
松材线虫自然侵染后松树感病进程中光合作用及抗氧化酶活性的变化
2014-11-25 02:59黄秀凤徐华潮
环境昆虫学报 2014年2期
张 慧,黄秀凤,徐华潮
(1.临安市林业局,浙江临安 311300;2.富阳市林业局,浙江富阳 311400;3.浙江农林大学林业与生物技术学院,浙江临安 311300)
松材线虫Bursaphelenchus xylophilus 是国际上公认的重要检疫性有害生物。它能引起松材线虫病(Pine Wilt Disease 或称Pine Wood Nematode)(柴希民等,2003;杨宝君,2003),使感病植株迅速枯死,且传播蔓延极快,一旦感病将造成极为严重的损失,不论是幼龄小树,还是几十年生的参天大树都可被害致死,松树感病后一般1-3个月内即可全株枯死(秦复牛等,2003)。该病最早于1905年在日本九州岛的长崎发生。我国于1982年在南京中山陵的黑松Pinus thunbergii 上发现松材线虫,现已迅速蔓延。
尽管松材线虫病的发现已有近百年的历史(Yano,1913),但松材线虫的致病性却在1971年最先由清原和德重在接种试验中才得到证实(清原友也,1971),后续的研究进一步明确了松材线虫是引起松树死亡的病原。人们只是在近四十年内,才对松材线虫病进行了较为系统的研究,主要涉及不同寄主松树品种的抗病性、病原线虫的形态学和生物学、线虫的传播媒介、病害的发生和扩散规律以及防治和控制等(张星耀等,2003;李兰英等,2006;胡霞等,2007;刘洪剑等,2007;谢立群等,2007)。由于松材线虫病是以病原线虫为主导,综合有人为参与、媒介天牛传播、寄主松树以及环境因素互作的一种复杂病害系统,因此随着研究的深入,人们发现该病与通常认识的松材线虫有许多不同,尤其是其致病机理尚不明确,致使对该病的防治和控制工作进展缓慢(朱克恭,1992;赵博光等,2000)。
绿色植物光合作用是森林生态系统能量流动和物质循环的基础,也是生物固碳的最主要方式,因此,以光合作用为基础,研究植被生产力形成过程及其对未来气候变化的响应,一直是植物生理学及陆地碳循环研究领域的热点。目前,国内外对植物光合作用的研究大多集中在农作物、草木或树苗等易观测对象上(于强等,1999),而对高大乔木的研究则相对较少,尤其是在受病虫害危害后对植物光合生理的影响方面,鲜有文献报道。
部分学者探讨了感染松材线虫病后,松树光合作用与致病性的相互关系。Melakeberhan 等用松材线虫接种欧洲赤松幼苗,发现接种后24-36 h叶绿素荧光及针叶水势会降低;研究后指出,对光合作用的影响可能亦是导致松树死亡的重要因素(Melakebaran et al.,1990,1991)。但另有研究表明,在病害发展早期,当叶部水势高时松苗光合特性不受影响,到后期松苗光合作用降低是由于叶部水分亏缺引起的(Fukuda et al.,1992)。但对野外自然条件下感染松材线虫病后松树进行光合生理研究的报道尚未见到。本实验以黑松、马尾松为研究对象,研究探讨了松材线虫在自然侵染植株后,对寄主光合作用和针叶部分生理生化指标的影响,为进一步研究松材线虫的致病机理和早期诊断提供理论依据。
1 材料与方法
1.1 供试材料
试验样地分别选在浙江省平湖市平湖林场和余姚市牟山镇,两地均属亚热带湿润季风气候区,四季分明、气候温和、日照充足、雨量充沛。分别选取自然感病状态下的黑松和马尾松。根据树势生长状况、松针颜色及松脂流量变化,结合线虫分离,将不同受害程度的松树分为三个阶段,同时选择健康的一类(无线虫分离到)作为对照,具体标准如下:
健康期(HW):外观健康,松脂分泌正常,未能分离到松材线虫;
感病初期(IP):外观健康,松脂分泌明显减少,天牛取食点附近有少量松针褪色、萎蔫;
感病中期(MP):顶梢及其它嫩枝仅基部偶见黄色针叶,其余枝条上出现部分黄色针叶,其它针叶失去光泽;
感病重期(TP):顶梢及1 a 生嫩枝成灰绿色、萎蔫,部分针叶黄绿色,多年生枝条大多无绿色针叶。
1.2 采样方法
选取处于不同感病阶段的平湖黑松、余姚马尾松各3 棵,采用离体测定,每株分上中下东西南北共测定12 次,枝条采样后立即水培同时进行光合作用的测定;测定叶绿素含量和含水量指标的样品选取两种松树针叶部分,于低温保鲜箱中保存至室内测定;测量抗氧化酶活性的针叶样品则于液氮罐中冷冻保存至室内进行测定。
1.3 气体交换和叶绿素荧光测定
采用美国LI-COR 公司生产的便携式光合系统(LiCor-6400,USA)测定光响应曲线和荧光叶室(6400-09)测定叶绿素荧光参数,重复12 次;光响应曲线测定按Kitao 等(2003)的方法进行,光量子通量密度(PARI)设置为0,100,200,400,600,900,1500 (单位μmol/m2/s)。其他的叶绿素荧光参数按Genty 等(1989)的公式计算。因针叶狭小,无法填满叶室,故采用一束针叶平铺叶室,将整个叶室表面填满,每束平均18 根针叶。
1.4 生化分析
采用直接浸提法(李得孝等,2005)测定针叶色素含量;烘干法(郑炳松,2006)测定针叶含水量(鲜重的);滴定法(郑炳松,2006)测定过氧化氢酶活性;分光光度法(郑炳松,2006)测定超氧化物歧化酶活性;愈创木酚法(王学,2006)测定过氧化物酶活性。
1.5 数据处理
用Sigmaplot 软件作不同感病阶段净光合作用速率图和叶绿素荧光参数响应曲线;利用Photosyn Asisstant 软件计算光补偿点和光饱和点;用SPSS软件作差异分析。
2 结果与分析
结果显示,两种松树各个感病阶段净光合速率差异性显著。由图1 可看出,自然感病黑松和马尾松各感病阶段净光合作用速率(Pn)大小依次为,感病初期>健康阶段>感病中期>感病重期,即在感病初期Pn 最高,其中马尾松的Pn 上升更加明显,但到感病中期后Pn 迅速下降,马尾松光合速率数值在感病中期已为负值。黑松的气孔导度(Gs)随着感病程度的递增,出现明显的下降,马尾松的Gs 在感病初期略有上升,随后迅速下降。但二者的胞间CO2浓度(Ci)数值未出现明显下降,反而略有上升趋势。黑松、马尾松的蒸腾速率(Tr)随着病程的加重出现下降趋势,其变化趋势同Gs 非常相似。
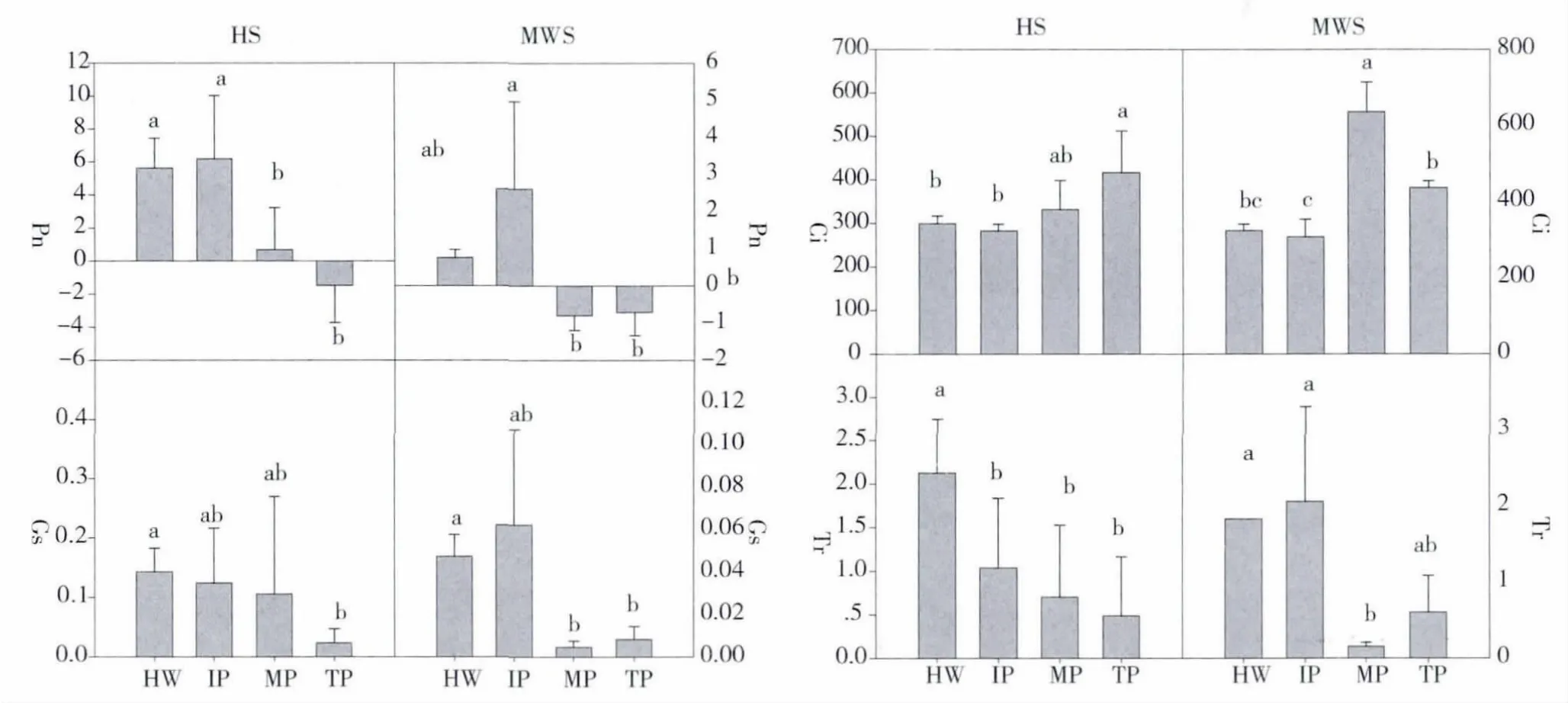
图1 黑松和马尾松各个感病阶段净光合作用速率变化(光强600μmol/m2/s)Fig.1 Lodgepole pine and Masson Pine all changes in the sense of disease stage of net photosynthesis rate(Light intensity 600μmol/m2/s)
由图2 可以看出,黑松和马尾松的ФPSⅡ在感病初期并没有下降,马尾松的ФPSⅡ反而略有上升,但随着感病程度的增加,两者的ФPSⅡ均迅速下降,在任何光强下,两者的ФPSⅡ均显著低于感病初期和健康木。Fv'/Fm'和qP 随着感病进程的增加,数值也不断下降,变化规律与ФPSⅡ基本相似。不同感病阶段的NPQ 数值也出现变化,黑松的NPQ 在感病初期明显上升,而感病中期只在低光强下高于健康木,在高光强下与健康木基本一致,在感病重期NPQ 则迅速下降。马尾松的NPQ 在感病初期与健康木基本一致,在感病中后期迅速下降。

图2 黑松和马尾松叶绿素荧光参数变化Fig.2 Changes of chlorophyll fluorescence parameters of lodgepole pine and Masson Pine
从表3 可见黑松和马尾松感病初期,针叶叶绿素a、b 和类胡萝卜素含量会先上升,随后会持续呈现下降的趋势。从图3 可以看出松材线虫侵染后,马尾松感病初期针叶含水量较健康阶段略有上升,随后逐渐下降,而黑松的针叶含水量则随感病程度持续下降,且下降速度略快于马尾松。黑松和马尾松受松材线虫感染后,导致了两种松树的抗氧化酶系统活性发生变化。实验结果表明,黑松和马尾松针叶CAT 活性出现持续下降,但马尾松下降速度更加迅速。两种松树的POD 活性则在感病初期略有升高,随后持续下降,两种松树的SOD 活性在感病初期也略有升高,但随后基本保持不变。
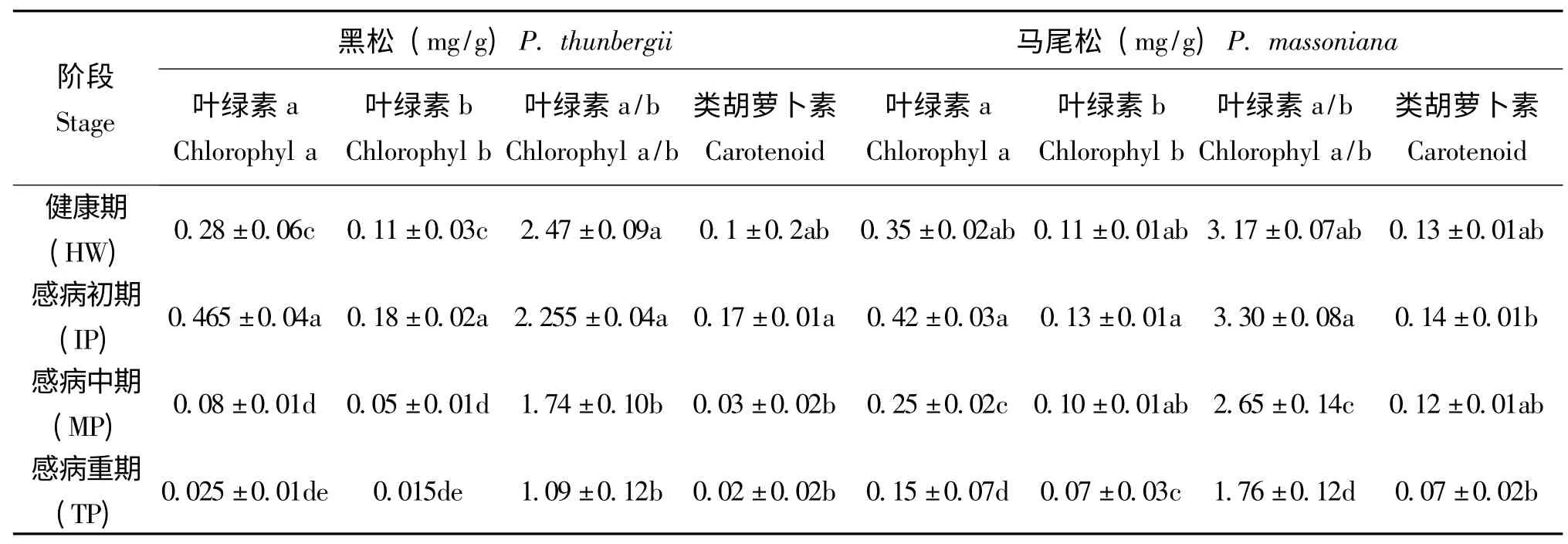
表3 松材线虫侵染马尾松、黑松后其针叶叶绿素和类胡萝卜素含量变化Table 3 Pine wood nematode infection of Pinus massoniana pine needles,the chlorophyll and carotenoid content change
4 结论与讨论
研究结果表明,松材线虫侵染黑松和马尾松后,在感病初期,马尾松的Pn 先出现上升现象,与健康阶段的差异性较大,为什么引起这一现象有待于进一步研究。由分析结果可看出,两种试验树随着病程的加重,两者的Pn 均明显下降。只是在感病中期马尾松的Pn 已出现负值,下降明显,说明马尾松对线虫病可能比黑松更敏感,而杨宝君等人(1987,1993)则表明马尾松抗性更强,这与本试验结果不一致,可能与试验地环境因子有关,具体原因还有待进一步研究。叶片含水量是引起Pn 下降的重要因素,有研究表明松材线虫侵入植株后会逐渐导致松树体内水分输导被中断造成的,进而导致水分亏缺 (谈家金等,2001),本研究中松树表现出萎蔫的症状,针叶含水量也下降,这与 Melakeberhan 和 Fukuda(Melakebaran et al.,1990;1991;Fukuda et al.,1992)研究结果相似,进一步证明了松树感染松材线虫后水分传导受阻的结论。由于含水量的下降引起了针叶Pn、蒸腾作用和Gs的下降,但胞间CO2浓度数值未下降,因此光合作用速率的下降与气孔因素无关。
PhiPS2 数值随着松材线虫的胁迫加深逐渐下降,表明电子传递速率下降,而ФPSⅡ则受Fv'/Fm'和qP 两者的影响。感病初期和健康阶段Fv'/Fm'无明显差异,随着线虫的大量繁殖出现持续下降,而qP 在感病中期下降明显,但随后基本不变,表明在感病中期电子传递的下降主要是Fv'/Fm'和qP 两者下降的结果,而在重期则是Fv'/Fm'下降的原因。叶绿素含量和类胡萝卜素含量随着病程的加重,也呈下降趋势,间接导致Pn 下降,但两者在感病初期均有所升高,这可能是植物的防御反应(邹春静等,2009)。
电子传递速率下降明显,还会引起多余电子,植物在吸收的光能超过所利用的能量时,会启动非辐射能量的耗散途径来耗散过剩的激发能以保护光合机构免受伤害,通过抗氧化酶系统清理过氧化氢酶能够将H2O2分解为H2O 和O2。过氧化物酶是植物体内清除H2O2的重要保护酶,广泛存在于植物的各个组织器官中,与植物抗性代谢关系很大。超氧化物歧化酶的活性与植物抗性密切相关,它能够防御活性氧或其他过氧化物自由基对细胞膜系统的伤害,普遍存在于植物的所有组织中。随着松材线虫侵染时间的延长,CAT 活性下降表明,受松材线虫感染松树体内清除氧自由基的能力下降,细胞膜损伤加重 (陈玉惠等,2002)。而受松材线虫感染后,POD 活性和SOD的活性升高则说明松树自身通过水水循环的作用发起了一定的防御措施,但这种抗逆性随着线虫的大量繁殖而减弱至消失。同时,随着非光化学淬灭系数(NPQ)数值的降低,寄主吸收的多余光能不能被耗散,形成自由基,进一步破坏抗氧化酶系统,引起Pn 下降。
References)
Chai XM,Jiang P.Occurrence and Control of Pine Wilt Disease [M].Beijing:China Agriculture.2003.[柴希民,蒋平.松材线虫病的发生与防治[M].北京:中国农业出版社,2003]
Chen YH,Ye JR,Wei CJ,et al.Effects of pine wood nematode infection on metabolism of active oxygen in Japanese Black Pine and Slash Pine Seedlings [J].Journal of Nanjing Forestry Unversity.2002,26 (4):21-22.[陈玉惠,叶建仁,魏初奖,等.松材线虫对黑松、湿地松幼苗活性氧代谢的影响[J].南京林业大学学报,2002,26 (4):21-22]
Fukuda K,Hogetsu T,Suzuki K.Photosynthesis and water status of pine-wood nematode infected pine seedings [J].J.Jpn.For.Soc.,1992,74 (1):1-8.
Fukuda K,Hogetsu T,Suzuki K.Cavitation and cytological changes in xylem of pine seedings inoculated with virulent and avirulent isolates of Bursaphelenchus xylophilus and B.mucronatus [J].Jan.For.Soc.,1992,74 (4):289-299.
Genty B,Briantais JM,Baker NR.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.Biochimica et Biophysica Acta,1989,990:87-92.
Hu Xia,Jiang XJ.Research progress on biological prevention technique of Bursaphelenchus xylophilus [J].Guangxi Forestry Science,2007,36 (4):199-201.[胡霞,蒋学建.松材线虫病生物防治研究进展.广西林业科学,2007,36 (4):199-201]
Kitao M,Utsugi H,Kuramoto S,et al.Light-dependent photosynthetic characteristics indicated by chlorophyll fluorescence in five mangrove species native to Pohnpei Island Micronesia[J].Physiologia Plantarum,2003,117:376-382.
Li LY,Gao L,Wen YL,et al.Advances in research on Bursaphelenchus xylophilus [J].Journal of Zhejiang Forestry Science and Technology,2006,26 (5):74-80.[李兰英,高岚,温亚利,等.松材线虫病研究进展[J].浙江林业科技,2006,26(5):74-80]
Li DX,Guo YX,Yuan HY,et al.Determined methods of chlorophyll from Maize [J].Chinese Agricultural Science Bulletin,2005,21(6):153-155.[李得孝,郭月霞,员海燕,等.玉米叶绿素含量测定方法研究[J].中国农学通报,2005,21 (6):153-155]
Liu HJ,Shu QL.Pine wilt disease by the media-monochamus alternatus advances in biological control [J].Anhui Agricultural Science Bulletin,2007,13 (8):127-128.[刘洪剑,束庆龙.松材线虫病的传播媒介-松墨天牛生物防治研究进展[J].安徽农学通报,2007,13 (8):127-128]
Melakeberhan H,Toivonen PMA,Vidaver WE,et al.Effect of Bursaphelenchus xylophilus on the water potential and water-splitting complex of photosystem Ⅱof Pinus sylvestris seedling [J].Physiological and Molecular Plant Pathology,1991,38 (2):83-91.
Melakebaran H,Webster JM.Effect of Btosaphelenchus xylophilus on the assimilation and translocation of C14 in Pinus sylvestris[J].Journal of Nematology,1990,22 (4):506-512.
Qin YYY,De CYS.Ma the ッstudents stumpage ni su ru,culture and inch filarial Bursaphelenchus sp full inoculation [J].Rilinzhi,1971,53:210-218.[清原友也,德重阳山.マッ生立木に文寸する丝虫Bursapherlenchus sp.の接种试验[J].日林志,1971,53:210-218]
Qin FN,Pan CS.Advances in research on Bursaphelenchus xylophilus[J].Journal of Anhui Agricultural University,2003,30 (4):370-376.[秦复牛,潘沧桑.松材线虫病研究进展.安徽农业大学学报,2003,30 (4):370-376]
Tan JJ,Fen ZX.Advance on physiopathology of Pine Wood Nematode Disease [J].Jiangxi Science,2001,19 (1):28.[谈家金,冯志新.松材线虫病病理生理学研究进展[J].江西科学,2001,19 (1):28]
Wang X.Plant Physiology and Biochemistry Experimental Principles and Techniques [M].Beijing:Higher Education Press,2006,l67-l68.[王学.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006,l67-l68]
Xie LQ,Ju YW,Zhao BG.Review on transmission mechanism of Bursaphelenchus xylophilus [J].Journal of Anhui Agricultural Sciences,2007,35 (19):5798-5800.[谢立群,巨云为,赵博光.松材线虫传播机理的研究进展[J].安徽农业科学,2007,35 (19):5798-5800]
Yang BJ.Pine Wilt Disease [M].Beijing:China Forestry.2003.[杨宝君.松材线虫病[M].北京:中国林业出版社.2003]
Yang BJ.The pesistance of pine sprcies to pine wood nematode,Bursaphelenchus xylophlus [J].Acta Phytopathologica Sinica,1987,17 (4):211-214.[杨宝君.不同松树品种对松材线虫的抗性[J].植物病理学报,1987,17 (4):211-214]
Yang BJ,Hu KJ,Wang QL.Studies on the resistance of pine trees to Pine Wood Nematode,Bursaphelenchus xylophilus [J].Forest Research,1993,6 (3):249-255.[杨宝君,胡凯基,王秋丽.松树对松材线虫的抗性研究[J].林业科学,1993,6(3):249-255]
Yano M.Investigation on the causes of pine mortality in Nagasaki Prefecture [J].Sanrin-Koho,1913,4 (supl.):1-14.
Yu Q,Xie XQ,Sun SF.Andances in simulation of plant photosynthetic productivity and canopy evapotranspiration [J].Acta Ecologica Sinica,1999,19 (5):744-753.[于强,谢贤群,孙淑芬,.植物光合生产力与冠层蒸散模拟研究进展[J].生态学报,1999,19 (5):744-753]
Zhang XY,Luo YQ.China's Major Forest Biological Disasters [M].Beijing:China Forestry.2003.1-29.[张星耀,骆有庆.中国森林重大生物灾害[M].北京:中国林业出版社.2003.1-29]
Zhu KG.Pine wilt disease infection source [J].Journal of Shandong Forestry Science and Technology,1992,4:45-48.[朱克恭.关于松材线虫病侵染源的研究[J].山东林业科技,1992,4:45-48]
Zhao BG,Guo DS,Gao R.A preliminary study on the relationship between the bacterium isolate B619 and Pine Wilt Disease [J].Journal of Nanjing Forestry University,2000,24 (4):72-74.[赵博光,郭道森,高蓉.细菌分离物B619 与松材线虫病关系的初步研究[J].南京林业大学学报,2000,24 (4):72-74]
Zheng BS.Modern Plant Physiology and Piochemistry Research Techniques [M].Zhejiang:Meteorological Press.2006,3-8.[郑炳松.现代植物生理生化研究技术[M].浙江:气象出版社.2006,3-8]
Zou CJ,Han SJ,Xu WD.Eco-physiological responses of Picea mongolica ecotypes to drought stress [J].Chinese Journal of Applied Ecology,2003,14 (9):1448-1449.[邹春静,韩士杰,徐文铎.沙地云杉生态型对干旱胁迫的生理生态响应[J].应用生态报,2003,14 (9):1448-1449]
猜你喜欢
美食(2022年12期)2022-12-09
浙江林业科技(2021年3期)2021-07-13
林业科学(2020年10期)2020-11-30
美食(2019年7期)2019-09-10
作文周刊·小学四年级版(2019年8期)2019-04-23
中国糖料(2018年4期)2018-07-31
长江大学学报(自科版)(2018年14期)2018-07-17
科学大众·小诺贝尔(2018年12期)2018-02-22
国际木业(2016年6期)2016-02-28
国际木业(2016年5期)2016-02-28
