后恢复到常绿阔叶次生林阶段,需要100~150a的演替才可达到稳定[20-22]。在常绿阔叶林的整个演替过程中,物种多样性指数对演替有重要的指示作用,Margalef[23]、Odum[24]、Auelair[25]、谢晋阳[9]等的研究都认为物种多样性指数随演替进行而增加,但在随后的演替阶段物种多样性又有降低趋势。本研究主要集中在常绿阔叶林演替的最后一个阶段,属于演替的中后期,该阶段乔木占有主导地位。以乔木树种的优势种将研究区的森林群落划分为米槠林、苦槠林、拟赤杨林、黄樟林、栲树林和枫香林,但6种森林类型的多样性指数的排序差异较大,对划分演替阶段的作用不明显。因此,有必要结合树种的生物学特性将6种森林类型进一步归并为栲类林,拟赤杨林及其他类型(黄樟林、枫香林),3种森林类型的各个多样性指数排序一致,其他类型(黄樟林、枫香林)最大,栲类林次之,拟赤杨林的最小,其物种多样性的变化与整个演替过程的变化一致,呈现先高后低的趋势,拟赤杨林为演替的较早期,枫香、黄樟林为中期,栲类林为演替的后期阶段。
研究区的Gleason指数(5.96)比其更偏北的古田山(6.91)、缙云山(8.69)、乌岩岭(10.30)3个地区的常绿阔叶林要小,说明研究区的阔叶林遭到严重的干扰,多样性指数低于其正常水平。研究区的常绿阔叶林之所以演替到不同的阶段,人为的干扰是重要原因之一,比如拟赤杨林主要分布在集体林中,在恢复过程中又有人为进行间断的破坏,导致其恢复速度较慢,处于演替的较早期阶段,栲类林多分布在人迹罕至,交通不便的山谷,能更好的得到保护,在演替过程中位于较后期的阶段。另外,群落的原生状态也是影响其演替的重要因素,在常绿阔叶林的恢复过程中,当人为干扰停止后,森林会趋向于恢复到受破坏前的原生状态[26]。同时,一些萌生能力较强的树种,如苦槠,青冈在研究区分布较广,在一定程度也会对演替的进行产生重要影响。
研究结果表明,栲类林是该地区常绿阔叶林演替的后期阶段,其物种组成丰富,有利于发挥森林的多重功能。因此,如何促进其他森林类型向该类型进行演替,并缩短其演替过程,是该地区常绿阔叶林经营过程中需要进一步解决的关键问题。可以通过实施抚育,补植等经营措施改善研究地区常绿阔叶林的树种组成,优化空间结构,进一步发挥森林的多重功能。
[1] 吴征鎰.中国植被[M].北京:科学出版社,1980.
[2] 宋永昌,陈小勇,王希华.中国常绿阔叶林研究的回顾与展望[J].华东师范大学学报:自然科学版,2005(1):1-8.SONG Y C,CHEN X Y,WANG X H.Studies on evergreen broad-leaved forests of China:a retrospect and prospect[J].Journal of East China Normal University:Nat.Sci.Edi.,2005(1):1-8.(in Chinese)
[3] 王绪高,李秀珍,贺红士,等.大兴安岭北坡落叶松林火后植被演替过程研究[J].生态学杂志,2004,23(5):35-41.WANG X G,LI X Z,HE H S,et al.Postfire succession of larch forest in the Northern slope of Daxinganling[J].Chinese Journal of Ecology,2004,23(5):35-41.(in Chinese)
[4] 余树全.浙江淳安天然次生林演替的定量研究[J].林业科学,2003,39(1):17-22.YU S Q.The quantitative study on the secondary forest succession in Chun'an county,Zhejiang Province[J].Scientia Silvae Sinicae,2003,39(1):17-22.(in Chinese)
[5] 温远光.常绿阔叶林退化生态系统恢复过程物种多样性的发展趋势与速率[J].广西农业大学学报,1998,17(2):93-106.WEN Y G.Restorative tend and rate of the species diversityof recovering process in degraded ecosystem of evergreen broad leaved forest[J].Journal of Guangxi Agricultural University,1998,17(2):93-106.(in Chinese)
[6] MARGALEF R.Information theory in ecology[J].General Systematics,1957(3):36-71.
[7] 谢晋阳,陈灵芝.暖温带落叶阔叶林的物种多样性特征[J].生态学报,1994,14(4):337-344.XIE J Y,CHEN L Z.Species diversity characteristics of deciduous forests in the warm temperate zone of North China[J].Acta Ecologica Sinica,1994,14(4):337-344.(in Chinese)
[8] 黄建辉,韩兴国.生物多样性和生态系统稳定性[J].生物多样性,1995(3):31-37.HUANG J H,HAN X G.Biodiversity and ecosystem stability[J].Chinese Biodiversity,1995(3):31-37.(in Chinese)
[9] 谢晋阳,陈灵芝.暖温带落叶阔叶林的物种多样性特征[J].生态学报,1994,14(4):337-343.XIE J Y,CHEN L Z.Species diversity characteristics of deciduous forests in the warm temperate zone of North China[J].Acta Ecologica Sinica,1994,14(4):337-343.(in Chinese)
[10] MARGALEF R.Diversity,stability and naturality in natural ecosystems[M].DOBBEN W H,LOWE-MCCONNELL R H,eds.Unifying concepts in ecology.Wageningen:Centre for Agricultural Publishing and Documentation,1975:151-160.
[11] 高贤明,陈灵芝.北京山区辽东栎(Quercus liaotungensis)群落物种多样性的研究[J].植物生态学报,1998,22(1):23-32.GAO X M,CHEN L Z.Studies on the species diversity of Quercus liaotungensis communities in Beijing mountains[J].Acta Phytoecologica Sinica,1998,22(1):23-32.(in Chinese)
[12] 高贤明,马克平,陈灵芝.暖温带若干落叶阔叶林群落物种多样性及其与群落动态的关系[J].植物生态学报,2001,25(3):283-290.GAO X M,MA K P,CHEN L Z.Species diversity of some deciduous broad-leaved forests in the warm-temperate zone and its relations to community stability[J].Acta Phytoecologica Sinica,2001,25(3):283-290.(in Chinese)
[13] 于明坚.青冈常绿阔叶林群落动态研究[J].林业科学,1999,35(6):42-45.YU M J.Dynamics of an evergreen broad-leaved forest dominated by cyclobalanopsis glauca in Southeast China[J].Scientia Silvae Sinicae,1999,35(6):42-45.(in Chinese)
[14] 韩天丰,周国逸,李跃林,等.中国南亚热带森林不同演替阶段土壤呼吸的分离量化[J].植物生态学报,2011,35(9):946-954.HAN T F,ZHOU G Y,LI Y L,et al.Partitioning soil respiration in lower subtropical forests at different successional stages in Southern China[J].Acta Phytoecologica Sinica,2011,35(9):946-954.(in Chinese)
[15] 袁金凤,胡仁勇,慎佳泓.4种不同演替阶段森林群落物种组成和多样性的比较研究[J].植物研究,2011,31(1):61-66.YUAN J F,HU R Y,SHEN J H.Comparison of species composition and diversity of four successional forest communities in Zhejiang Province,East China[J].Bulletin of Botanical Research,2011,31(1):61-66.(in Chinese)
[16] 范玮熠,王孝安,郭华.黄土高原子午岭植物群落演替系列分析[J].生态学报,2006,26(3):706-714.FAN W Y,WANG X A,GUO H.Analysis of plant community successional series in the Ziwuling area on the Loess Plateau[J].Acta Ecologica Sinica,2006,26(3):706-714.(in Chinese)
[17] 唐旭利,周国逸.南亚热带典型森林演替类型粗死木质残体贮量及其对碳循环的潜在影响[J].植物生态学报,2005,29(4):559-568.TANG X L,ZHOU G Y.Coarse woody debris biomass and its potential contri_bution to the carbon cycle in successional subtropical forests of Southern China[J].Acta Phytoecologica Sinica,2005,29(4):559-568.(in Chinese)
[18] 蒋有绪.中国森林群落分类及其群落学特征[M].北京:科学出版社,中国林业出版社,1998.
[19] 胡正华,于明坚,丁炳扬,等.古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究[J].应用与环境生物学报,2003,9(4):341-345.HU Z H,YU M J,DING B Y,et al.Types of evergreen broadleaved forests and their species diversity in Gutian mountain national nature reserve[J].Chinese Journal of Applied Environmental Biology,2003,9(4):341-345.(in Chinese)
[20] 李兴东,宋永昌.浙江东部常绿阔叶林次生演替的随机过程模型[J].植物生态学与地植物学学报,1993,17(4):345-351.LI X D,SONG Y C.The markov process modelling of secondary succession in broadleaved evergreen forest in the eastern part of Zhejiang Province,China[J].Acta Phytoecologica et Geobotanica Sinica,1993,17(4):345-351.(in Chinese)
[21] 丁圣彦.常绿阔叶林演替系列比较生态学研究[M].开封:河南大学出版社,1999.
[22] 丁圣彦,宋永昌.常绿阔叶林演替过程中马尾松消退的原因[J].植物学报,1998,40(8):755-760.DING S Y,SONG Y C.Declining causes of pinus massoniana in the processes of succession of evergreen broan-leaved forest[J].Acta Botanica Sinica,1998,40(8):755-760.(in Chinese)
[23] MARGALEF R.On certain unifying prineiples in ecology[J].Amer.Nat.,1963,97:357-364.
[24] ODUM E P.The strategy of ecosystem development[J].Science,1969,64:262-270.
[25] AUELAIR ALLAN N,GOFF GLENN F.Diversiry relations of upland forests in the Western great lakes area[J].Amer.Nat.,1972,105(946):499-528.
[26] 方运霆,莫江明,彭少麟,等.森林演替在南亚热带森林生态系统碳吸存中的作用[J].生态学报,2003,23(9):1685-1694.FANG Y T,MO J M,PENG S L,et al.Role of forest succession on carbon sequestration of forest ecosystems in lower subtropical China[J].Acta Ecologica Sinica,2003,23(9):1685-1694.(in Chinese)
猜你喜欢
乡村地理(2022年2期)2022-05-30 10:48:04
现代园艺(2021年23期)2021-12-01 07:47:44
环境与生活(2021年5期)2021-06-24 18:09:52
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
绿色中国(2019年18期)2020-01-04 01:57:08
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
当代贵州(2016年23期)2016-07-21 08:55:34
广西林业科学(2016年4期)2016-03-16 05:44:59
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:55
