
葡萄抗寒性综合评价方法的建立
2015-01-02 08:12苏李维李胜马绍英戴彩虹时振振唐斌赵生琴蒲彦涛
草业学报 2015年3期
苏李维,李胜,2*,马绍英,戴彩虹,时振振,唐斌,赵生琴,蒲彦涛
(1.甘肃农业大学生命科学技术学院,甘肃 兰州730070;2.甘肃省干旱生境作物学重点实验室,甘肃 兰州730070)
葡萄(Vitisvinifera)是葡萄属(Vitis)落叶藤本植物。其作为世界上栽培历史悠久的果树之一,具有营养价值丰富、产量高、经济效益显著等特点。常见葡萄除鲜食外,大量用于酿酒、制干、加工等,已成为我国农业产业化的一个重要分支。但近年来,由于我国北方葡萄栽培地区冬季特有的严寒气候,葡萄冻害频频发生,使得众多葡萄生产基地经济损失惨重,因此,选育葡萄抗寒良种并确定抗寒良种的鉴定方法尤为重要。但田间抗寒性研究周期长,可控度低;而室内鉴定常通过人工模拟冰冻试验对葡萄生理生化指标和生物物理指标的测定来比较,可以有效弥补田间试验的不足。
果树的抗寒性综合评价是现代果树抗性育种工作的重要内容之一。关于果树的抗寒性评价,国内外学者已经提出了一些与抗寒性有关的生理生化指标,如:可溶性糖、可溶性蛋白、电导率等。由于果树的抗寒性是许多数量或质量遗传基因综合作用的结果,单一指标评价葡萄抗寒性是片面的[1],用多个指标进行综合性评价才是科学的[2]。目前,葡萄抗寒性的鉴定方法及抗寒生理的研究已有诸多报道。杜永吉等[3]对结缕草(Zoysiasp.)抗寒性进行了综合评价。张倩等[4]对5个葡萄种群18个葡萄品种根系的抗寒适应能力和抗寒适应性观察,通过Logistic方程确定其拐点温度,为将LT50作为葡萄抗寒种质的筛选指标提供了理论和实践依据。Lindén[5]运用Logistic方程,成功分析了植物茎干的冻害数据。艾琳等[6]将不同葡萄品种经低温胁迫10h后,测定根系的细胞膜透性和可溶性糖含量及脯氨酸含量变化,得出细胞膜透性、可溶性糖含量、脯氨酸含量变化与抗寒性的关系。张文娥等[7]应用隶属函数法对葡萄属12个种的膜透性变化、含水量动态变化及萌芽能力进行了综合评价。曹建东等[8]通过模拟低温处理,研究了7个葡萄砧木的萌芽率、生根率、愈伤组织率、电导率、可溶性糖、可溶性蛋白、游离脯氨酸和丙二醛含量与抗寒性的关系,并通过主成分分析对葡萄的抗寒性生理指标进行了筛选,对不同葡萄品种的抗寒性进行了综合评价。但对葡萄抗寒性运用Logistic方程结合隶属函数法的综合评价却鲜有报道。
本文研究了各品种葡萄枝条在低温胁迫过程中可溶性糖、可溶性蛋白、丙二醛、游离脯氨酸、组织含水量、相对电导率等抗寒性相关指标的变化,确定判断葡萄抗寒性的主要指标及衡量抗寒能力的低温有效范围。并通过主成分分析结合隶属函数法对葡萄抗寒能力进行鉴定,以期为葡萄的抗寒性评价提供一种科学的方法。
1 材料与方法
1.1 材料采集
于2013年11月上旬取材于嘉峪关紫轩葡萄基地,采集露地栽培、长势良好且一致的一年生枝条,每品种50枝,长度约50cm,在实验室剪口封蜡后,置于10℃条件下贮藏备用。采集品种有‘双优’(Shuangyou)、‘北冰红’(Beibinghong)、‘左优红’(Zuoyouhong)、‘黑比诺’(Pinot noir)、‘贝达’(Beda)、‘5C’、‘5BB’。
1.2 材料处理
于2013年11月上旬,将供试一年生枝条,用蒸馏水冲洗干净,选择充分成熟、粗度相对一致的健壮枝条分割成段,每段长约15cm,剪口用石蜡封闭。每个葡萄品种分6份,每份约12段,将同一条件处理的各品种葡萄枝条用干净纱布包好,放入泡沫盒中,置于SANYO超低温冰箱中。分别在-5、-15、-25、-35、-45℃等不同温度模拟低温处理[9],并以常温10℃处理为对照。以4℃/h的速率降至目的温度后持续12h。之后逐步升温至10℃放置8h。然后进行相关指标的测定,每个指标测定采用随机取样。
1.3 测定指标
相对电导率用电导仪测定[10],可溶性糖含量用蒽酮法测定,可溶性蛋白采用考马斯亮蓝法测定,丙二醛含量采用硫代巴比妥酸法测定,脯氨酸含量采用茚三酮比色法测定[11],含水量采用烘干法测定。运用Logistic方程计算各品种葡萄枝条的半致死温度(LT50)。
1.4 数据处理
以未经低温胁迫处理(10℃放置)的枝条测定值为对照值,以各低温处理后各主要抗寒性指标的测定值为处理值,如式(1)计算变化率α,对变化率α进行标准化处理,并进行主成分分析。
利用Logistic方程[12],如式(2)。其中Y为电导率,a、b为方程参数,K为曲线的渐近线,即最大电解质透出率,求曲线的拐点温度LT50,即低温半致死温度。

其他数据采用Microsoft excel 2003和SPSS 17.0进行统计与分析,并用Duncan’s新复极差法进行差异显著性检验。
抗寒性综合评价隶属函数公式为[13]:
隶属函数值计算公式:R(Xij)=(Xij-Xjmin)/(Xjmax-Xjmin)
反隶属函数值计算公式:R(Xij)=1-(Xij-Xjmin)/(Xjmax-Xjmin)
式中,R(Xij)表示i种类j指标的抗寒隶属函数值;Xij表示i种类j指标的测定值;Xjmin表示所有种类j指标的最小值;Xjmax表示所有种类j指标的最大值;i表示某个品种;j表示某指标。
2 结果与分析
2.1 相对电导率与抗寒性关系
植物细胞电解质的外渗程度可用相对电导率来表示,来反映植物细胞膜系统的低温伤害程度[14]。由图1可知,各品种葡萄枝条相对电导率随处理温度的降低呈慢-快-慢的增加趋势,说明低温处理使细胞膜受损程度加重。在-15~-35℃之间,各品种葡萄枝条相对电导率显著增加(P<0.05),但不同葡萄品种相对电导率的增加幅度不同,且双优相对电导率变化量明显低于其他各品种。而经-35℃后继续低温处理时,由于-35℃已超过大多品种葡萄的临界温度LT50,葡萄枝条在-35~-45℃之间电导率变化无显著差异(P>0.05),因此,-35~-45℃低温处理段各品种葡萄枝条相对电导率的变化已不能说明葡萄的抗寒能力。故根据-15~-35℃处理阶段相对电导率增幅大小,可知葡萄各品种抗寒性的强弱程度。其抗寒强弱为双优>左优红>贝达>北冰红>5C>5BB>黑比诺。
2.2 可溶性糖含量与抗寒性关系
在逆境条件下,植物体内渗透调节物质大量积累,赋予各种植物渗透调节的能力。可溶性糖是植物细胞内重要的渗透调节物质,能降低细胞溶液的结冰点和减少低温对细胞的伤害,其次,糖分的积累还可增加渗透压,起冰冻保护剂的作用,此外还可提供能量和代谢底物,促进其他与抗寒性相关的生理生化过程的进行。
由图2可以看出,在低温驯化过程中,葡萄枝条组织中可溶性糖的含量总体呈上升趋势。在-15℃处理后,双优的可溶性糖含量最高,其值达1.24%(P<0.05),而黑比诺含量最小,为0.6%(P<0.05)。在10~-15℃的低温处理过程中,可溶性糖含量急剧升高。其中,增速最快为北冰红,其值达0.73%(P<0.05),最小为黑比诺,其值为0.25%(P<0.05),且这几个品种的可溶性糖含量的变化基本表现出与半致死温度一一对应的关系,因此,可溶性糖含量在10~-15℃的变化量可以作为葡萄抗寒性鉴定的主要指标之一。但在-25~-35℃之间,可溶性糖含量急剧下降,且变化显著(P<0.05),可能是由于枝条在低温下,为适应外界环境,机体代谢增强,糖的分解加速所致。而在-35~-45℃之间,可溶性糖的含量又急剧升高(P<0.05),可能由于极限低温刺激机体低温信号,机体感受极限低温以后,诱导可溶性糖合成的相关代谢酶基因表达以及与合成糖类有关的特殊物质生成所致。
2.3 可溶性蛋白含量与抗寒性关系
可溶性蛋白是植物组织的保护物质,能明显增强细胞的持水力,其含量的增加,一方面降低了组织或细胞的冰点温度,同时也使细胞的水合度增大,保水能力增强,避免原生质在低温下的脱水伤害[15]。各葡萄品种可溶性蛋白测定结果如图3所示,在低温胁迫期间,各品种葡萄枝条可溶性蛋白含量变化呈升-降-升的趋势。在低温胁迫初期,可溶性蛋白对温度的变化敏感,随处理温度降低,各品种可溶性蛋白含量急剧升高。经-5℃处理后,可溶性蛋白含量均达到峰值。左优红变化最大,黑比诺变化最小(P<0.05),其余品种差异较大。各品种在10~-5℃之间的变化量与枝条半致死温度LT50基本呈对应关系,因此,选择10~-5℃范围内可溶性蛋白含量的变化量作为葡萄抗寒性鉴定的衡量指标之一。
2.4 丙二醛含量与抗寒性的关系
各葡萄枝条内丙二醛含量的测定结果见图4。由图4可知,葡萄枝条在低温处理初期MDA含量呈缓慢下降趋势,随着胁迫的深入,MDA含量逐渐升高。说明在进行抗寒锻炼的初期,其生理上有一个适应的过程。随着温度的持续降低,各品种中MDA含量呈升高趋势,特别在-15℃处理后,增加趋势更加明显。在-15~-25℃处理期间,所有葡萄枝条的丙二醛含量均呈升高趋势,且变化显著(P<0.05),但双优、左优红的变化较小,表明这时双优、左优红对膜脂过氧化产物的清除能力强,抗寒能力强。而黑比诺的变化趋势最大,表明低温处理后在供试葡萄品种中黑比诺对膜脂过氧化产物的清除能力最弱,抗寒能力最弱,且在-25℃时黑比诺、5BB枝条丙二醛含量最大,表明达到此温度时,葡萄枝条细胞膜脂过氧化程度高,受冻害严重,温度继续下降,组织逐渐死亡,葡萄枝条膜脂过氧化程度下降。此处理段各枝条MDA含量的变化量与半致死温度LT50基本吻合。因此,-15~-25℃处理间MDA的变化量可以作为葡萄抗寒性鉴定的主要指标。

图1 低温处理后枝条相对电导率变化Fig.1 Changes of relative conductivity in shoots after low temperature treatments

图2 不同低温处理可溶性糖含量变化Fig.2 Changes of soluble sugar content after different levels of low temperature treatments

图3 低温处理后枝条可溶性蛋白含量变化Fig.3 Changes of soluble protein contents in shoots after different low temperature treatments
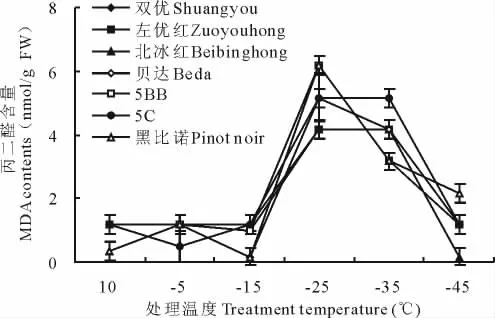
图4 不同低温处理丙二醛含量变化Fig.4 Changes of MDA contents in shoots after low temperature treatments
2.5 脯氨酸含量变化与抗寒性关系
脯氨酸介导植物体内很多细胞和亚细胞的反应,脯氨酸的积累是由于植物体内脯氨酸生物合成激活和脯氨酸降解的抑制共同导致的[16],并且在逆境解除后被迅速分解利用[17]。而植物体内游离脯氨酸能促进蛋白质的水合作用,蛋白质胶体亲水面积增大,能使植物处于低温时免受伤害。各葡萄枝条脯氨酸含量变化如图5所示,各枝条中脯氨酸含量均在原有基础上随外界温度的降低基本呈规律性变化,其变化趋势为升-降-升,但各品种间变化幅度存在一定的差异,由图5可知,双优在10~-25℃之间变化幅度最大,其他品种变化较小,且各品种在10~-25℃之间变化显著(P<0.05),而依据此处理阶段游离脯氨酸的变化量来判断葡萄抗寒性的结果与LT50判断结果基本相同。因此,可以根据10~-25℃之间脯氨酸的变化量来判断葡萄的抗寒性。
2.6 组织含水量变化与抗寒性关系
植物体内的水分常以束缚水和自由水的形式存在。自由水和束缚水含量与植物生长与抗性密切相关。自由水相对含量较高的植物组织或器官,代谢活动旺盛,生长较快,但抗逆性较差;反之则生长较缓慢,但抗逆性较强,由图6可以看出,随处理温度的逐渐降低,枝条组织含水量呈上升趋势。并且在枝条低温处理的各个时期,枝条组织含水量的变化与葡萄抗寒性均无对应关系,因此,11月上旬采摘的葡萄枝条中组织含水量不能作为葡萄抗寒性鉴定的有效指标。

图5 不同低温处理下枝条脯氨酸含量变化Fig.5 Changes of free proline content in shoots after low temperature treatments

图6 不同低温处理下的含水量Fig.6 Changes of water content after different levels of low temperature treatments
2.7 抗寒指标相关性分析
环境胁迫对机体的影响是多方面的。芽是植物对低温最敏感的部位,经过低温处理后,各品种枝条均受不同程度的伤害。随着处理温度的降低,枝条电导率逐渐增大,在植物抗寒研究中,利用电解质外渗50%时的胁迫温度作为植物抗寒强弱的标志,已在抗寒研究中广泛应用。因此,运用Logistic方程计算各葡萄枝条的低温半致死温度LT50,以LT50作为葡萄抗寒性的衡量指标。各抗寒性指标相关性分析结果如表1所示。葡萄的可溶性糖含量、丙二醛含量、游离脯氨酸含量与葡萄的抗寒能力极显著相关,可溶性蛋白含量、相对电导率与抗寒性显著相关。
2.7.1 抗寒指标的选取 根据相关性分析结果,选取与葡萄抗寒性显著或极显著相关的可溶性蛋白含量、相对电导率、可溶性糖含量、丙二醛含量、脯氨酸含量作为评价葡萄抗寒性综合评价的主要指标。在这5个指标中,电导率的变化反映出脂膜透性的大小,随脂膜伤害程度的增大相对电导率也随之增大,从而反映所测葡萄材料抗寒性的强弱。各相关抗寒指标随处理温度的降低,均发生一定变化,且在一定低温处理阶段,都可鉴定葡萄的抗寒性,但不能完全解释葡萄抗寒的实质。因此,以葡萄枝条可溶性糖在10~-15℃、可溶性蛋白在10~-5℃、丙二醛在-15~-25℃、游离脯氨酸在10~-25℃、相对电导率在-15~-35℃的变化量,结合隶属函数法,对葡萄的抗寒性进行综合评价。
低温处理后葡萄枝条各抗寒指标在有效低温内的变化率(α)如表2所示,5个生理生化指标中,抗寒性指标比对照都有所增大(α>0),但变化率的大小有一定的差异,这是由于葡萄品种间差异造成,这些指标都与抗寒性有密切的关系,但由于众变量之间存在一定的相关性,使得观测数据所反映的信息存在重叠现象。故采用主成分分析,将原始指标转化成综合的主成分,能够较好地反映葡萄的抗寒性。
低温胁迫后各指标的变化量利用SPSS 17.0分析软件进行标准化。利用各抗寒性指标的变化率作为综合评价的原始数据,得到标准化数据如表3所示。

表1 葡萄抗寒指标间的相关性分析Table 1 Correlation analysis of cold resistance indexes of grape

表2 低温处理后葡萄枝条各抗寒指标的变化率(α)Table 2 The change rate of each cold resistance index of grape branches in low temperature process

表3 标准化数据Table 3 Standardized date
利用SPSS 17.0对标准数据进行主成分分析,如表4所示,葡萄抗寒性与可溶性糖含量、可溶性蛋白含量、游离脯氨酸含量呈正相关;而与丙二醛含量、相对电导率呈负相关。第1主成分的特征根大于1,且对综合抗寒性的贡献率达到73.225%,可以反映大部分的信息,所以提取第1主成分对葡萄抗寒性作综合评价。
2.7.2 抗寒指标的权重 通过第一主成分中各指标负荷量F与贡献值Y计算各抗寒性指标对葡萄抗寒性的作用大小,确定权重Wi,如表5所示。对葡萄抗寒性影响由强到弱的指标依次是:相对电导率、可溶性蛋白含量、丙二醛含量、脯氨酸含量、可溶性糖含量。

表4 主成分分析结果Table 4 The result of principal components analysis

表5 葡萄枝条各抗寒性指标的负荷量和权重Table 5 Capacity and weight of cold resistance indicator of different variety branches

表6 葡萄枝条各抗寒指标的隶属度值Table 6 Subordination value of cold resistance indicator of different variety branches
2.8 葡萄抗寒性指标的隶属度值与综合评价指标
由于各指标的单位、性质和数量不同,需要进行标准数量化。各指标的变化具有连续性,采用隶属度函数法进行标准数量化,根据主成分因子负荷量的正负确定隶属函数的升降顺序。
根据各指标隶属度值与权重,通过加乘法则Y=Wij×R(Xij),Wij为各指标权重,R(Xij)为各指标隶属度值,对各指标进行合成,得到各葡萄品种的抗寒性综合指数Y。如表6所示。根据抗寒性综合指数的大小,不同葡萄品种抗寒性强弱的顺序是:双优>左优红>北冰红>贝达>5BB>5C>黑比诺。
2.9 综合抗寒指数与半致死温度相关性分析
半致死温度是表征植物抗寒性的主要生态因子[18],运用Logistic方程求曲线拐点(相对电导率随处理温度的变化率最大的一点),即半致死温度(LT50),如表7所示,是用来鉴定植物抗寒性的经典方法。葡萄抗寒性综合指数与半致死温度的相关性分析如表8所示,两种方法评价的结果完全一致。
3 讨论
研究表明,相对电导率、可溶性糖含量、可溶性蛋白含量、丙二醛含量、游离脯氨酸含量等指标虽不能直接判断品种间的抗寒性强弱,但都与抗寒性有密切的关系[19-20]。其中,可溶性糖含量的增加可提高细胞的渗透浓度,降低水势,增加保水能力,从而降低冰点,同时,可溶性糖的积累,还为低温处理下机体代谢提供能量;而低温胁迫下可溶性蛋白含量与可溶性糖含量密切相关,它们常作为植物抗寒鉴定的指标;MDA是膜脂过氧化作用的最终产物,其累积量是反映植物遭受逆境伤害程度的重要指标[21];脯氨酸是植物体内的渗透调节物质,并且积累的指数与抗逆性有关[22]。

表7 各葡萄品种的致死性温度Table 7 LT50of different variety branches
本试验通过测定各低温处理后可溶性糖含量、可溶性蛋白含量、丙二醛含量、游离脯氨酸含量、组织含水量、相对电导率等6个相关指标发现,在整个低温处理过程中,可溶性蛋白对低温变化比较敏感,在10~-5℃之间,可以根据可溶性蛋白含量的变化初步判断各品种的抗寒性强弱。而在10~-25℃之间,可溶性糖、游离脯氨酸的变化在一定程度上体现了各品种的抗寒能力。在-15~-25℃范围内MDA的变化量能更准确的反应葡萄的抗寒性。随着低温的深入,相对电导率逐渐增大,根据相对电导率在-15~-35℃处理段的变化量,可知葡萄各品种抗寒性的强弱为双优>左优红>贝达>北冰红>5C>5BB>黑比诺。但植物组织含水量与葡萄抗寒性并无显著相关关系。
根据前人研究,相对电导率的变化反映出脂膜受伤害的程度,可作为抗寒性鉴定的指标,结合Logistic方程求得低温半致死温度(LT50)能更好、更加准确的反应植物的抗寒性。由于植物是一个有机整体,其受到逆境胁迫而引起植物生理生化指标变化的因素是错综复杂的[23],因此,运用单一指标很难反应葡萄抗寒的实质,也不能充分揭示抗寒的本质,为了全面准确的鉴定葡萄的抗寒性,克服单一指标在葡萄抗寒鉴定中的不足,本文根据与LT50的相关性大小确定了主要判断葡萄抗寒性的5个指标:相对电导率、可溶性糖、可溶性蛋白、丙二醛、游离脯氨酸。运用主成分分析方法从原始指标中抽提出更少的新指标,更客观的解释原始指标里所包含的信息[24-25]。本文将衡量葡萄抗寒性的各指标在有效低温衡量范围内的变化率用主成分分析将多个指标转化成综合的主成分,再根据主成分的负荷量确定各个抗寒性指标在葡萄抗寒作用中的权重,结合隶属函数法建立综合评价方法。根据综合抗寒指数Y的大小,得出葡萄抗寒能力的强弱顺序:双优>左优红>北冰红>贝达>5BB>5C>黑比诺。该排序结果与LT50的计算结果完全吻合,也与前人研究结果一致。本试验通过测定低温处理后葡萄枝条各项抗寒生理指标的变化,在多指标测定的基础上运用隶属函数法进行综合评价。
葡萄抗寒性综合评价方法的建立,不仅为葡萄的抗寒性选育和生产发展提供了更加科学的鉴定方法,也为各相关植物抗寒性的研究提供了理论依据。但不同时期葡萄的各个生理指标可能发生变化,对其他时间采集的材料进行抗寒评价时,各生理生化指标与LT50的相关性可能发生变化,需要建立新的综合评价方法进行评价。植物体抗寒机制非常复杂,影响其抗寒性的因素较多,因此产生了多种测定植物抗寒性的方法,目前,国内外测定木本植物抗寒性的主要方法有电解质渗出率法(EL)[26]、全株冰冻测试法[27]、电解质渗出率法[28]、叶绿素荧光法[29]、热分析法[30]等,国内在形态结构、膜脂的成分及蛋白质、抗寒生理生化、抗寒基因等方面的研究已取得显著进展[31]。但在具体评定葡萄抗寒性时,还应多方面综合评价,同时结合田间观察,才能使结果更加准确、科学。
Reference:
[1]Liu X H,He X Y,Su D M,etal.Establishment and application of integrated assessment system on cold resistance of different Zenia insignis Provenances.Scientia Silvae Sinicae,2007,43(10):45-50.
[2]Lv Y D,Dong F X,Wang G X,etal.Establishment and application of integrated assessment systemon cold resistance of Hybrid Hazelnut.Scientia Silvae Sinicae,2008,44(9):31-35.
[3]Du Y J,Yu L,Sun J X,etal.Comprehensive assessment of cold resistance of threeZoysiajaponicavarieties.Acta Prataculturae Sinica,2008,17(3):6-16.
[4]Zhang Q,Liu C H,Guo D L,etal.Relationship between LT50and cold adaptability of five grape varieties.Journal of Northwest A & F University(Natural Science Edition),2013,41(5):149-154.
[5]Lindén L.Measuring Cold Hardiness in Woody Plants[M].University of Helsinki,2002.
[6]Ai L,Zhang P,Hu C Z,etal.Effects of low temperatures tolerance on the electrolyte osmotic rate and the content of soluble sugar,proline of grape roots.Journal of Xinjiang Agricultural University,2004,27(4):47-50.
[7]Zhang W E,Wang F,Pan X J.Comprehensive evaluation on cold hardiness of Vitis species by Subordinate Function(SF).Journal of Fruit Science,2007,24(6):849-853.
[8]Cao J D,Chen B H,Wang L J,etal.Cold resistance indexes identification and comprehensive evaluation of grape varieties.Acta Botanica Boreali-Occidentalia Sinica,2010,30(11):2232-2239.
[9]Lin X S,Lin Z X,Lin D M,etal.Cold-tolerance of 5species of Juncao under low temperature stress.Acta Prataculturae Sinica,2013,22(2):227-234.
[10]Pan X Y,Wang G X,Cao Q D.Winter injury index and lethal low temperature for introducted American Almond in Lanzhou,China.Acta Horticulturae Sinina,2002,29(1):63-65.
[11]Zhang Z L.Plant Physiology Experimental Guidance[M].Beijing.Higher Education Press,2002:127-128,159-160.
[12]Zhu G H,Liu Z Q,Zhu P R.A study on determination of lethal temperature with logistic function.Journal of Nanjing Agricultural University,1986,3(3):11-16.
[13]Zhang J X,Wu X C,Yang Y Z,etal.Comprehensive evaluation of juice making grape cultivars introduced from USA to cold-resistance.Northern Horticulture,2011,(24):1-5.
[14]Lu J X,Jiang H Y,Li W.Effects of low temperature stress on the cold resistance of rootstock and branch of wine grapes.Journal of Fruit Science,2012,29(6):1040-1046.
[15]Song F H,Shi Y J,Wu Z B.Analysis on cold resistance of branches ofZiziphusjujubaMill.Xinjiang Agicultural Sciences,2009,46(6):1212-1215.
[16]Nakashima K,Satoh R,Kiyosue T,etal.A gene encoding proline dehydrogenase is not only induced by proline and hypoosmolarity,but is also developmentally regulated en the reproductive organs ofArabidopsis.Plant Physiology,1998,118:1233-1241.
[17]Trotel-Aziz P,Niogret M F,Deleu C,etal.The control of proline consumption by abscsic acid during os motic stress recovery of canola leaf discs.Physiology Plantarum,2003,117:213-221.
[18]Guo W D,Zhang Z Z,Jiang X W,etal.Semilethal temperature of fingered Citron(Citrusmedicavar.sarcodactylis Swingle)under low temperature stress and evaluation on their cold resistance.Acta Horticulturae Sinina,2009,3(1):81-86.
[19]Xie L F.Studies on Cold Resistance Identification and Mechanism of Grapevine Rootstocks[D].Hohhot:Inner Mongolia Agricultural University,2007.
[20]Yang F X,Jin F,Yan X.Comprehensive evaluation of different strawberry varieties tolerance to coldness.Journal of Fruit Science,2010,27(3):368-372.
[21]Hui Z L,Li C Z,Shi W X,etal.A study on the use of fulvic acid to improve growth and resistance in continuous cropping of potato.Acta Prataculturae Sinica,2013,22(4):130-136.
[22]Xu Y,Wei X H,Li B B,etal.Effects of exogenous nitric oxide on seed germination and seedling oxidative damage inMedicagosativaunder NaCl stress.Acta Prataculturae Sinica,2013,22(5):145-153.
[23]Wang L H,Li X,Liu W.A study on resistance and purifying ability of SO2on four warm-season turfgrasses.Acta Prataculturae Sinica,2013,22(1):225-233.
[24]Zhang W E,Wang F,Pan X J.Hardiness resistance of grape progenies hybridization and self pollination.Acta Agriculturae Boreali-Occidentalis Sinica,2009,18(4):290-294.
[25]Guo A H,Chen Y,Yao Y J,etal.Principle component analysis method of almonds tolerance to cold.Journal of Shanxi Agricultural University(Natural Science Edition),2007,27(3):234-237.
[26]Zhang G,Xiao J Z,Chen D F.Electrical impedance spectroscopy method for measuring cold hardiness of plants.Journal of Plant Physiology and Molecular Biology,2005,31(1):19-26.
[27]Tanaka Y,Brotherton P,Hostetter S,etal.The operational planting stock quality testing program at Weyerhaeuser.New Forests,1997,13:423-437.
[28]Deans J D,Billington F J,Harvey F J,etal.Assessment of frost damage to leafless stemtissues of Quercuspetraea:a reappraisal of themethod of relative conductivity.Forestry,1995,68:25-34.
[29]Maxwell K,Johnson G N.Chlorophyll fluorescence-apractical guide.Journal of Experimental Botany,2000,51:659-668.
[30]Kang S K,Motosugi H,Yonemori K,etal.Supercooling characterics of some deciduous fruit trees as related towater movement within the bud.Journal of Horticultural Science and Biotechnology,1998,73(2):165-172.
[31]Ma Y Z,Liang W B,Chen J H.Advances in researches on plant frost hardiness.Nonwood Forest Research,2005,23(4):89-94.
[1]柳新红,何小勇,苏冬梅,等.翅荚木种源抗寒性综合评价体系的构建与应用.林业科学,2007,43(10):45-50.
[2]吕跃东,董凤祥,王贵禧,等.平欧杂交榛抗寒性综合评价体系的建立与应用.林业科学,2008,44(9):31-35.
[3]杜永吉,于磊,孙吉雄,等.结缕草3个品种抗寒性综合评价.草业学报,2008,17(3):6-16.
[4]张倩,刘崇怀,郭大龙,等.5个葡萄种群的低温半致死温度与其抗寒适应性的关系.西北农林科技大学学报,2013,41(5):149-154.
[6]艾琳,张萍,胡成志.低温胁迫对葡萄根系膜系统和可溶性糖及脯氨酸含量的影响.新疆农业大学学报,2004,27(4):47-50.
[7]张文娥,王飞,潘学军.应用隶属函数法综合评价葡萄种间抗寒性.果树学报,2007,24(6):849-853.
[8]曹建东,陈佰鸿,王利军,等.葡萄抗寒性生理指标筛选及其评价.西北植物学报,2010,30(11):2232-2239.
[9]林兴生,林占熺,林冬梅,等.低温胁迫5种菌草的抗寒性评价.草业学报,2013,22(2):227-234.
[10]潘晓云,王根轩,曹琴东.兰州地区引种的美国扁桃的越冬伤害与临界致死低温.园艺学报,2002,29(1):63-65.
[11]张志良.植物生理学实验指导[M].北京:高等教育出版社,2002:127-128,159-160.
[12]朱根海,刘祖祺,朱培仁.应用Logistic方程确定植物组织低温半致死温度的研究.南京农业大学学报,1986,3(3):11-16.
[13]张剑侠,吴行昶,杨亚州,等.引进美国制汁葡萄品种抗寒性的综合评价.北方园艺,2011,(24):1-5.
[14]鲁金星,姜寒玉,李唯.低温胁迫对砧木及酿酒葡萄枝条抗寒性的影响.果树学报,2012,29(6):1040-1046.
[15]宋锋惠,史彦江,吴正保,等.枣树枝条的抗寒性测定分析,新疆农业科学,2009,46(6):1212-1215.
[18]郭卫东,张真真,蒋小韦,等.低温胁迫下佛手半致死温度测定和抗寒性分析.园艺学报,2009,3(1):81-86.
[19]谢丽芬.葡萄砧木抗寒性鉴定及抗寒机理的研究[D].呼和浩特:内蒙古农业大学,2007.
[20]杨凤翔,金芳,颜霞.不同草莓品种抗寒性综合评价.果树学报,2010,27(3):368-372.
[21]回振龙,李朝周,史文煊,等.黄腐酸改善连作马铃薯生长发育及抗性生理的研究.草业学报,2013,22(4):130-136.
[22]徐严,魏小红,李兵兵,等.外源NO对NaCl胁迫下紫花苜蓿种子萌发及幼苗氧化损伤的影响.草业学报,2013,22(5):145-153.
[23]王丽华,李西,刘尉,等.四种暖季型草坪草对SO2的抗性及净化能力的比较.草业学报,2013,22(1):225-233.
[24]张文娥,王飞,潘学军.葡萄种间杂交和自交后代抗寒性分析.西北农业学报,2009,18(4):290-294.
[25]郭爱华,陈钰,姚月俊.杏品种抗寒性主成分分析.山西农业大学学报,2007,27(3):234-237.
[26]张钢,肖建忠,陈段芬.测定植物抗寒性的电阻抗图谱法.植物生理与分子生物学学报,2005,31(1):19-26.
[31]马英姿,梁文斌,陈建华.经济植物的抗寒性研究进展.经济林研究,2005,23(4):89-94.
猜你喜欢
中老年保健(2022年3期)2022-11-21
今日农业(2021年20期)2021-11-26
河北果树(2020年4期)2020-11-26
河北果树(2020年1期)2020-02-09
科学与财富(2019年15期)2019-10-21
湖南林业科技(2017年1期)2017-02-06
新疆农垦科技(2016年2期)2016-08-21
广西林业科学(2016年2期)2016-03-20
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
