
异色瓢虫越冬聚集行为对其能量代谢的影响
2014-12-16 01:57李晓莉许永玉
环境昆虫学报 2014年6期
赵 静,李晓莉,许永玉,王 甦
(1.潍坊科技学院,山东寿光 262700;2.山东农业大学植物保护学院,山东泰安 271018;3.北京市农林科学院植物保护环境保护研究所,北京 100089)
在很多种类的昆虫中,聚集行为的发生是一种很常见的现象。有文献报道的具有聚集行为的非社会性节肢动物达124个属(Wertheim et al.,2005),其中昆虫115 属,例如瓢虫和蝴蝶等以成虫越冬的昆虫在越冬期间会发生聚集行为。每年秋天成千上万的君主斑蝶Danaus plexippus 群集性迁飞至其固定地点越冬。越冬期间有的瓢虫呈小规模的聚集,如几只到几十只;而有些瓢虫聚集规模很大,如美国的会聚长足瓢虫Hippodamia convergens 最大的聚集群体可达几千万只。昆虫的聚集行为能够对群体中的许多动态过程产生影响,如增强抵御天敌的效果(Hatle & Salazar,2001)、减少水分损失(Yoder et al.,2002)、增强繁殖力(Wells et al.,1990)等。对于昆虫来说,这种行为所产生的群体效应(Group effect),具有极其重要的意义(Glass et al.,1998),是昆虫抵御逆境、提高生存能力的一种生态对策(Clark & Faeth,1997)。
异色瓢虫Harmonia axyridis(Pallas)属鞘翅目瓢虫科,对蚜虫、叶螨、介壳虫等重要害虫具有很强的捕食能力,是一种重要的生防天敌(Koch,2003;王甦等,2007)。自然界中,异色瓢虫以成虫滞育越冬,越冬期间聚集行为尤为显著,一般几百甚至上千头聚集在向阳山洞的枯枝落叶或者屋檐下越冬。越冬期间异色瓢虫为什么要大量聚集在一起?有什么适应意义?目前对这些问题尚没有满意的答案。本文以异色瓢虫不同聚集群体(1 头、2 头、5 头、10 头、20 头和50 头)为对象,研究越冬成虫聚集行为对呼吸速率(rate of respiration,Rr)及能量代谢速率(rate of metabolism,Rm)的影响;此外,采用“Y”形嗅觉仪测定了异色瓢虫越冬种群与实验种群成虫的趋性选择,初步探讨异色瓢虫越冬聚集的原因。
1 材料与方法
1.1 供试虫源
实验种群的饲养:异色瓢虫采集自山东省泰安市郊区桃园内(36°15' N,116°59'E),采回后挑选健康成虫引入到人工气候室内饲养。具体饲养流程如下:将成虫置入养虫笼(金属框架+尼龙网(100 目);(50cm×50cm×50cm;30 头/笼)内集中饲养,每笼内放置60-80 头成虫,提供有足量豆蚜Aphis curviness Koch 的蚕豆苗,且每48 h更换一次。待雌雄成虫交配产卵后,取出带有卵块的蚕豆叶片移入洁净的玻璃培养皿(D=9.0 cm)中,卵孵化后转移至另一养虫笼内继续饲养豆蚜至成虫,羽化成虫在养虫笼内单独饲养(25 头/笼)。饲养期间为降低幼虫自残行为的发生,每笼内饲养80-100 头幼虫,且每48 h 更换一次蚕豆苗以提供充足的蚜虫。人工气候室环境条件为温度T=25±1℃,湿度RH=70±5%,光周期L∶D=16 h∶8 h。室内异色瓢虫种群均饲养2 代以上,且随机选取羽化10 d 左右的成虫(♀∶♂=1∶1)进行试验。
自然种群的采集:异色瓢虫越冬成虫于2009年10月末(平均温度为15℃左右)在晴朗的天气10∶00-13∶00 采集自泰安市天外村山(36°12'N,117°06'E)的向阳屋檐下,将采集到的成虫置于玻璃瓶中加入枯枝落叶并用纱布封口保存在室外。
1.2 越冬成虫呼吸量测定
实验共设置Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ和Ⅵ6个聚集度(分别用1 头、2 头、5 头、10 头、20 头和50 头表示),每个聚集度各设5个重复。随机选用个体大小一致的健康越冬成虫(自然种群),且不重复使用。
根据沈卫德等(1989)方法并加以改进,采用红外线CO2分析仪(GXH-3051,北京均方理化科技研究所)测定呼吸量。分别称取待测群体成虫的体重,然后将其转移至分析仪的暗室(4 cm×3 cm×2 cm)中测定。间隔30 s 记录1 次数据,共记录10 min。在整个测定过程中,CO2分析仪置于光照培养箱中(T=15±1℃)。
呼吸速率和能量代谢的计算(苏延乐等,2007):选择稳定的数据段N=N2-N1,计算出所选时间段样品呼吸释放CO2的量V(μL)。
呼吸速率Rr 用单位时间内单位重量试虫释放CO2的体积数(μL/g/s)表示,Rr=V/T M,式中V 为试虫释放CO2总体积(μL),T 为时间(s),M 为试虫体重(g)。
新陈代谢速率Rm 用能量单位W/g 计量,昆虫的能量消耗与呼吸释放CO2的关系系数一般为0.0245 J/μL CO2(Schmidt-Nielsen,1980),又由1 W=1 J/s,因此新陈代谢速率的计算公式:Rm=0.0245V/T M=0.0245 Rr。
1.3 异色瓢虫越冬种群与实验种群的趋性选择
用“Y”形嗅觉仪测定异色瓢虫实验种群与越冬种群成虫对不同味源的选择趋性,“Y”形管的两臂管长21 cm,两臂夹角60,直管长15 cm,内直径4 cm,接口均为标准磨口。“Y”形嗅觉仪的直管是试验瓢虫的入口,“Y”形管两臂管分别与两个味源瓶相连,在味源瓶前连接活性炭过滤器(以净化空气)。然后将活性炭过滤器的进气口与大气采样仪连接,由大气采样仪控制进气的流量和速度。上述装置除了磨口直接连接外,其余均用Teflon 管连接。两个味源瓶中分别放入50 头异色瓢虫实验种群或者越冬种群或者空气。每臂气体流量用大气采样仪控制在200 mL/min。试验前先行通气10 min,然后将异色瓢虫逐头引入嗅觉仪的直管内,观察瓢虫在15 min 内的行为反应。瓢虫爬过气味源臂的1/2 处并持续停留5 min 以上记为有趋性;如果瓢虫在15 min 内仍未到达臂的1/2 处,记为无趋性反应。试验中供试瓢虫每次测定30 头,测定结束后将整个装置先用去污粉清洗,再用95%酒精清洗热风吹干,以消除原有的味源残留,同时将嗅觉仪的两个味源瓶左右调换,以消除瓢虫对方位和味源的习惯性趋向。每次重复都用不同的瓢虫个体,共重复5 次。
行为指标:聚集指数(Aggregation index,AI)根据发生反应的个体在不同气味区内的分布比例来说明试虫在气味区内的聚集程度(Ishii,1967)。AI=(RN-CN/TN)×100,RN 为反应区的虫数,CN 为对照区的虫数,TN 为测试用的总虫数。引诱率(Attractant percent,AP)根据被引诱到味源区内的个体数占总试虫的比率来反映味源物质对试虫的引诱效果(Carlson,1973)。
1.4 数据处理
利用统计软件SPSS17.0 for windows 对各实验内容所得数据进行统计分析,获得平均值及标准误。对不同处理间的差异进行单因素方差分析(One-Way ANOVA)和Duncan 氏多重比较(P<0.05)。
2 结果与分析
不同聚集度间异色瓢虫越冬成虫的呼吸速率Rr 和新陈代谢速率Rm 均存在显著性差异(F=32.034;df=5;P<0.01)。聚集度Ⅱ和Ⅲ、Ⅲ和Ⅳ、Ⅲ和Ⅴ、Ⅳ和Ⅴ的Rr 和Rm 值间没有显著差异,其他聚集度之间均有显著性差异。
异色瓢虫越冬成虫的Rr 值(R2=0.8096)与聚集度之间呈现著负相关。即随着聚集度的增加,越冬成虫的呼吸速率Rr 呈逐渐降低的趋势(图1)。Rm 值的分析结果同Rr(图2)。
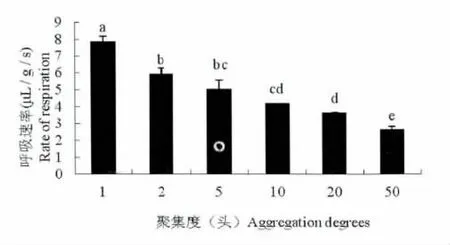
图1 异色瓢虫越冬成虫在不同聚集度下的呼吸速率Fig.1 The respiration rates of over-wintering adults in Harmonia axyridis in different group size
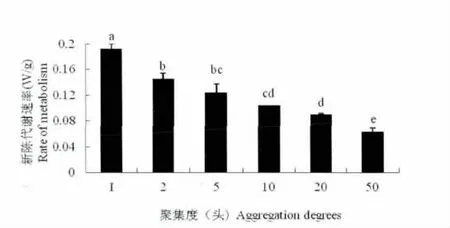
图2 异色瓢虫越冬成虫在不同聚集度下的能量代谢速率Fig.2 The metabolism rates of over-wintering adults in Harmonia axyridis in different group size
试验开始时异色瓢虫成虫绝大多数在“Y”型管内沿管壁环形前进,在其爬行过程中表现出对气味的探索行为。当进入某一味源所在的臂后,一部分便会停留,一部分会在两壁之间往返后作出选择,另有少数一直逗留在直管部分。由测定的行为指标可以看出(表1):异色瓢虫越冬种群与实验种群成虫均对各自的群体表现出趋性,但是当两种气味源分别为越冬种群和实验种群时,越冬成虫的这种趋性降低,而饲养成虫的趋性却无明显变化。

表1 异色瓢虫越冬种群和实验种群成虫的趋性选择Table 1 Tropism selection of overwintering population and experimental population of Harmonia axyridis adults
3 结论与讨论
异色瓢虫越冬期间聚集行为尤为显著,一般几百甚至上千头聚集在向阳山洞的枯枝落叶或者屋檐下越冬。本研究表明异色瓢虫越冬成虫的呼吸速率和新陈代谢速率随群体聚集度的增加而显著降低,这说明聚集行为会造成异色瓢虫呼吸量减少和生理活动减慢。这与Sakurai 等(1992)研究日本异色瓢虫越冬聚集现象时的结论相一致。聚集个体之间的相互抚慰作用使聚集个体较单独个体的活动量减少,进而降低个体的呼吸及新陈代谢速率(Tojy et al.,2005)。此外,聚集群体中特殊的微环境,例如低氧、高CO2等对呼吸的抑制作用也能导致能量代谢速率的降低。
聚集群体的大小对昆虫种群的很多生态过程都有显著影响(Costa et al.,2007)。本研究中不同聚集度下异色瓢虫越冬成虫的呼吸和新陈代谢速率存在显著差异也说明了这一问题。但是,昆虫的新陈代谢速率不会随着群体的逐渐增加一直减慢,其与昆虫群体大小间应该存在一个最适的临界点。当群体达到较大规模时,昆虫是否因为拥挤而活动量增加,进而导致新陈代谢速率的提高?另外,Clarke(2006)认为环境温度对生物体的新陈代谢影响显著,那么昆虫是如何在自然状态下调整其种群大小的?这些都是有待解决的问题。
瓢虫是如何聚集在一起的呢?有的认为是信息素的存在,或是先行者释放的气味。异色瓢虫的越冬聚集无方位性,Nalepa 等(2000)通过野外试验认为不存在挥发性的信息素,可能是基于接触交流信息或以往残留的气味来识别。本试验结果表明:异色瓢虫越冬种群与实验种群成虫均对各自的群体表现出趋性,但是当两种气味源分别为越冬种群和实验种群时,越冬成虫的这种趋性降低,而饲养成虫的趋性却无明显变化。这说明两种味源对异色瓢虫越冬成虫的趋性具有干扰作用,其越冬聚集行为的发生需要一种特殊的气味物质。异色瓢虫的越冬聚集的石洞内有一股特殊的气味,这是识别异色瓢虫越冬地的一个标志。进一步找到异色瓢虫越冬个体间通讯的物质基础和响应机制,这也是将要进行深入研究的。
聚集行为能够降低异色瓢虫越冬成虫的新陈代谢速率,减少能量消耗,提高存活率,这对其越冬来说具有非常重要的意义。研究昆虫的越冬聚集行为,找到个体间通讯的物质基础和响应机制,确定种群发生聚集现象的时间、地点、规模及其与各种环境因子的关系,监测聚集昆虫的行为及生理等,这可以为昆虫生物多样性保护、农林病虫害防治等提供理论基础和技术支持(苏岩乐等,2007)。如果一些害虫有这些习性,我们可利用来防治。北方的马铃薯瓢虫Henosepilachna vigintioctomaculata 有时会大量聚集在一起,找到其越冬地可集中进行防治。对于天敌瓢虫来说,应该好好保护这些越冬地点。但是,在美国很多地区,秋季异色瓢虫会选择民居或建筑物作为聚集地,而这种聚集行为会使人们产生厌恶感,人们会使用合成除虫菊酯和薄荷醇来驱赶异色瓢虫(Heulsman et al.,2002)。
References)
Carlson DA,Roan CS,Yost RA.Aggregation an mycophagous Drosophila:Candidate pheromone and field responses[J].Annals of the Entomological Society of America,1973,(67):696-704.
Clark BR,Faeth SH.The consequences of larval aggregation in the butterfly Chlosyne lacinia[J].Ecological Entomology,1997,(22):408-415.
Clarke A.Temperature and the metabolic theory of ecology[J].Functional Ecology,2006,(20)2:405-412.
Costa JF,Cosio W,Gianoli E.Group size in a gregarious tortoise beetle:patterns of oviposition vs.larval behavior[J].Entomologia Experimentalis et Applicata,2007,(125)2:165-169.
Glass E,Yoder J,Needham G.Clustering reduces water loss by adult American house dust mites,Dermatophagoides farinae(Acari:Pryoglyphidae)[J].Experimental and Applied Acarology,1998,(22):31-37.
Hatle JD,Salazar BA.Aposematic coloration of gregarious insects can delay predation by an ambush predator[J].Environmental Ecology,2001,(30):51-54.
Heulsman MF,Kovach J,Jasinski J.Multicolored Asian lady beetle Harmonia axyridis as a nuisance pest in household in Ohio.In:Jones GC,Zhai J,Robinson WH,Proceeding of 4thInternational Conference on Urban Pest[C].2002,243-250.Reno,Nevada.
Ishii S,Kuwahara Y.Aggregation pheromone of the German Cockroach Blattella germanica[J].Applied Entomology Zoology,1967,(2):203-218.
Koch RL.The multicolored Asian lady beetle,Harmonia axyridis:A review of its biology,uses in biological control,and non-target impacts[J].Journal of Insect Science,2003,(3):1-16.
Nalepa CA,Kidd KA,Hopkins DIJ.The multicolored Asian lady beetle(Coleoptera:Coccinellidae):orientation to aggregation sites[J].Journal of Entomology Science,2000,(35):150-157.
Sakurai H,Kawai T,Takeda S.Physiological changes related to diapause of the lady beetle,Harmonia axydis(Coleoptera:Coccinellidae)[J].Applied Entomology Zoology,1992,(27):479-487.
Schmidt-Nielsen K.Animal Physiology:Adaptation and Environment[M].2nded.Cambridge:Cambridge University Press,1980.
Tojo S,Nagase Y,Filippi L.Reduction of respiration rates by forming aggregations in diapausing adults of the shield bug,Parastrachia japonensis[J].Journal of Insect Physiology,2005,(51):1075-1082.
Wells H,Wells P,Cook P.The importance of overwinter aggregation for reproductive success monarch butterflies(Danaus plexippus)[J].Journal of Theoretical Biology,1990,(147):115-131.
Wertheim B,Baalen EA,Dicke M,et al.Pheromone-mediated aggregation in nonsocial arthropods:an evolutionary ecological perspective[J].Annual Review of Entomology,2005,(50):321-346.
Yoder JA,Hobbs HH,Hazelton MC.Aggregate protection against dehydration in adult females of the cave cricket,Hadenoecus cumberlandicus(Orthoptera,Rhaphidophoridae)[J].Journal of Cave and Karst Studies,2002,(64)2:140-144.
Shen WD,Hamano K,Mukaiyama F.Effect of environmental temperature on the body temperature and respiratory of silkworm[J].Acta Entomologica Sinica,1989,(12)1:12-16.[沈卫德,滨野國滕,向山文雄.环境温度对人工饲料育家蚕体温和呼吸量的影响[J].昆虫学报,1989,(12)1:12-16]
Su YL,Lv ZZ,Song Q,et al.Effects of overwintering aggregation behavior on energy metabolism in the firebug Pyrrhocoris apterus(Heteroptera:Pyrrhocoridae)[J].Acta Entomologica Sinica,2007,(50)12:1300-1303.[苏延乐,吕昭智,宋菁,等.始红蝽越冬聚集行为对其能量代谢的影响[J].昆虫学报,2007,(50)12:1300-1303]
Wang Su,Zhang RZ,Zhang F.Research progress on biology and ecology of Harmonia axyridis Pallas(Coleoptera:Coccinellidae)[J].Journal of Applied Ecology,2007,(18)9:2117-2126[王甦,张润志,张帆.异色瓢虫生物生态学研究进展[J].应用生态学报,2007,(18)9:2117-2126]
猜你喜欢
今日农业(2021年21期)2021-11-26
上海蔬菜(2020年6期)2020-12-24
河北果树(2020年4期)2020-11-26
红蜻蜓·低年级(2020年9期)2020-09-26
小学生作文(低年级适用)(2018年4期)2018-11-29
现代园艺(2017年11期)2017-06-28
作物研究(2017年3期)2017-05-11
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
