
马尾松二代种子园无性系花量分析
2014-12-27 10:09:53唐效蓉伍新云曾令文周国瑾王晓锋
中南林业科技大学学报 2014年12期
唐效蓉 ,伍新云 ,曾令文 ,张 翼 ,周国瑾 ,唐 黎 ,王晓锋 ,杨 骏
(1.湖南省林业科学院,湖南 长沙 410004;2.桂阳县林业局,湖南 桂阳 424400;3. 城步苗族自治县云马国有林场,湖南 城步 422500;4.句容市林业科技推广中心,江苏 句容 212400)
马尾松二代种子园无性系花量分析
唐效蓉1,伍新云2,曾令文3,张 翼1,周国瑾3,唐 黎2,王晓锋4,杨 骏1
(1.湖南省林业科学院,湖南 长沙 410004;2.桂阳县林业局,湖南 桂阳 424400;3. 城步苗族自治县云马国有林场,湖南 城步 422500;4.句容市林业科技推广中心,江苏 句容 212400)
2012年于桂阳县苗圃马尾松二代种子园Ⅰ大区、Ⅴ大区以及城步县林科所马尾松二代种子园Ⅲ大区分别随机选取27、28、26个无性系,并于2012年和2013年对其胸径、冠幅以及全株雌、雄球花数量进行定株观测与分析。结果表明:不同地点、不同年度、不同无性系类型,其无性系的雌、雄球花花量平均值差异明显,平均值变幅及其差异较大;无性系间雌球花和雄球花花量主要表现为显著或极显著差异,年份间雌、雄球花花量主要表现为极显著差异;桂阳种子园雌球花的无性系×年份互作效应显著,其他互作效应不显著;地点间雌球花花量的差异均达极显著,雄球花花量的差异不显著;雌、雄球花花量无性系×地点的互作效应均不显著。雌球花和雄球花花量与冠幅、胸径主要表现为正相关,年度间和年度内雌球花花量与雄球花花量均为负相关;年度间雌球花花量与雌球花花量呈正相关,雄球花花量与雄球花花量呈正相关。不同无性系的雌、雄球花贡献率差异明显,有约10%无性系的雌、雄球花贡献率超过平均值的2~3倍,约20.00%无性系的雌、雄球花贡献率达50.00%左右。
马尾松;二代种子园;无性系;雌球花;雄球花;花量
建立林木种子园的目的在于持续、稳定、大量生产遗传品质优良的种子[1-2]。林木种子园无性系开花习性和花量是种子园建园无性系选择和配置的重要依据[3]。种子园的产量与其无性系的花期、花量以及建园地的气候条件密切相关[4]。马尾松种子园母树雌、雄球花的数量对种子园的产量有极大影响[5]。因此,研究马尾松种子园无性系雌、雄球花的数量及其变化规律,对提高其种子产量和品质有重要意义[2,6]。近年来,林木良种基地建设受到国家林业局种苗总站的高度重视。全国各地均建立了相应的国家级良种基地。国家财政给予了良种基地稳定的经费补贴。全国马尾松产区先后建立了较大面积的马尾松二代种子园。但现有马尾松种子园大多产量不高且不稳定[7],其原因是多方面的,其中病虫为害、花粉少、交配不充分、雌、雄花量少[7-9]等是主要原因。同时,二代种子园建园材料选择时,最初重点考虑的是其速生性,而对优树的开花结实能力关注较少[9]。这也对种子园的产量有较大影响。目前,对马尾松种子园开花习性及结实性能的研究报导较多[1-16],对马尾松种子园遗传多样性的研究也有一些报道[17-18],而针对马尾松二代种子园开花结实性能的研究报导较少。我们对湖南省桂阳县苗圃、城步林科所马尾松二代种子园无性系的开花习性和结实性能进行了研究,以期为二代马尾松种子园的建立和经营提供参考。
1 研究区概况
1.1 桂阳县苗圃马尾松二代种子园
桂阳县苗圃马尾松二代种子园位于东经112°38′52″~ 112°39′33″、 北 纬 25°44′50″~25°45′23″;属丘岗地貌类型,海拔228.5~357.8 m,多为坡度在20 °以下的阳坡或半阳坡;最低气温-9 ℃、最高气温41.3 ℃,年均气温17.7 ℃,年积温5 241 ℃,无霜期280 d,年降雨量1 473 mm,年日照时数1 644 h,相对湿度81%,4~6月为丰雨期,气候特征是四季分明、雨量充沛,日照充足,温和湿润[19];土壤为红壤,母岩以石灰岩为主,嵌有少量砂岩和页岩,土层厚度100 cm以上,肥力中等,pH值5.0~6.5。
1.2 城步县林科所马尾松二代种子园
城步县林科所马尾松二代种子园位于中亚热带南岭山脉,湘桂边境的越城岭与雪峰山交界处;地理坐标为东经 109°58′~ 110°37′、北纬 25°58′~26°42′,地处云贵高原东缘向湘中丘陵过渡地带;属低山地貌,海拔508.7~616.0 m;坡度15~30 °,坡向以南坡和北坡为主,地势开阔,阳光充足。该地属亚热带湿润气候区,为亚热带常绿阔叶混交林带,具有气候温和、热量充足、雨量充沛、四季分明、夏无酷暑、冬无严寒的特点;年平均气温16.1 ℃,1月平均气温4.7℃,7月平均气温26.6℃,极端最高气温38.6℃,极端最低气温-8.1℃,无霜期274 d,年平均日照时数1 487 h,年降水量1 221.4 mm,相对湿度76%~82%;成土母岩以板岩为主,间有少量页岩,土壤主要为山地黄壤,pH值5.5~6.0,肥力中等,土层厚度80~150 cm[20]。
2 研究方法
2.1 样株选择
2012年于桂阳种子园Ⅰ大区、Ⅴ大区以及城步种子园Ⅲ大区分别随机选取27、28、26个无性系(2013年桂阳种子园Ⅰ大区、Ⅴ大区仅调查24、21个无性系,2012年桂阳种子园Ⅴ大区与城步种子园Ⅲ大区相同无性9个,2013年桂阳种子园Ⅴ大区与城步种子园Ⅲ大区相同无性8个),其中桂阳种子园Ⅴ大区和城步种子园Ⅲ大区的无性系为湖南省及少量自然条件相近的、湖南省以北的周边地区引进的基因材料,于2005年春季嫁接;桂阳种子园Ⅰ大区的无性系为湖南省以南的周边地区引进的速生性好,特别是树高生长较突出的基因材料,于2005年冬季嫁接。每1个无性系选择6株生长势好、无病虫害、均匀分布于3个不同小区的母树作为定株观察样株。
2.2 观测时间
2个种子园均于2012年3~4月和2013年2~3月进行观测。
2.3 观测方法
观测样株的胸径、冠幅和全株的雌、雄球花数量。
2.4 数据处理
采用Excel 2003 和SPSS 17.0进行数据统计以及方差分析和相关分析。
3 结果与分析
3.1 不同无性系花量差异
不同无性系雌、雄球花的花量变化见图1~3,花量平均值及其变幅见表1。 由图1~3和表1可以看出:

图1 城步种子园Ⅲ大区2012年和2013年雌、雄花花量比较Fig.1 Comparison of numbers of male and female fl ower in region No. III of Seed Orchard of Chengbu County in 2012 and 2013

图2 桂阳种子园Ⅴ大区2012年和2013年雌、雄花花量比较Fig.2 Comparison of numbers of male and female fl ower in region No. V of Seed Orchard of Guiyang County in 2012 and 2013

图3 桂阳种子园Ⅰ大区2012年和2013年雌、雄花花量比较Fig.3 Comparison of numbers of male and female fl ower in region No. I of Seed Orchard of Guiyang County in 2012 and 2013
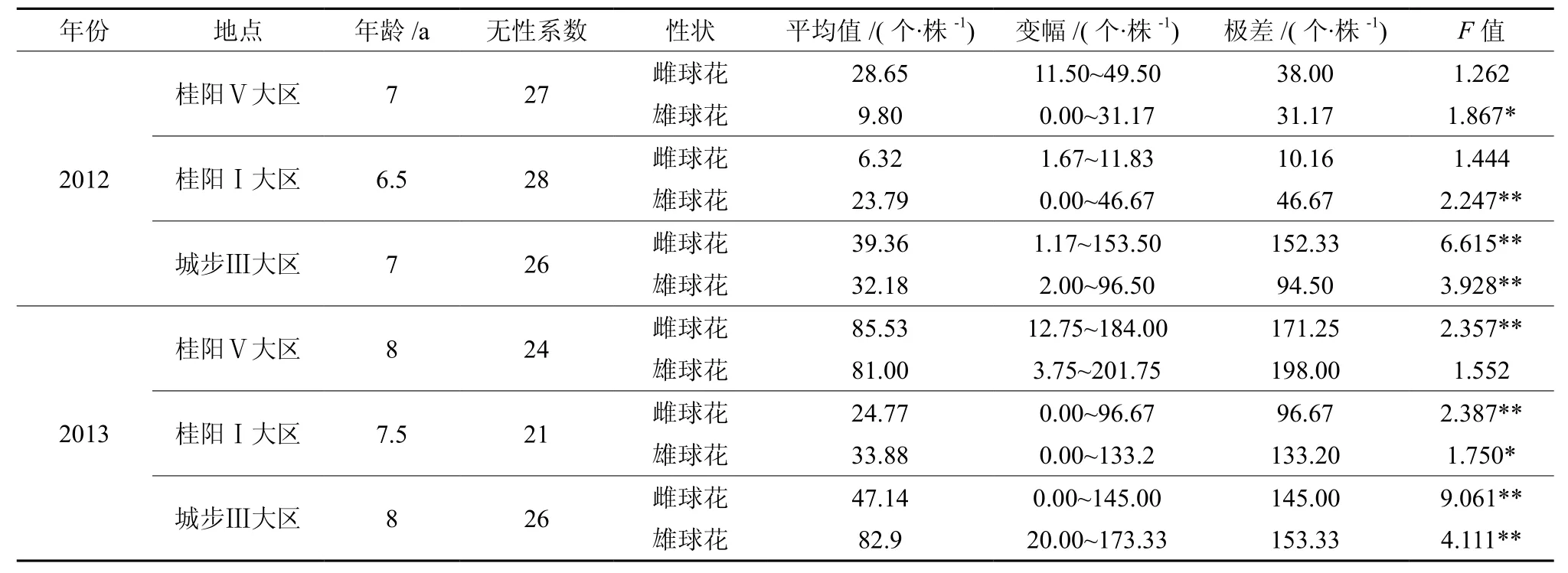
表1 无性系间雌、雄球花花量方差分析Table1 Variance analysis for numbers of male and female flower
(1)不同地点、不同年度、不同无性系类型,其无性系的雌、雄球花花量平均值差异明显。2个年度的雌球花花量以2013年桂阳Ⅴ大区的最大,达85.53 个·株-1;以2012年桂阳Ⅰ大区的最小,仅为6.32 个·株-1。雄球花花量以2013年城步Ⅲ大区的最大,达82.90 个·株-1;以2012年桂阳Ⅴ大区的最小,仅为9.80 个·株-1。2012年的雌、雄球花花量均以城步Ⅲ大区的最大,分别达39.36个·株-1和32.18 个·株-1。2013年的雌球花花量以桂阳Ⅴ大区的最大,雄球花花量以城步Ⅲ大区的最大。2012年2个种子园3个小区的无性系平均雌球花花量为24.78 个·株-1,平均雄球花花量为21.92个·株-1;2013平均雌球花花量为52.48 个·株-1,平均雄球花花量为65.93 个·株-1。2013年的雌、雄球花平均花量分别是2012年的2.12和3.01倍,年度间花量差异明显。相同来源的无性系,2012年城步Ⅲ大区的雌、雄球花平均花量均大于桂阳Ⅴ大区的,分别大10.71 个·株-1和22.38 个·株-1;2013年桂阳Ⅴ大区的雌球花平均花量则反而明显大于城步Ⅲ大区的(相差38.39 个·株-1),雄球花平均花量则相近(仅相差1.9 个·株-1)。桂阳种子园的2种类型的无性系,2个年度中Ⅴ大区的雌球花平均花量均明显大于Ⅰ大区的,2012年和2013年分别相差 22.33 个·株-1和 60.76 个·株-1;2012 年Ⅴ大区的雄球花平均花量明显小于Ⅰ大区的,相差13.99 个·株-1;2013年Ⅴ大区的雄球花平均花量则相反,明显大于Ⅰ大区的,相差47.12 个·株-1。
(2)不同地点、不同年度、不同无性系类型,其无性系的雌、雄球花花量变幅及其差异较大。2个年度的雌、雄球花花量变幅均以2013年桂阳Ⅴ大区的最大,分别达171.25 个·株-1和198.00个·株-1;以2012年桂阳Ⅰ大区的雌球花花量变幅最小,仅为10.16 个·株-1;以2012年桂阳Ⅴ大区的雄球花花量变幅最小,仅为31.17 个·株-1。2012年的雌、雄球花花量变幅均以城步Ⅲ大区的最大,分别达 153.33 个·株-1和 94.50 个·株-1。2013年的雌、雄球花花量变幅均以桂阳Ⅴ大区的最大。2012年2个种子园3个小区的无性系平均雌球花花量变幅为66.83 个·株-1,平均雄球花花量变幅为57.45 个·株-1;2013平均雌球花花量变幅为137.64个·株-1,平均雄球花花量变幅为161.51 个·株-1。2013年的雌、雄球花平均花量变幅分别是2012年的2.06和2.81倍,年度间花量变幅差异明显。相同来源的无性系,2012年城步Ⅲ大区的雌、雄球花平均花量变幅均大于桂阳Ⅴ大区的,分别相差114.33 个·株-1和 63.33 个·株-1;2013 年桂阳Ⅴ大区的雌、雄球花平均花量变幅反而均大于城步Ⅲ大区的,分别相差26.25 个·株-1和44.67 个·株-1。桂阳种子园的2种类型的无性系,2个年度中Ⅴ大区的雌球花平均花量变幅均明显大于Ⅰ大区的,2012年和2013年分别相差27.84 个·株-1和74.58个·株-1;2012年Ⅴ大区的雄球花平均花量变幅小于Ⅰ大区的,相差15.50 个·株-1;2013年Ⅴ大区的雄球花平均花量变幅则相反,明显大于Ⅰ大区的,相差 64.80 个·株-1。
3.2 无性系间花量差异显著性
无性系间方差分析结果见表1。由表1可以看出:2012年和2013年城步种子园不同无性系间的雌、雄球花花量以及2012年桂阳种子园Ⅰ大区的雄球花花量和2013年桂阳种子园2个大区的雌球花花量差异均达极显著;桂阳种子园2012年Ⅴ大区和2013年Ⅰ大区不同无性系间的雄球花花量差异均达显著;其他无显著差异。说明雌、雄球花的花量显著受无性系的遗传控制。
3.3 年份效应
不同地点年份间无性系的雌、雄球花花量方差分析结果见表2。由表2可以看出:城步种子园Ⅲ大区不同无性系间的雌、雄球花花量以及桂阳种子园Ⅴ大区的雌球花花量和Ⅰ大区的雄球花花量差异均达极显著,桂阳种子园Ⅴ大区的雄球花花量和Ⅰ大区的雌球花花量差异均达显著,其他差异不显著。城步种子园Ⅲ大区雄球花花量以及桂阳种子园Ⅴ大区的雌、雄球花和Ⅰ大区雌球花年份间的差异均达极显著,其他差异不显著。桂阳种子园2个大区的雌球花花量的无性系×年份互作效应显著,雄球花花量的无性系×年份互作效应不显著;城步种子园的无性系×年份的互作效应不显著。说明雌、雄球花花量除受遗传控制外,也与其生长历程相关[7]。从表1中雌、雄球花花量平均值也可以看出,嫁接后8 a的马尾松种子园较7 a的均大,特别是雄球花花量,增长显著。说明马尾松种子园早期雄花量不足,随着年龄的增长而增长显著。这也是影响马尾松种子园早期产量的因素之一[5]。
3.4 地点效应
不同年份地点间无性系的雌、雄球花花量方差分析结果见表3。由表3可以看出:2012年2个种子园中相同的9个无性系间雌球花和雄球花花量差异均不显著;2013年2个种子园中相同的8个无性系间雌球花花量差异达极显著,雄球花花量差异不显著。2012年和2013年2个年度的地点间雌球花花量的差异均达极显著,雄球花花量的差异不显著;雌、雄球花花量无性系×地点的互作效应均不显著。说明雌、雄球花花量除受遗传控制外,还与其生长环境相关[7]。

表2 不同地点年份间无性系的雌、雄球花花量方差分析Table2 Variance analysis for numbers of male and female flower inter-year in different places
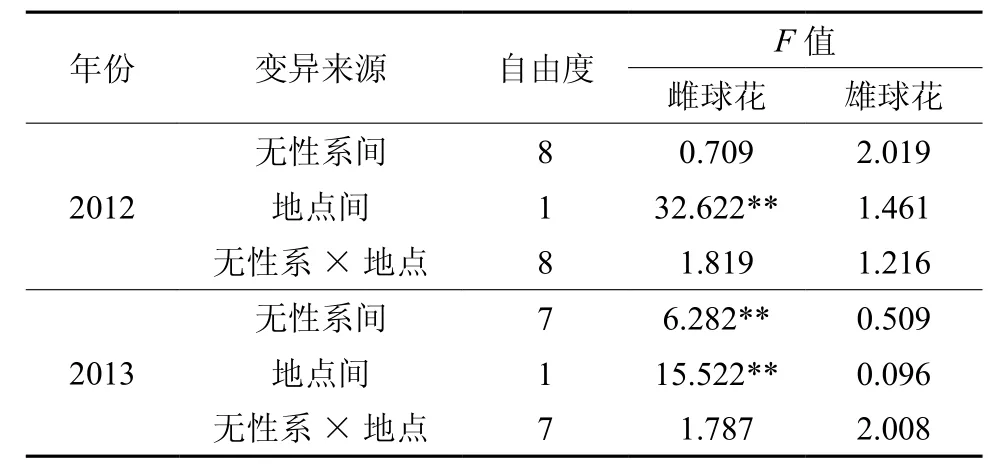
表3 不同年份地点间无性系雌、雄球花花量方差分析Table3 Variance analysis for numbers of male and female flower in one place in different year
3.5 花量与冠幅的相关性
雌、雄球花花量与冠幅的相关系数见4。由表4可以看出:雌球花和雄球花花量与冠幅主要表现为正相关,其中2013年桂阳种子园Ⅴ大区的雄球花花量与冠幅呈极显著正相关,2012年城步种子园Ⅲ大区和2012年桂阳种子园Ⅰ大区的雄球花花量与冠幅呈显著正相关;桂阳种子园Ⅰ大区的雌球花花量与冠幅呈弱度负相关。雌球花花量与雄球花花量均为负相关,其中桂阳种子园2012年Ⅰ大区的雌球花花量与雄球花花量呈极显著负相关,其他为弱度负相关。
3.6 花量与胸径及雌、雄球花间花量的相关性
雌、雄球花花量与胸径的相关系数见5。由表5可以看出:雌球花和雄球花花量与胸径主要表现为正相关,其中2012年桂阳种子园Ⅰ大区的雄球花花量与胸径呈极显著正相关,2013年桂阳种子园Ⅴ大区的雄球花花量与胸径呈显著正相关;桂阳种子园Ⅰ大区的雌球花花量与胸径呈负相关,其中2012年的为显著负相关。雌球花花量与雄球花花量均为负相关,其中桂阳种子园2012年Ⅰ大区的雌球花花量与雄球花花量呈极显著负相关,其他为弱度负相关。

表4 雌、雄球花花量与冠幅的相关系数Table4 Correlation coefficients of numbers of male and female flower with crown

表5 雌、雄球花花量与胸径的相关系数Table5 Correlation coefficients of numbers of male and female flower with diameter
3.7 花量的年度间相关性
雌、雄球花花量年度间相关系数见表6。由表6可以看出:2个种子园2012年的雌球花花量与2012年的雄球花花量均呈负相关,其中桂阳种子园Ⅰ大区的达极显著负相关;2013年的雌球花花量与2012年的雌球花花量均呈正相关,其中城步种子园Ⅲ大区的达极显著正相关;2013年的雌球花花量与2012年和2013年的雄球花花量均呈负相关。2013年的雄球花花量与2012年的雄球花花量均呈正相关,其中城步种子园Ⅲ大区和桂阳种子园Ⅰ大区的均呈极显著正相关;2013年的雄球花花量与2012年的雌球花花量主要表现为负相关,其中桂阳种子园Ⅰ大区的为极显著负相关,但桂阳种子园Ⅴ大区的为弱度正相关。

表6 雌、雄球花花量年度间相关系数Table6 Correlation coefficients of numbers of male and female flower with different year
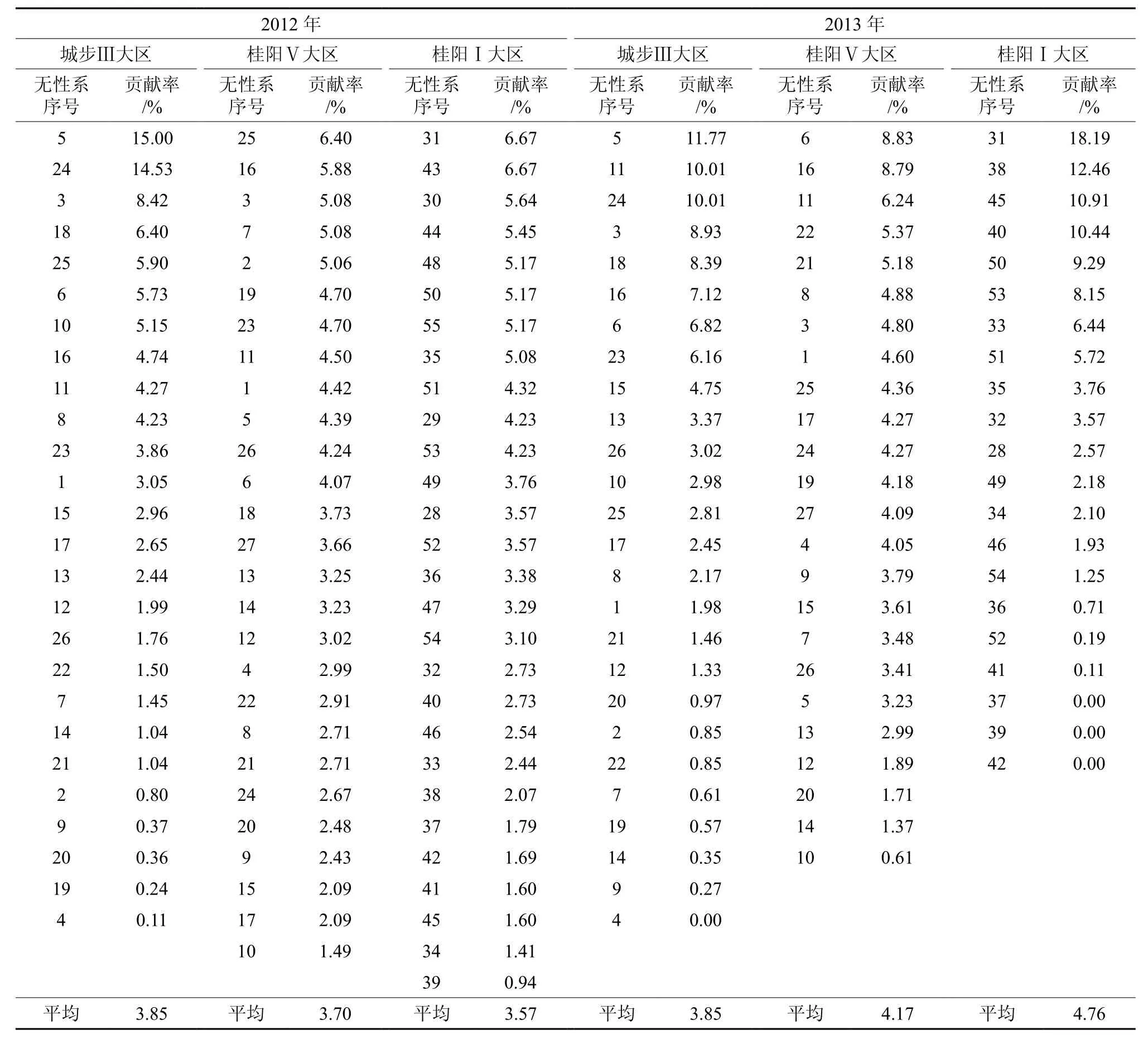
表7 无性系的雌球花贡献率排序Table7 Sequencing of contribution rate of male flowers of clones
3.8 雌、雄球花的贡献率
由表7、表8可以看出,不同无性系的雌球花贡献率差异明显,2个种子园的2个年度中均至少有1个年度有全雌花(无雄花)型或全雄花(无雌花)型无性系。2012年桂阳Ⅴ大区的27个无性系中有7个无性为全雌花、无雄花的无性系,2012年桂阳Ⅰ大区的28个无性系中有1个无性为全雌花、无雄花的无性系,2013年桂阳Ⅰ大区的21个无性系中3个无性为全雌花、无雄花的无性系,3个无性为全雄花、无雌花的无性系,2013年城步Ⅲ大区的26个无性系中有1个无性为全雄花、无雌花的无性系。
3.8.1 雌球花的贡献率
无性系的雌球花贡献率排序结果见表7。由表7可以看出:①2012年城步种子园Ⅲ大区有3个无性系的雌球花贡献率超过26个无性系贡献率平均值2倍,有2个无性系的雌球花贡献率超过3倍。②2013年城步种子园Ⅲ大区有4个无性系的雌球花贡献率超过26个无性系贡献率平均值的2倍,有1个无性系的雌球花贡献率超过3倍。桂阳种子园Ⅴ大区有2个无性系的雌球花贡献率超过24个无性系贡献率平均值的2倍;Ⅰ大区4个无性系的雌球花贡献率超过21个无性系贡献率平均值的2倍,有1个无性系的雌球花贡献率超过3倍。③城步Ⅲ种子园大区2012年和2013年平均19.23%无性系的雌球花贡献率达49.68%,2013年桂阳种子园Ⅰ大区20.00%无性系的雌球花贡献率达52.00%。
3.8.2 雄球花的贡献率
无性系的雄球花贡献率排序结果见表8。由表8可以看出:① 2012年城步种子园Ⅲ大区有3个无性系的雄球花贡献率超过26个无性系贡献率平均值2倍;桂阳种子园Ⅴ大区有4个无性系的雄球花贡献率超过27个无性系贡献率平均值的2倍,有2个无性系的雄球花贡献率超过3倍。②2013年城步Ⅲ大区有1个无性系的雄球花贡献率超过2倍;桂阳种子园Ⅴ大区有3个无性系的雄球花贡献率超过24个无性系贡献率平均值2倍;桂阳种子园Ⅰ大区有5个无性系的雄球花贡献率超过21个无性系贡献率平均值的2倍,有1个无性系的雄球花贡献率超过3倍。③桂阳种子园Ⅴ大区2012年和2013年平均19.61%无性系的雄球花贡献率达45.41%,2013年桂阳种子园Ⅰ大区20.00%无性系的雄球花贡献率达53.78%。
综合以上结果,除2012年桂阳种子园受虫害影响规律性不明显外,其他有约10%无性系的雌、雄球花贡献率超过平均值的2~3倍,约20.00%无性系的雌、雄球花贡献率达50.00%左右。表明马尾松种子园的雌、雄球花花量较强地受到部分无性系的遗传控制。

表8 无性系的雄球花贡献率排序Table8 Sequencing of contribution rate of female flowers of clones
4 结论与讨论
不同地点、不同年度、不同无性系类型,其无性系的雌、雄球花花量平均值差异明显,平均值变幅及其差异较大,其规律同徐进[5]、方彦[7]的研究结果。
2012年和2013年2个种子园,除2012年桂阳种子园的雌球花花量和2013年Ⅴ大区的雄球花花量无显著差异外,其他的雌球花和雄球花花量的差异均达到显著或极显著。桂阳种子园Ⅴ大区的雌、雄球花和Ⅰ大区的雌球花以及城步种子园Ⅲ大区的雄球花花量年份间的差异均达极显著,其他差异不显著;桂阳种子园2个大区的雌球花花量的无性系×年份互作效应显著,其他互作效应不显著。这可能是由于2012年松梢螟的危害在桂阳种子园相对较重,经综合防治,至2013年已基本得到控制,影响了调查结果。松梢螟的危害主要是从当年新梢中部蛀入髓心,致使新梢死亡。被害后的新梢基部往往萌生许多小芽,小芽长成重生状细枝,导致翌年无结果枝。
2012年和2013年2个年度的地点间雌球花花量的差异均达极显著,雄球花花量的差异不显著;雌、雄球花花量无性系×地点的互作效应均不显著。
雌球花和雄球花花量与冠幅、胸径主要表现为正相关,雌球花花量与雄球花花量均为负相关,但桂阳种子园Ⅰ大区的雌球花花量与冠幅、胸径均呈负相关。可能因为该小区的无性为引进行基因材料,其高生长的速生性和对环境的适应性影响了其结实性。这些还有待进一步研究。
2个种子园2012年的雌球花花量与2012年的雄球花花量均呈负相关;2013年的雌球花花量与2012年的雌球花花量均呈正相关,与2012年和2013年的雄球花花量均呈负相关;2013年的雄球花花量与2012年和2013年的雄球花花量均呈正相关,与2012年的雌球花花量主要表现为负相关,但桂阳种子园Ⅴ大区的为弱度正相关。这也可能与该小区的无性基因材料来源有关。
不同无性系的雌、雄球花贡献率差异明显,2个种子园的2个年度中均至少有1个年度有全雌花(无雄花)型或全雄花(无雌花)型无性系,其规律同赖焕林[3]的研究结果;除2012年桂阳种子园受虫害影响规律性不明显外,其他有约10%无性系的雌、雄球花贡献率超过平均值的2~3倍,有约10%无性系的雌、雄球花贡献率超过平均值的2~3倍,约20.00%无性系的雌、雄球花贡献率达50.00%左右。雌、雄球花贡献率大的无性系在种子园子代遗传物质组成中必然将占有较高的份额[7],进而影响种子园的遗传多样性,并对种子园高产稳产起着主导作用。
[1] 金国庆,秦国峰,周志春,等. 马尾松无性系种子园球果产量的遗传变异[J]. 林业科学研究,1998,11(3):277-284.
[2] 黄启明,廖 明. 马尾松种子园无性系开花习性的研究[J]. 贵州林业科技,1989,(4):21-28.
[3] 赖焕林,王章荣. 马尾松无性系种子园花期花量分析[J]. 浙江林学院学报,1996,13(4):405-410.
[4] 王以珊,罗 敏,曾令海,等. 气候条件对马尾松种子园产量的影响[J]. 广东林业科技,2002,18(3):10-15.
[5] 徐 进,洪永辉,朱金秋. 马尾松种子园的花量分析[J]. 江苏林业科技,1998,25(2):7-11.
[6] 陈天华,王章荣,李江蔺. 马尾松种子园无性系花期观察与分析 [D]. 马尾松种子园建立技术论文集. 北京:学术书刊出版社,1998. 126-135.
[7] 方 彦,张 薇,陈月旻,等. 马尾松实生种子园花量分析[J].林业科技开发,2007,21(6):43-46.
[8] 陈力友. 马尾松无性系种子园球花量变化研究[J]. 福建林业科技,2001,28(2):50-53.
[9] 邱进清. 建立马尾松实生种子园途径探讨[J]. 南京林业大学学报,1998,22(3):99-102.
[10] 芦 笛. 中国对英国邱园的千年种子库计划的贡献[J]. 生物灾害科学, 2013, 36(1): 121-126.
[11] 陈永庆,赵世远. 马尾松种子园无性系开花习性的观察[J].四川林业科技,1993,14(2):58-63.
[12] 谭健晖. 马尾松种子园无性系开花习性研究[J]. 广西林业科学,2001,30(2):76-78.
[13] 孔凡斌. 马尾松开花结实特征及种子产量预测 [J]. 林业科学研究,2002,15(2):150-155.
[14] 罗 敏,王以珊,曾令海,等. 马尾松种子园无性系开花特性与产地的关系[J]. 广东林业科技,2002,18(2):10-14.
[15] 覃开展,罗筱娥. 马尾松种子园无性系生长结实规律研究[J].广西林业科学,2001,30(1):28-31.
[16] 高大雄,刘宗友,王希群,等. 太子山马尾松种子园营建技术研究[J]. 湖南林业科技,2003,(1)(总123):15-21.
[17] 张冬林,杨玉洁,杨模华,等. 湖南城步马尾松种子园遗传多样性的ISSR研究[J].中南林业科技大学学报,2010,30(12):6-10.
[18] 杨玉洁,张冬林,杨模华,等. 湖南桂阳马尾松种子园遗传多样性的ISSR分析[J].中南林业科技大学学报,2010,30(3):85-89.
[19] 唐效蓉,徐清乾,蒋秀文. 间伐对马尾松天然次生林林下植物多样性的影响[J].湖南林业科技,2012,39(1):5-9.
[20] 曾令文,陈雪姣,杨宗琦. 城步县马尾松良种基地建设现状与发展思路[J].湖南林业科技,2009,36(5):57-59.
Analysis on the numbers of fl ower of clones of Pinus massoniana in second generation seed orchard
TANG Xiao-rong1, WU Xin-yun2, ZENG Ling-wen3, ZHANG Yi1, ZHOU Guo-jin3, TANG Li2, WANG Xiao-feng4, YANG Jun1
(1. Hunan Academy of Forestry, Changsha 410004, Hunan, China; 2.Forestry Bureau of Guiyang County, Guiyang 424400, Hunan,China; 3.Yunma State-owned Forest Farm of Chengbu Miao Minority Autonomous County; Chengbu 422500, Hunan, China; 4.Jurong Forestry Science and Technology Promotion Cente, Jurong 212400, Jiangsu, China)
Twenty seven, twenty eight and twenty six clones of Pinus massoniana were randomly selected from the corresponding region I and region V in the Seed Orchard of Guiyang County and region III of the Seed Orchard of Chengbu County in 2012. The stub observations and analyses on the diameter, crown, numbers of male and female fl ower of designation plants were performed in 2012 and 2013. The results show that among the P. massoniana clones growing different sites, with different tree age and different clone species, there were signif i cant differences in average values of male and female fl ower numbers, also obvious differences in changing of average value; There were signif i cant or very signif i cant differences in numbers of male and female fl ower of different clones, and very signif i cant differences in that of different years; The interaction effects of clone-year of female fl ower in the Seed Orchard of Guiyang County were signif i cant, and no signif i cant differences in other interaction effects; There were very signif i cant differences in numbers of female flower in different place, and no significant differences in the amount of the male flower ball; There were no significant differences of numbers of male and female fl ower in the interaction effects of different clone-year; There were the positive correlation among the numbers of male and female fl owers to the crown and diameter, there was the negative correlation between numbers of male and female fl owers inter-year and within year; There were the positive correlation between male and male fl owers or between female and female fl owers inter-year; there were signif i cant differences in contribution rate of male and female fl owers of different clones, and about 10% of them surpassed the average contribution rate by 2 to 3 times, and 20% of they reached about 50.00%.
Pinus massoniana; second generation seed orchard; clones; female fl ower; male fl ower; numbers of fl ower
S791.248
A
1673-923X(2014)12-0016-08
2014-01-25
“十二五”国家科技支撑计划课题“南方针叶树种高世代育种技术研究与示范”(2012BAD01B02);湖南省林业科技支撑项目“湖南马尾松高世代良种选与优良种质资源收集”(2008hl-10)
唐效蓉(1964-),女,湖南邵东人,研究员,主要从事林木遗传育种与栽培技术研究
[本文编校:文凤鸣]
猜你喜欢
东坡赤壁诗词(2022年2期)2022-04-15 22:05:45
戏剧之家(2021年13期)2021-11-14 07:45:17
花卉(2021年21期)2021-11-11 02:57:46
下一代英才(2020年10期)2020-11-12 02:12:31
广西林业科学(2020年3期)2020-11-03 06:38:52
今古传奇·故事版(2020年1期)2020-08-25 01:50:29
散文诗(2017年22期)2017-06-09 07:55:21
小溪流(故事作文)(2017年4期)2017-05-09 23:38:56
安徽农业科学(2016年21期)2016-09-13 08:21:08
东北林业大学学报(2014年3期)2014-03-06 03:19:16
