
不同品系高羊茅应答高温胁迫的初级代谢产物分析
2015-01-02 08:12赵状军胡龙兴胡涛傅金民
草业学报 2015年3期
赵状军,胡龙兴,胡涛,傅金民
(中国科学院武汉植物园,植物种质资源创新与特色农业重点实验室,湖北 武汉430074)
高温是限制植物分布、生长和生产力的主要环境因子之一[1]。高温对植物的影响首先表现在表观形态,进而影响植物正常生理代谢。大量研究表明,高温会影响植物体外观形态、光合作用、呼吸作用、细胞膜稳定性、糖、脂及氨基酸代谢等一系列生理生化过程[2-4]。短时间的极端高温会使叶片失水萎蔫枯黄、根系褐变等[5]。长时间的亚高温会使植物发生一系列的生长障碍,如叶片和根系生长受阻、植株早衰,甚至导致植株死亡[6-7]。
代谢组学在植物逆境研究中已经得到了广泛的应用。Cramer等[8]比较了葡萄在脱水和盐胁迫下的代谢谱,分析发现脱水处理的植物含有更高浓度的葡萄糖、苹果酸和脯氨酸。Du等[9]利用GC-MS技术测定分析了C4植物狗牙根(Cynodondactylon)和C3植物草地早熟禾(Poapratensis)在高温胁迫下的代谢产物的差异,认为高温胁迫下不同初级代谢产物的积累量是对高温响应差异的主要原因。Kaplan等[10]借助GC-MS技术研究了其在高温与低温环境下野生型拟南芥(Arabidopsisthaliana)的代谢指纹图谱,分析发现与高温胁迫相比,低温胁迫更显著地影响了野生型拟南芥代谢产物的改变。Rizhsky等[11]的代谢谱分析显示,在剧烈的脱水和热胁迫结合处理的拟南芥中,蔗糖将会取代脯氨酸作为主要的渗透保护剂。Wienkoop等[12]使用GC-MS代谢组学研究拟南芥响应热休克和冷休克的代谢产物变化发现,响应热休克所产生的大部分代谢产物与冷休克响应所产生的代谢产物是重叠的。结果表明包含有脯氨酸、蔗糖、肌醇和棉子糖等相容性溶质的代谢网络对耐受温度胁迫具有重要作用。
高羊茅(Festucaarundinacea)是一种亚热带和温带地区常用的冷季型草坪草种,成坪快,绿期长,抗逆性强,耐酸、耐瘠薄,其抗旱特性和耐热性在冷季型草坪中首屈一指[13-14]。高羊茅无论作为牧草,还是草坪草,都有着十分广阔的应用前景。然而高温严重限制了高羊茅的种植及推广。目前国内外对高羊茅耐热性的研究多数集中在外观形态、生理生化和基因表达变化的研究[14-16],而缺乏从代谢组学方面来研究高温对高羊茅代谢影响的报道。本研究以不同耐热性高羊茅基因型PI 578718(耐热)和PI 234881(热敏感)为试验材料,经人工模拟高温处理,通过表型和生理评价其耐热性的差异,同时对初级代谢产物进行测定分析,以期明确高温对不同耐热性高羊茅初级代谢的影响,从代谢途径上揭示高羊茅的耐热机理,为高羊茅的遗传改良和耐热品种选育提供理论基础和实践指导。
1 材料与方法
1.1 试验材料与培养
试验材料为本实验室通过高温试验筛选出的一个耐热型高羊茅基因型(PI 578718)和一个热敏感型高羊茅(PI 234881)。两份高羊茅材料通过人工扩繁,培养在褐炭土和细沙按质量比1∶1的土壤基质中,间隔2d浇灌半剂量的Hoagland营养液,同时进行自来水喷灌并定期修剪,培养2个月后进行后续高温试验。
1.2 试验方法
1.2.1 试验处理 采用盆栽试验,培养基质为褐炭土和细沙按质量比1∶1混匀。试验在中国科学院武汉植物园的人工智能光照培养箱中进行,培养箱设置处理组温度为白天40℃,晚上30℃;对照组温度为白天25℃,晚上20℃;相对湿度75%;光强600μmol/(m·s);光周期14h/10h。共处理21d,每个处理设置3个重复,分别在0,2,6,14,21d取叶片样进行测定,在0,6,21d取根系样进行测定。
1.2.2 测定指标及方法 草坪质量打分:采用9分制目测评估,评估内容包括色泽、均一性、质地、草坪盖度。9分为叶片绿色均匀,基本无枯黄,质地较好,0分为完全枯黄,6分为可接受的草坪质量。
叶片相对含水量的测定参照邹琦[17]的称量法;相对电导率的测定参照邹琦[17]的电导法;根系活力的测定参照邹琦[17]的TTC法并略加修改。
叶绿素含量的测定参照郭慧娟等[18]的方法。取完全展开的功能叶片0.1g,放入含10mL二甲亚砜(MDSO)的离心管中,黑暗中放置48h(至叶片失绿透明),每天摇匀一次。吸取1mL萃取液于比色皿中,加入二甲亚砜2mL,混匀后,以二甲亚砜为空白对照,使用分光光度计测定OD663、OD645值。并按下列公式计算叶绿素a、叶绿素b和总叶绿素含量。
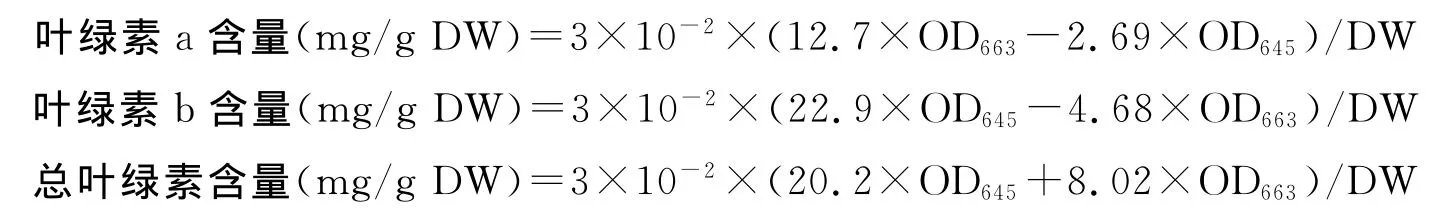
初级代谢产物的测定:代谢产物浸提参照Roessner等[19]的方法并略加修改后,通过GC-MS技术参照Du等[9]的方法对浸提物中的初级代谢产物进行分析。具体流程如下:
1)样品提取:精确称取液氮研磨后的样品0.1g,加入1.4mL 100%的甲醇,同时加入20μL浓度为2 mg/mL的核糖醇作为极性成分的内标,室温提取2h后放入金属浴中70℃恒温15min,然后12000r/min离心5 min,上清液转入10mL离心管中,加入750μL氯仿和1.5mL纯净水,涡旋混匀后于12000r/min离心5min,转移上清液(极性相)移至新离心管中。
2)衍生化:上清液300μL至2mL进样瓶中,在真空浓缩仪(900r/min,30℃,2h)中真空浓缩至完全干燥后加入80μL浓度为20mg/mL盐酸甲氧胺,37℃振荡2h,然后再加入50μL MSTFA至样品中,37℃200r/min振荡2h后进样分析。
3)气相色谱和质谱条件:采用安捷伦5975-7890色谱质谱联用仪测定分析,色谱柱为DB-5ms(30mm×0.25 mm,0.25μm)毛细管柱,载气为氦气(99.999%),流速为1mL/min。进样模式为不分流进样,进样量1μL。升温程序:70℃保持5min,以每分钟5℃的速率升至300℃,温度最终升高至310℃,保持10min,并在下次进样前70℃平衡1min。质谱采用电子轰击(electron impact,EI)离子源,电子能量为70eV,传输线温度为260℃,离子源温度为200℃,扫描速度0.6scan/s,溶剂延迟时间为6min,质量扫描范围为50~600m/z。
将从GC-MS分析仪上得到的数据文件以简单模式方式分析,所有色谱峰通过Varian300自带软件和NIST进行比对,然后运用气质联用化学工作站(Agilent MSD Productivity Chemstation),通过比对出峰时间、荷质比和保留指数对各类初级代谢产物进行鉴定分析。
1.3 数据分析及作图
用SPSS 17.0软件对数据进行方差分析和多重比较,用Origin Pro 9.0软件制图。通过 Multi Experiment Viewer software v.4.8对初级代谢产物进行层次聚类分析(Hierarchical clustering analysis)。
2 结果与分析
2.1 高温胁迫对不同品系高羊茅草坪目测质量和叶绿素含量的影响
热处理之前,热敏感型高羊茅PI 234881的草坪质量要优于耐热型高羊茅PI 578718。高温胁迫下热敏感基因型高羊茅的草坪质量呈显著下降趋势,而耐热基因型只有在胁迫21d后才显著下降。在整个高温胁迫期间,热敏感型高羊茅的草坪质量均显著低于耐热型高羊茅的草坪质量(图1)。
高温导致高羊茅体内叶绿素含量降低(图2)。热处理之前,热敏感型高羊茅的叶绿素含量要显著高于耐热型高羊茅的叶绿素含量。随着高温胁迫的进行,热敏感型高羊茅的叶绿素含量迅速下降,而耐热型高羊茅的叶绿素含量前期变化不大,直到21d时才显著降低。因此,在高温处理14和21d时,热敏感型高羊茅的叶绿素含量要显著低于耐热型高羊茅的叶绿素含量。
2.2 高温胁迫对不同品系高羊茅叶片相对含水量和叶片电解质渗漏的影响
热处理之前,两个高羊茅品种叶片相对含水量差异不大,接近90%(图3)。随着高温胁迫的进行,高羊茅叶片相对含水量降低,尤其是热敏感的高羊茅品种,在高温胁迫下,叶片相对含水量急剧下降,热胁迫21d时,其叶片相对含水量仅为55%,显著低于耐热型高羊茅的叶片相对含水量。

图1 高温胁迫0,2,6,14和21d对不同品系高羊茅草坪质量的影响Fig.1 Effects of heat stress on turf quality in two tall fescue genotypes at 0,2,6,14and 21days after treatment
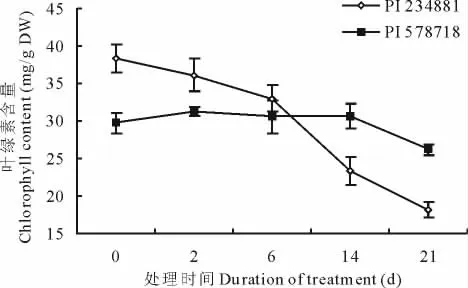
图2 高温胁迫0,2,6,14和21d对不同品系高羊茅叶绿素含量的影响Fig.2 Effects of heat stress on Chl content in two tall fescue genotypes at 0,2,6,14and 21days after treatment
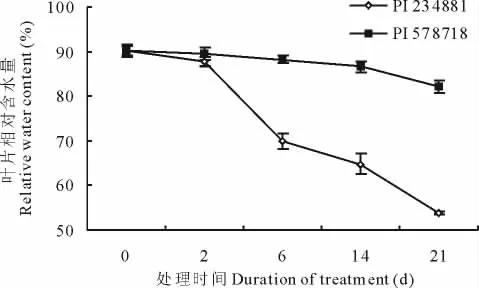
图3 高温胁迫0,2,6,14和21d对不同品系高羊茅叶片相对含水量的影响Fig.3 Effects of heat stress on relative leaf water content in two tall fescue genotypes at 0,2,6,14 and 21days after treatment
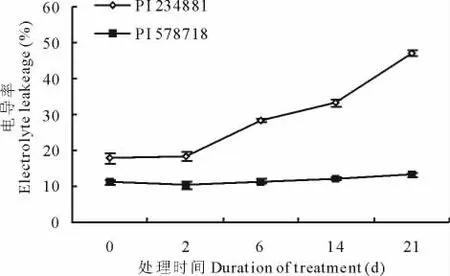
图4 高温胁迫0,2,6,14和21d对不同品系高羊茅叶片电解质渗漏的影响Fig.4 Effects of heat stress on leaf electrolyte leakage in two tall fescue genotypes at 0,2,6,14 and 21days after treatment

图5 高温胁迫0,6和21d对不同品系高羊茅根系活力的影响Fig.5 Effects of heat stress on turf quality in two tall fescue genotypes at 0,6and 21dafter treatment
热处理之前,热敏感型高羊茅叶片的相对电解质渗漏显著高于耐热型高羊茅叶片的相对电解质渗漏。随着高温胁迫的进行,虽然前两天热敏感高羊茅叶片的相对电解质渗漏相比热处理前没有显著性差异,随着胁迫时间的延长,热敏感高羊茅叶片的相对电解质渗漏加剧,到21d时,热敏感高羊茅叶片的相对电解质渗漏高达47.2%,显著高于热敏感高羊茅叶片的相对电解质渗漏(图4)。
2.3 高温对不同品系高羊茅根系活力的影响
高温导致高羊茅根系活力降低(图5)。热处理之前,耐热型高羊茅的根系活力高达607.6μg/(g·h),要显著高于热敏感型高羊茅的根系活力。随着高温胁迫的进行,热敏感型高羊茅的根系活力迅速下降,而耐热型高羊茅的根系活力前期没有显著性差异,直到21 d时才显著降低。因此,在高温处理21d时,热敏感型高羊茅的根系活力仅为261.5μg/(g·h),要显著低于耐热型高羊茅的根系活力。
2.4 高温对高羊茅初级代谢产物的影响
运用气质联用化学工作站(Agilent MSD Productivity Chemstation),通过比对出峰时间和荷质比分别检测鉴定出包括游离氨基酸、糖类、有机酸、糖苷和糖醇在内的25种代谢产物(表1)。将鉴定出的25种代谢产物进行Hierarchical clustering层次聚类分析(图6)发现,高温导致高羊茅体内代谢产物大量积累,其中氨基酸(如脯氨酸、缬氨酸、谷氨酸)和糖类(如葡萄糖、蔗糖)在两个品系高羊茅叶片中大量积累,而有机酸和脂肪酸类(如柠檬酸、棕榈酸)在高羊茅根系中大量积累。两个不同品系高羊茅叶片和根系中的25种代谢产物的变化情况,其中有7种代谢产物含量变化非常明显,这7种代谢产物将会在接下来结果和讨论部分重点分析(表2和表3)。

表1 高温胁迫0,2,6,14和21d高羊茅叶片和根系中鉴定的25种代谢产物Table 1 List of 25metabolites identified in leaves and roots of tall fescue exposed to heat stress at 0,2,6,14and 21days
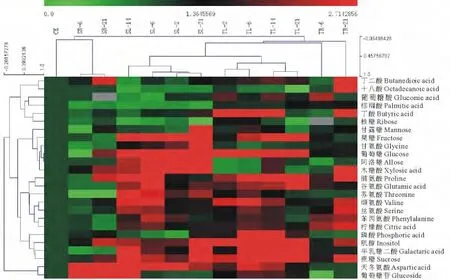
图6 不同品系高羊茅叶片和根系应答高温胁迫的代谢产物变化的层次聚类分析Fig.6 Hierarchical clustering analysis of metabolites in samples obtained from leaves and roots of tolerant tall fescue genotype(T)and sensitive tall fescue genotype(S)

表2 高温胁迫0,2,6,14和21d,不同品系高羊茅叶片中25种代谢产物变化情况Table 2 Change of 25metabolites identified in leaves of two tall fescue genotypes exposed to heat stress at 0,2,6,14and 21days
在轻度高温胁迫(6d)、中度高温胁迫(14d)和严重高温胁迫(21d)条件下,耐热型高羊茅PI 578718叶片和根系中柠檬酸的积累均显著高于其在热敏感型高羊茅PI 234881中的积累(图7);缬氨酸的积累在高羊茅2个基因型的叶片中先上升后下降,在根系中,缬氨酸的积累显著上升,严重高温胁迫(21d)条件下,耐热型高羊茅PI 578718中缬氨酸的积累要显著高于在热敏感型高羊茅PI 234881中的(图8);谷氨酸的积累在高温处理第6天的高羊茅2个基因型叶片中均出现显著增加,但在重度胁迫(21d)下,热敏感型高羊茅PI 234881叶片中谷氨酸含量下降,而耐热型高羊茅PI 578718叶片中谷氨酸积累上升,同时谷氨酸含量在高羊茅2个基因型根系中含量均上升,而且在耐热型高羊茅PI 578718中上升更明显(图8);脯氨酸含量在高温处理第2,6,14天的热敏感型高羊茅PI 234881中与高温处理第2,6,21天的耐热型高羊茅PI 578718中均显著升高(图8);蔗糖和肌醇的积累在高羊茅2个基因型叶片与根系中均显著升高,而且在长期高温胁迫条件下(21d),耐热型高羊茅PI 578718叶片和根系中的蔗糖和肌醇含量均要显著高于热敏感型高羊茅PI 234881中的(图9);棕榈酸在高羊茅2个基因型的叶片与根系中被检测到,与耐热型高羊茅PI 578718相比,高温处理前,热敏感型高羊茅PI 234881叶片和根系中含有更高水平的棕榈酸,重度高温胁迫下,热敏感型高羊茅PI 234881和片和根系中棕榈酸含量下降,而耐热型高羊茅PI 578718叶片和根系中棕榈酸含量上升(图10)。

表3 高温胁迫0,6和21d,不同品系高羊茅根系中25种代谢产物变化情况Table 3 Change of 25metabolites identified in roots of two tall fescue genotypes exposed to heat stress at 0,6and 21days

图7 高温胁迫对不同品系高羊茅叶片(0,2,6,14和21d)和根系(0,6和21d)中柠檬酸的影响Fig.7 Citric acid identified in leaves and roots of two tall fescue genotypes exposed to heat stress at 0,2,6,14,21days and 0,6,21days
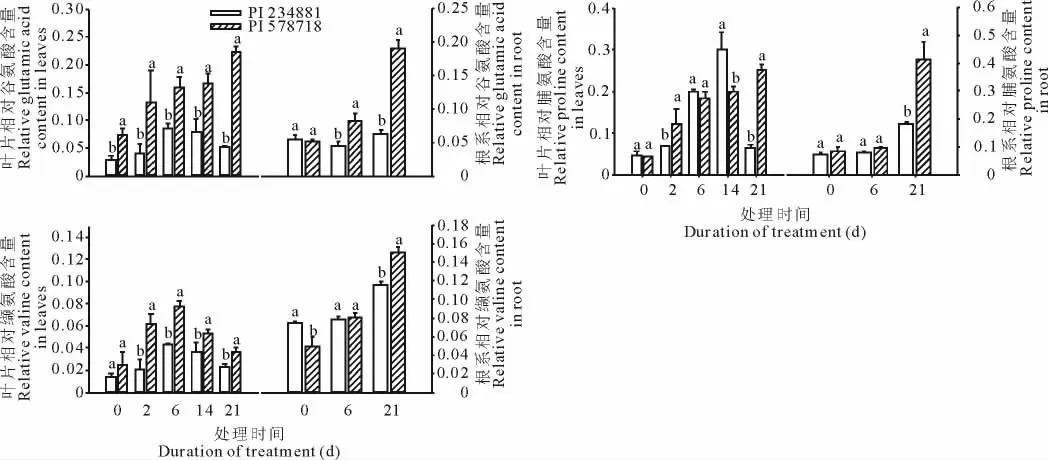
图8 高温胁迫对不同品系高羊茅叶片(0,2,6,14和21d)和根系(0,6和21d)中谷氨酸、缬氨酸和脯氨酸的影响Fig.8 Proline,valine and glutamic acid identified in leaves and roots of two tall fescue genotypes exposed to heat stress at 0,2,6,14,21days and 0,6,21days

图9 高温胁迫对不同品系高羊茅叶片(0,2,6,14和21d)和根系(0,6和21d)中蔗糖和肌醇的影响Fig.9 Sucrose and inositol identified in leaves and roots of two tall fescue genotypes exposed to heat stress at 0,2,6,14,21days and 0,6,21days

图10 高温胁迫对不同品系高羊茅叶片(0,2,6,14和21d)和根系(0,6和21d)中棕榈酸的影响Fig.10 Hexadecanoic acid identified in leaves and roots of two tall fescue genotypes exposed to heat stress at 0,2,6,14,21days and 0,6,21days
3 讨论
3.1 高温对高羊茅外观形态及生理的影响
草坪质量是草坪在其生长和使用期内叶片色泽、质地、均一性等表型的综合体现[20]。冷季型草坪草对高温敏感,在夏季草坪休眠或者死亡,草坪会发生枯黄,从而降低草坪的观赏性[21-22]。本试验发现高温降低了高羊茅的草坪质量,相比耐热性高羊茅,热敏感型高羊茅的草坪质量下降得更明显。
叶绿素反映叶片光合能力的强弱;植物叶片相对含水量的大小,可以部分反映植物抗逆性的能力;膜的功用保持不变是维持光合作用和呼吸作用的关键[15-16]。叶片相对电解质渗漏能够指示叶片膜结构破坏程度,叶片相对电解质渗漏的大小,可以作为判断植物耐热性水平的评价指标之一[23-24]。高温处理下,叶片电解质渗漏加剧,可见持续高温对高羊茅叶片膜结构造成了破坏,导致膜结构损伤,电解质渗漏加剧。热敏感型高羊茅植株失水严重,植物叶片含水量的变化一定程度上反映了植株的保水能力和抗逆能力,含水量变化越小,其保水能力和抗逆性就越强[16,24]。从实验结果可以看到,在高温胁迫下,热敏感型高羊茅品种叶绿素含量和叶片相对含水量显著下降,叶片电解质渗漏明显加剧均表明其对高温变化更加敏感。
根系活力反映根系吸收能力的强弱,根系活力强,根系吸水和吸收矿质营养的水平也高,植物抗逆能力也强[25]。本实验发现,高温胁迫导致高羊茅根系褐变,活力下降,相比耐热型高羊茅,热敏感高羊茅的根系活力下降得更明显。
高温胁迫虽然引起了高羊茅2个基因型草坪质量、叶片RWC、叶绿素含量以及根系活力的降低,叶片EL的升高,但耐热型高羊茅PI 578718较热敏感型高羊茅PI 234881能保持相对高水平的叶片水分状态、光合能力以及更稳定的膜系统,尤其在经过长时期高温处理以后,该研究结果与实验室进行的高羊茅耐高温筛选结果相一致,即在相同高温条件下,耐热型高羊茅PI 578718比热敏感型高羊茅PI 234881具有更强的耐热能力。而高温胁迫给高羊茅造成的伤害程度以及基因型间耐受高温胁迫的差异,可能与代谢产物的不同积累密切相关,下面将详细进行讨论。
3.2 高温对高羊茅代谢产物的影响
3.2.1 对有机酸的影响 在高羊茅2个基因型的叶片或根系中,共检测出包括丁酸、丁二酸和柠檬酸等共5种有机酸,其中柠檬酸是参与呼吸作用过程中三羧酸循环的中间产物。本试验中,高温胁迫下,高羊茅叶片和根系中柠檬酸含量显著上升,而且耐热型高羊茅叶片和根系中的柠檬酸含量均显著高于其在热敏感型高羊茅中的(图7),而且在严重高温胁迫(21d)下,耐热型高羊茅叶片和根系中柠檬酸含量分别是热敏感高羊茅的1.8和3.2倍。研究表明,柠檬酸在植物体内是通过丙酮酸脱羧后形成的乙酰辅酶A与草酰乙酸缩合形成6C的柠檬酸[26]。而且以往的研究发现,柠檬酸合成酶受高温的影响,植物可以通过缓解柠檬酸合成酶的热变性,并促进热变性的柠檬酸合成酶复性来抵御高温胁迫,高温下植物通过对柠檬酸合成酶的有效保护,不但可以增加线粒体的耐热性,而且可以提高植物的抗热能力[27]。但是目前,有关柠檬酸与耐热性之间的联系尚未明确。
3.2.2 对氨基酸的影响 氨基酸在植物生长代谢过程中具有十分重要的作用。氨基酸不仅能够合成蛋白质,而且还是许多植物应答非生物胁迫相关代谢产物的前体[28]。本研究中,缬氨酸含量在高羊茅2个基因型的叶片中先上升后下降,在根系中,其含量显著上升,而且耐热型高羊茅PI 578718中缬氨酸含量要显著高于在热敏感型高羊茅PI 234881中的(图8)。缬氨酸受胁迫后表达量的升高在许多逆境条件下均有报道[10,29]。在本研究中,缬氨酸含量在耐热型高羊茅PI 578718根系中的升高,可能作为初生代谢物而产生更多的次生代谢物,以保护植物组织免受由高温胁迫所引发的伤害。
通常非生物胁迫会导致细胞内蛋白质的降解,促使中间铵的积累[30]。据相关研究报道,不同形式或者不同亚型的谷氨酸,常作为逆境响应物质在受到不同胁迫环境的植物体中积累[31]。在本研究中,谷氨酸含量在高羊茅2个基因型根系中含量均明显增加,尤其是在耐热型高羊茅体内(图8),以往的研究表明,谷氨酸作为代谢途径中重要的中间物,在植物代谢过程起到了非常重要的作用[31],这也说明谷氨酸含量的维持和积累有助于高羊茅更好地适应和耐受高温胁迫。
与氨基酸不同,脯氨酸则是一种非蛋白且与胁迫相关的氨基酸[32]。脯氨酸作为能量库、亚细胞结构稳定剂、渗透调节剂、自由基清除剂以及胁迫信号分子,在植物的防御系统中扮演了诸多角色[33]。在高温胁迫的初期(6 d),脯氨酸在热敏感型高羊茅PI 234881叶片中的迅速积累,表明在轻度高温胁迫下脯氨酸可能作为一种胁迫信号分子而起作用。在高温处理第14天时,热敏感型高羊茅PI 234881叶片中的脯氨酸含量显著高于耐热型高羊茅PI 578718中的,而在高温处理第21天时,耐热型高羊茅PI 578718叶片中的脯氨酸含量显著高于热敏感型高羊茅PI 234881中的,这表明脯氨酸在重度高温条件下的积累,可能是高羊茅(尤其是耐热基因型)适应高温胁迫的一种响应。
3.2.3 对糖与糖醇的影响 在植物体中,逆境环境下经常会诱导各种糖类和糖醇等相容性物质产生积累[32]。这些低分子量的有机化合物,在环境刺激下出现积累,具有渗透调节、细胞功能保护、细胞组分完整性的维持等功能[10,33]。高温胁迫初期,植物叶片的相对含水量仍然较高,一些糖类和糖醇类物质的积累有助于细胞维持渗透势,以限制或减少植物水分的散失[34-35]。许多糖类,如果糖、半乳糖、葡萄糖、核糖以及蔗糖,均可作为高温条件下起渗透调节作用[10,36]。尤其是蔗糖,已证明在剧烈的脱水和热胁迫结合处理的拟南芥中,蔗糖将会取代脯氨酸作为主要的渗透保护剂[11]。除了作为渗透调节物质、胁迫诱导代谢产物改变的底物或者信号分子以外,蔗糖在一系列被测试的小分子糖中具有最强的抗氧化能力,同时也在植物呼吸代谢,能量供应过程中起到了非常重要的作用[37-39]。糖醇是水化的碳水化合物,通过似水的羟基基团,能够在大分子周围形成水化层[40]。糖醇不仅与碳的转运与贮存、超低温保护、硼的运输与能量传递紧密相关;更重要的是,它还参与逆境条件下植物渗透调节与活性氧的抑制[41]。肌醇是棉籽糖与水苏糖的前体,在植物中它们常作为渗透调节剂或者自由基的有效清除剂来提高植物的抗逆性[42]。在本研究中,蔗糖和肌醇的含量在高羊茅2个基因型叶片与根系中均显著升高,而且在长期高温胁迫条件下(21d),耐热型高羊茅PI 578718叶片和根系中的蔗糖和肌醇含量均要显著高于热敏感型高羊茅PI 234881中的(图9),耐热型高羊茅PI 578718可能正是通过借助蔗糖和肌醇更强的抗氧化能力和渗透保护能力来提高其耐热水平。
3.2.4 对脂肪酸的影响 研究表明,脂质的组成及饱和程度与植物对逆境胁迫的适应能力关系密切[43-44]。亚油酸,亚麻酸和棕榈酸是主要的质膜脂质成分。植物体内高水平的脂肪酸含量,尤其是不饱和脂肪酸的含量,能够维持逆境胁迫下植物细胞膜脂质双层更好的流动性[45-46]。在本研究中,高温处理前,相比耐热性高羊茅,热敏感型高羊茅PI 234881叶片和根系中含有更高水平的棕榈酸(图10)。高温胁迫下,热敏感型高羊茅PI 234881叶片和根系中棕榈酸含量下降,而耐热型高羊茅PI 578718叶片和根系中棕榈酸含量上升。这表明,高羊茅叶片和根系中棕榈酸含量上升有助于提高其耐热性,作为饱和脂肪酸,棕榈酸可能确保细胞膜质具有一定的流动性后,通过维持细胞膜更好的稳定性来提高高羊茅对高温的耐受能力。
总之,通过高羊茅2个基因型对高温胁迫的生理响应研究,发现在相同高温条件下耐热型高羊茅PI 578718比热敏感型高羊茅PI 234881更耐热。在受高温胁迫的高羊茅2个基因型叶片或者根系中,GC-MS分析鉴定出25种代谢产物,包括9种有机酸与脂肪酸、9种氨基酸、7种糖与糖醇。受高温胁迫的叶片中某些代谢产物表达量的升高或者维持,可能是耐热型高羊茅PI 578718与热敏感型高羊茅PI 234881之间存在耐热性差异的主要原因。在中度和/或重度高温胁迫下(14,21d),耐热型高羊茅PI 578718叶片中谷氨酸、脯氨酸、蔗糖、肌醇、棕榈酸等代谢产物的更多积累,使其比热敏感型高羊茅PI 234881更耐热。在重度高温胁迫下(21d),耐热型高羊茅PI 578718根系中缬氨酸、谷氨酸、脯氨酸、蔗糖、肌醇、棕榈酸等代谢产物更高水平的积累,使其比热敏感型高羊茅PI 234881具有更好地耐受高温胁迫的能力。而且本研究结果表明,柠檬酸作为抗氧化剂并参与呼吸作用三羧酸循环途径,它的高水平积累,可能更有助于提高高羊茅对高温胁迫的耐性和适应性。
Reference:
[1]Song M L,Wen X Z,Li Y L.Review on the effect of root rhizosphere on growth and metabolism in plants.Chinese Journal of Ecology,2010,29(11):2258-2264.
[2]Wang J Z,Cui L J,Wang Y,etal.Growth,lipid peroxidation and photosynthesis in two tall fescue cultivars differing in heat tolerance.Biologia Plantarum,2009,53:237-242.
[3]AL-Ghumaiz N S,Motawei M I.Productivity,forage quality and presence of dehydrin genes in some introduced pasture grass cultivars growing under heat stress in central region of Saudi Arabia.Australian Journal of Crop Science,2011,5:1001-1006.
[4]Yu J J,Du H M,Xu M,etal.Metabolic responses to heat stress under elevated atmospheric CO2concentration in a cool-season grass species.Journal of the American Society for Horticultural Science,2012,137:221-228.
[5]Jin B,Wang L,Wang J,etal.The effect of experimental warming on leaf functional traits,leaf structure and leaf biochemistry inArabidopsis thaliana.Biomed Central Plant Biology,2011,11:35-45.
[6]Yu J J,Chen L H,Xu M,etal.Effects of elevated CO2on physiological responses of tall fescue to elevated temperature,drought stress,and the combined stresses.Crop Science,2012,52:1848-1858.
[7]Nakagami H,Pitzschke A,Hirt H.Emerging MAP kinase pathways in plant stress signalling.Trends in Plant Science,2005,10:339-346.
[8]Cramer G R,Ergul A,Grimplet J,etal.Water and salinity stress in grapevines:early and late changes in transcript and metabolite profiles.Functional &Integrative Genomics,2007,7:111-134.
[9]Du H,Yu W,Liu Y,etal.Differential metabolic responses of perennial grass cynodon transvaalensis xCynodondactylon(C4)andPoaprotensis(C3)to heat stress.Physiologia Plantarum,2011,141:251-264.
[10]Kaplan F,Kopka J,Haskell D W,etal.Exploring the temperature-stress metabolome ofArabidopsis.Plant Physiology,2004,136:4159-4164.
[11]Rizhsky L,Liang H J,Shuman J,etal.When defense pathways collide.The response ofArabidopsisto a combination of drought and heat stress.Plant Physiology,2004,134:1683-1696.
[12]Wienkoop S,Morgenthal K,Wolschin F,etal.Integration of metabolomic and proteomic phenotypes.Molecular & Cellular Proteomics,2008,7:1725-1736.
[13]Du H M,Wang Z L,Huang B R.Differential responses of tall fescue and zoysiagrass to heat and drought stress.Proceedings of the Iind International Conference on Turfgrass Science and Management for Sports Fields,2008,8:207-213.
[14]Han C M,Zhang X Q,Peng Y,etal.Studies on the heat tolerance of 18Festucaacundinaceacultivars.Grassland and Turf,2006,114(1):35-44.
[15]Lu S Y,Guo Z F.Physiological responses of turfgrass to abiotic stresses.Acta Prataculturae Sinica,2003,12(4):7-13.
[16]Xu S,Li J L,Zhao D H.Research advances in physiological ecological and biochemical characteristics ofFestucaarundinacea.Acta Prataculturae Sinica,2004,13(1):58-64.
[17]Zhou Q.Laboratory Precedure for Plant Physiology[M].Beijing,China Agriculture Press,2000:11-12,62-63,159-160.
[18]Guo H J,Hu T,Fu J M.Effect of saline sodic stress on growth and physiological responses ofLoliumperenne.Acta Prataculturae Sinica,2012,21(1):118-125.
[19]Roessner U,Cornelia Wagner J K,Trethewey R N,etal.Simultaneous analysis of metabolites in potato tuber by gas chromatograsphy-mass spectrometry.Plant Journal,2000,23:131-142.
[20]Xu Y F,Jin J W,Sun X L,etal.A study on a model of turf quality evaluation based on reflectance spectroscopy.Acta Prataculturae Sinica,2009,18(4):256-259.
[21]He H R,Li Z D,Zhou E W,etal.Physiological responses to chilling of six warm-season turfgrasses.Acta Agrestia Sinica,2005,16(2):150-153.
[22]Zheng M Z,Liu J R,Qin W Z,etal.Effects of silicon on Kentucky bluegrass under drought stress.Pratacultural Science,2011,28(6):1009-1031.
[23]Zhao X,Li Y L.Variation of several physiological indices of five cool season turfgrasses under high temperature stress.Acta Prataculturae Sinica,2001,11(4):85-91.
[24]Zhao L L,Chen C,Ma L,etal.Physiological response and heat tolerance evaluation ofLotuscorniculatusunder high temperature stress.Pratacultural Science,2013,30(12):2018-2023.
[25]Zhang Y Q,Miao G Y.Effects of soil root-growing space on root physiological characteristics and grain yield of sorghum.Chinese Journal of Applied Ecology,2006,17(4):635-639.
[26]Arbona V,Manzi M,Ollas C,etal.Metabolomics as a tool to investigate abiotic stress tolerance in plants.International Journal of Molecular Sciences,2013,14:4885-4911.
[27]Charles G,Fatma K,Joachim K,etal.Metabolomics of temperature stress.Physiologia Plantarum,2008,132:220-235.
[28]Less H,Galili G.Principal transcriptional programs regulating plant amino acid metabolism inresponse to abiotic stresses.Plant Physiology,2008,147:316-330.
[29]OliverM J,Cuo L,Alexander D C,etal.A sister group contrast using untargeted global metabolomic analysis delineates the biochemical regulation underlying desiccation tolerance inSporobolusstapfianus.Plant Cell,2011,23:1231-1248.
[30]Gilbert G A,Gadush M Y,Wilson C,etal.Amino acid accumulation in sink and source tissues ofColeusblumeiBenth during salinity stress.Journal of Experimental Botany,1998,49:107-114.
[31]Semel Y,Schauer N,Roessner U,etal.Metabolite analysis for the comparison of irrigated and non-irrigated field grown tomato of varying genotype.Metabolomics,2007,3(3):289-295.
[32]Liu A R,Zhang Y B,Zhong Z H,etal.Effect of salt stress on the growth and osmotica accumulation ofColeusblumei.Acta Prataculturae Sinica,2013,22(2):211-218.
[33]Nanjo T,Kobayashi M,Yoshiba Y,etal.Biological functions of prolinein morphogenesis and osmotolerance revealed in antisense transgenicArabidopsisthaliana.The Plant Journal,1999,18:185-193.
[34]Bartels D,Sunkar R.Drought and salt tolerance in plants.Critical Reviews in Plant Sciences,2005,24:23-58.
[35]Martinelli T,Whittaker A,Bochicchio A,etal.Amino acid pattern and glutamate metabolism during dehydration stress in the resurrection plantSporobolusstapfianus:a comparison between desiccation-sensitive and desiccation-tolerant leaves.Journal of Experimental Botany,2007,58:3037-3046.
[36]Whittaker A,Martinelli T,Farrant J M,etal.Sucrose phosphate synthase activity and the co-ordination of carbon partitioning during sucrose and amino acid accumulation in desiccation-tolerant leaf material of the C4resurrection plantSporobolusstapfianusduring dehydration.Journal of Experimental Botany,2007,58:3775-3787.
[37]Vasquez-Robinet C,Mane S P,Ulanov A V,etal.Physiological and molecular adaptations to drought in andean potato genotypes.Journal of Experimental Botany,2008,59(8):2109-2123.
[38]Smirnoff N,Cumbes Q J.Hydroxyl radical scavenging activity of compatible solutes.Phytochemistry,1989,28:1057-1060.
[39]MorelliR,Russo-VolPe S,Bruno N,etal.Fenton dependent damage to carbohydrates:free radical scavenging activity of some simple sugars.Journal of Agriculture and Food Chemistry,2003,51:7418-7425.
[40]Nishizawa A,Ytlkinori Y,Shigeoka S.Galactinol and raffinose as a novel function to protect plants from oxidative damage.Plant Physiology,2008,147:1251-1263.
[41]Schobert B.Is there an osmotic regulatory mechanism in algae and higher plants.Journal of Theoretical Biology,1977,68:17-26.
[42]Brownp H,Hu H.Phloem mobility of boron is species dependent:evidence for phloem mobility in sorbitol-rich species.Annals of Botany,1996,77:497-505.
[43]Smirnoff N.Role of active oxygen in the response of plants to water deficit and desiccation.New Phytologist,1993,125:27-58.
[44]Navari-Izzo F,Quartacci M F,Melfi D,etal.Lipid composition of plasma membranes isolated from sunflower seedlings grown under waterstress.Physiologia Plantarum,1993,87:508-514.
[45]Navari-Izzo F,Ricci F,Vazzana C,etal.Unusual composition of thylakoid membranes of the resurrection plantBoeahygroscopica:changes in lipids upon dehydration and rehydration.Physiologia Plantarum,2006,94(1):135-142.
[46]Peng Y,Li Z.Effect of drought preconditioning on physiological responses to heat stress in two Kentucky bluegrass.Acta Prataculturae Sinica,2013,22(5):229-238.
[1]宋敏丽,温祥珍,李亚灵.根际高温对植物生长和代谢的影响综述.生态学杂志,2010,29(11):2258-2264.
[14]韩春梅,张新全,彭燕,等.18个高羊茅品种的耐热性比较研究.草原与草坪,2006,114(1):35-44.
[15]卢少云,郭振飞.草坪草逆境生理研究进展.草业学报,2003,12(4):7-13.
[16]徐胜,李建龙,赵德华.高羊茅的生理生态及其生化特性研究进展.草业学报,2004,13(1):58-64.
[17]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2000:11-12,62-63,159-160.
[18]郭慧娟,胡涛,傅金民.苏打碱胁迫对多年生黑麦草的生理影响.草业学报,2012,21(1):118-125.
[20]许岳飞,金晶炜,孙小玲,等.基于反射光谱技术评价草坪质量模型的研究.草业学报,2009,18(4):256-259.
[21]何会蓉,李志东,周而卫,等.几种暖季型草坪草对低温的生理反应.草地学报,2005,16(2):150-153.
[22]郑明珠,刘金荣,秦伟志,等.硅对干旱胁迫下草地早熟禾形态及生理特性的影响.草业科学,2011,28(6):1009-1031.
[23]赵昕,李玉霖.高温胁迫下冷地型草坪草几项生理指标的变化特征.草业学报,2001,11(4):85-91.
[24]赵丽丽,陈超,马林,等.高温胁迫下百脉根生理生化响应及耐热性评价.草业科学,2013,30(12):2018-2023.
[25]张永清,苗果园.根土空间对高粱根系生理特性及产量的影响.应用生态学报,2006,17(4):635-639.
[32]刘爱荣,张远兵,钟泽华,等.盐胁迫对彩叶草生长和渗透调节物质积累的影响.草业学报,2013,22(2):211-218.
[46]彭燕,李州.干旱预处理对抗寒性不同的2个草地早熟禾品种耐热性能的影响.草业学报,2013,22(5):229-238.
猜你喜欢
中老年保健(2022年3期)2022-11-21
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
农业与技术(2020年2期)2020-02-05
科学与财富(2019年15期)2019-10-21
小学生必读(低年级版)(2019年5期)2019-08-30
山西农经(2019年5期)2019-02-22
湖南林业科技(2017年1期)2017-02-06
孩子(2016年4期)2016-04-13
中国蜂业(2013年18期)2013-01-17
作物研究(2012年7期)2012-11-06
