
烟草类胡萝卜素异构酶基因的克隆及功能研究
2014-11-24 06:34:06史艳梅王燃杨军罗朝鹏李锋武明珠魏春阳林福呈屈凌波魏攀
中国烟草学报 2014年6期
史艳梅,王燃,杨军,罗朝鹏,李锋,武明珠,魏春阳,林福呈,屈凌波,魏攀
1 郑州大学化学与分子工程学院,河南省郑州市高新技术产业开发区科学大道100号 450001;
2 中国烟草总公司郑州烟草研究院,河南省郑州市高新技术产业开发区枫杨街2号 450001;
3 河南工业大学化学化工学院,郑州高新技术产业开发区莲花街100号 450001
生物技术
烟草类胡萝卜素异构酶基因的克隆及功能研究
史艳梅1,2,王燃2,杨军2,罗朝鹏2,李锋2,武明珠2,魏春阳2,林福呈2,屈凌波1,3,魏攀2
1 郑州大学化学与分子工程学院,河南省郑州市高新技术产业开发区科学大道100号 450001;
2 中国烟草总公司郑州烟草研究院,河南省郑州市高新技术产业开发区枫杨街2号 450001;
3 河南工业大学化学化工学院,郑州高新技术产业开发区莲花街100号 450001
为进一步研究烟草类胡萝卜素异构酶(Carotenoid isomerase,CRTISO)基因的功能,从普通烟草(Nicotiana tabacum)的cDNA中成功克隆出CRTISO基因编码区(CDS)全长,长度为1716 bp,Blast结果显示与马铃薯(Solanum tuberosum)和番茄(Solanum lycopersicum)的前番茄红素异构酶(Prolycopene isomerase)基因的相似度达93%。将CRTISO基因的部分序列插入烟草脆裂病毒(Tobacco rattle virus,TRV)载体中,构建重组载体pTRV2-CRTISO。通过TRV病毒诱导的基因沉默(Virus induced gene silencing,VIGS)系统抑制本氏烟草(Nicotiana benthamiana)CRTISO基因的表达,同时利用实时荧光定量PCR(Real-time PCR)检测CRTISO下游基因的表达量变化。结果表明,与对照组相比,pTRV-CRTISO载体侵染的烟草叶片出现黄色斑点;CRTISO基因的沉默导致下游番茄红素ε环化酶(ε-LCY)、番茄红素β环化酶(β-LCY)和胡萝卜素β-环羟化酶(β-OHase)基因表达量降低。对烟草CRTISO基因功能的研究有助于揭示烟草中类胡萝卜素合成代谢途径的调节机制,为选育优质烟草品种奠定理论基础。
普通烟草;类胡萝卜素异构酶;实时荧光定量PCR;病毒诱导的基因沉默
类胡萝卜素合成代谢途径在高等植物中扮演重要角色[1]。该途径的中间代谢产物,如β-胡萝卜素、黄体素等参与植物的光合作用[2];终产物包括脱落酸、独脚金内酯等,与植物抗氧化和生长发育相关[3]。烟草中的类胡萝卜素属于萜烯类化合物,其含量的高低能够影响烟叶的外观和内在品质,它不仅与调制后烟叶的颜色有关,而且其降解和热裂解产物包含近百种香气物质,与烟叶的香气有密切关系[4]。因此,烟草类胡萝卜素的研究在国内外烟草行业中始终是焦点,而烟草作为一种重要的模式植物,也为研究提供了良好的材料。1993年,Misawa等将细菌八氢番茄红素脱氢酶基因(crtⅠ)转入烟草,使其过量表达,结果转基因烟草中β-胡萝卜素的累积量明显提高[5]。2000年,Mann等将烟草中β-胡萝卜素酮酶基因(crt0)过表达,使虾青素等酮基类胡萝卜素在烟草中不断累积,最终使花蜜腺的颜色由黄变红[6]。
CRTISO基因是植物中类胡萝卜素合成的关键基因之一,它催化多反式番茄红素形成多顺式番茄红素,该基因表达量降低能够导致β-胡萝卜素大量积累[7]。然而烟草中关于该基因的研究甚少,我们只能从其它植物的CRTISO基因的研究中获取一些有价值的信息。Kato等发现柑橘中CRTISO基因下调能够使果实中β-胡萝卜素含量提高,证实了该基因与β-胡萝卜素的合成代谢相关[8]。陶能国等克隆了纽荷尔脐橙的CRTISO基因,该基因与拟南芥、番茄和柑橘的CRTISO基因高度相似,可以推测纽荷尔脐橙的CRTISO基因也与β-胡萝卜素的合成累积有密切关系[9]。
本研究从普通烟草中克隆CRTISO基因,并对该基因的表达和功能进行研究,旨在揭示该基因在烟草类胡萝卜素合成通路中的作用,为通过基因工程手段培育优良烟草品种奠定理论基础。
1 材料与方法
1.1 实验材料
普通烟草红花大金元和本氏烟草由郑州烟草研究院国家烟草基因研究中心于温室中栽培。普通烟草的培养条件为(27 ±1)℃,光照16 h,黑暗8 h,RH(60±2)%;本氏烟草的栽培条件为(23±1)℃,光照16 h,黑暗8 h,RH(60±2)%。
RNA反转录试剂盒、质粒提取试剂盒、限制性内切酶、LA Taq酶、DH5α、pMDR19-T Simple Vector、DL2000 DNA Marker和 SYBR Green均购自宝生物工程(大连)有限公司;胶回收试剂盒购自QIAGEN公司;Trans5K DNA Marker和PCR试剂购自北京全式金生物技术有限公司;其它试剂为进口或国产分析纯;引物合成及测序委托生工生物工程(上海)股份有限公司进行。
1.2 方法
1.2.1 总RNA提取
取普通烟草红花大金元的植株叶片,液氮研磨成粉,Trizol法提取总RNA,1%琼脂糖凝胶电泳检测RNA质量,-80℃保存备用。
1.2.2 引物设计
参考NCBI网站上番茄和马铃薯的前番茄红素异构酶基因序列,利用DNAMAN软件设计上游引物CRTISO-F:5’-ATGGCGTTGAGGTTTCAACCTTTT TC-3’;下游引物CRTISO-R:5’-TCATATCCCTA CAGCATCAAGAAGCTG-3’。引物由生工生物工程(上海)股份有限公司合成。
1.2.3 CRTISO基因CDS全长的克隆
按照反转录试剂盒Reverse transcription system说明书将提取的烟草RNA反转录成cDNA作为模板,然后用CRTISO的特异性引物进行扩增。PCR反应体系为50 μL,模板2 μL,上下游引物各2.5 μL(10 μM),dNTPs 4 μL(10 μM),10×Buffer 5 μL,LA Taq酶0.5 μL,无菌水33.5 μL。1%琼脂糖凝胶电泳检测PCR结果。PCR反应条件为94℃,3 min;94℃,30 s,60℃,30 s,72℃,1.5 min,35 cycles;72℃,10 min;4℃,pause。
用DNA胶回收试剂盒回收PCR产物,纯化后与pMDR19-T Simple Vector连接,连接产物转化DH5α感受态细胞,在LB/Amp/IPTG/X-gal平板上培养,挑选白色菌落,PCR验证阳性克隆,阳性菌液送生工生物工程(上海)股份有限公司测序。
1.2.4 pTRV2-CRTISO重组载体的构建和农杆菌转化
以测序正确的CRTISO基因为模板,分别设计扩增CRTISO关键区域片段的引物。上游引物CRTISOVIGS-F:5’-ATGGTACCGATCTCCTAGCCTTCT TGCTTGC-3’(含KpnⅠ酶切位点);下游引物CRTISO-VIGS-R:5’-CACTCGAGTGTACTCATTG CTTCGGCGAT-3’(含XhoⅠ酶切位点)。PCR反应条件为94℃,3 min;94℃,30 s,58℃,30 s,72℃,45 s,35 cycles;72℃,10 min;4℃,pause。1%琼脂糖凝胶电泳检测PCR结果,胶回收目的片段。
将胶回收目的片段和pTRV2载体分别使用限制性内切酶KpnⅠ、XhoⅠ酶切后连接,转化DH5α感受态细胞,提取重组质粒。以重组质粒为模板,使用上述引物进行PCR验证;KpnⅠ、XhoⅠ对质粒进行双酶切验证。对应的阳性菌液送生工生物工程(上海)股份有限公司测序。然后通过冻融法转化农杆菌GV3101。
1.2.5 农杆菌侵染
由于八氢番茄红素去饱和酶基因(PDS)具有光保护作用,其沉默后植物出现光漂白现象,因此以PDS基因作为报告基因来监控pTRV载体的侵染效率。分别挑取pTRV1、pTRV2、pTRV2-PDS和pTRV2-CRTISO菌落接种至5 mL LB培养基(50 mg/L Kan,25 mg/L Rif),28℃过夜活化。将活化后的菌液分别接入同样的50 mL LB培养基中,再加入10 mM 2-N-吗啉基乙磺酸(MES)和20 μM乙酰丁香酮(AS),28℃过夜培养。低速离心菌液后,用侵染缓冲液(10 mM MgCl2,10 mM MES,200 μM AS)重悬菌体。使用分光光度计在600 nm波长下调整OD值至1.0,室温静置3 h后接种。空白对照注射生理盐水,pTRV1 & pTRV2(TRV)作为阴性对照,pTRV1 & pTRV2-PDS(TRV-PDS)作为阳性对照,pTRV1 & pTRV2-CRTISO(TRV-CRTISO)为实验组,各菌液按照1:1的比例混合,用无针头的无菌注射器注入烟草的下位叶片中。
1.2.6 荧光定量PCR
利用SYBR Green染料法在BIO-RAD荧光定量PCR仪上反应,同时选取26S rRNA作为内标基因,定量分析普通烟草在农杆菌侵染后CRTISO及其下游基因的表达量差异。
荧光定量PCR体系为:1 μL cDNA,10 μL SYBR Green,CRTISO基因上下游引物各1 μL(5 μM),加ddH2O补至20 μL。26S rRNA基因Realtime PCR反应体系为:1 μL cDNA,10 μL SYBR Green,26S上 下 游 引 物 各 1 μL(5 μM), 加ddH2O补至20 μL。PCR反应条件:95℃,3 min;95℃,20 s,60℃,20 s,40 cycles;4℃,pause。对照组和实验组各3个生物学重复,每个生物学重复的样品检测3次,取3个Cp值的平均值,用2-△△Cp法计算。
1.2.7 统计学分析
实验数据使用SPSS 19.0统计软件处理,采用单因素方差分析(One-way ANOVA),P<0.05具有统计学意义,P<0.01具有高度统计学意义。
2 结果与分析
2.1 总RNA的检测
如图1所示,提取的样品总RNA经1%琼脂糖凝胶电泳检测,结果显示有28S和18S两条清晰条带,RNA无降解;NanoDrop 2000超微量分光光度计检测OD260/OD280均在1.8~2.0之间,浓度大约为 870 ng/μL。

图1 RNA电泳结果Fig.1 Agarose gel electrophoresis result of RNA
2.2 CRTISO基因CDS的克隆与检测
以反转录cDNA为模板,PCR扩增CRTISO基因的CDS全长序列。如图2所示,1%琼脂糖凝胶电泳显示在1700 bp左右有清晰、特异的条带。胶回收目的片段,连接到pMDR19-T Simple Vector上进行测序,测序结果为1716 bp。如图3所示,Blast结果显示该序列与马铃薯和番茄的前番茄红素异构酶基因的相似度达93%。上述检测结果表明已成功获得普通烟草CRTISO基因的CDS全长序列。将该基因命名为NtCRTISO,并上传基因序列至GenBank,登录号为KJ909512。
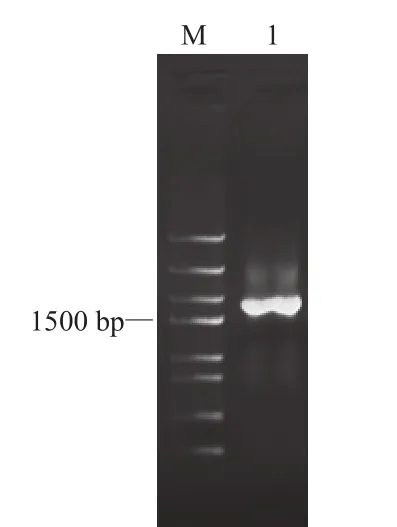
图2 CRTISO的PCR电泳结果Fig.2 Agarose gel electrophoresis result of CRTISO

图3 CRTISO的Blast结果Fig 3 The Blast result of CRTISO
2.3 pTRV2-CRTISO重组载体的检测
以重组质粒pTRV2-CRTISO为模板,使用上下游引物CRTISO-VIGS-F & -R扩增CRTISO基因的关键区域片段。如图4所示,在600 bp左右有清晰的特异性条带。

图4 CRTISO-VIGS片段的PCR电泳结果Fig.4 Agarose gel electrophoresis result of CRTISO-VIGS
使用KpnⅠ、XhoⅠ对重组质粒pTRV2-CRTISO进行双酶切,电泳结果显示在600 bp左右有清晰的特异性条带(图5)。经PCR和双酶切鉴定的阳性质粒送公司测序,结果显示插入的目的片段长度为638 bp,表明pTRV2-CRTISO重组载体构建成功。

图5 重组质粒pTRV2-CRTISO双酶切电泳结果Fig.5 Agarose gel electrophoresis result of double digestion pTRV2-CRTISO
2.4 农杆菌侵染
如图6所示,pTRV-PDS农杆菌侵染10天后,新烟叶出现光漂白现象,表明烟草的PDS基因被成功沉默,TRV载体的侵染效率很高。

图6 pTRV-PDS光漂白效应Fig.6 pTRV-PDS photobleaching in leaves
如图7所示,农杆菌侵染35天后,与空白对照和阴性对照相比,侵染pTRV-CRTISO的烟草叶片产生黄色斑块,表明CRTISO基因被沉默后产生了较明显的表型。

图7 农杆菌侵染本氏烟草35天表型示意图Fig.7 Phenotypes comparison between infected pTRV-CRTISO and controls
2.5 CRTISO基因沉默后对下游基因的影响
实时荧光定量PCR检测农杆菌侵染后CRTISO基因的沉默效果,以及CRTISO下游基因的表达量变化。
如图8所示,与空白对照组(Control)相比,侵染pTRV-CRTISO农杆菌之后,CRTISO基因的表达量明显下降(P<0.05),而阴性对照与空白对照之间差异无统计学意义(P>0.05),结果表明CRTISO基因被有效地沉默。

图8 农杆菌侵染后CRTISO基因的Real-time PCR结果Fig.8 Expression levels of CRTISO after infiltration of pTRVCRTISO
如图9~11所示,CRTISO基因沉默后,导致其下游基因ε-LCY、β-LCY和β-OHase的表达量降低(P<0.05)。

图9 ε-LCY基因的Real-time PCR结果Fig.9 Expression levels of ε-LCY after infiltration
3 讨论
15-顺式-八氢番茄红素是类胡萝卜素合成途径中的第一个成分,它在八氢番茄红素脱氢酶(PDS)的催化下生成ζ-胡萝卜素,随后被ζ-胡萝卜素脱氢酶(ZDS)脱氢生成顺式番茄红素。类胡萝卜素异构酶(CRTISO)就是在此将顺式番茄红素(前番茄红素)催化生成全反式番茄红素,然后在番茄红素ε-和β-两种环化酶的作用下生成α-和β-胡萝卜素,再经过胡萝卜素β-环羟化酶(β-OHase)催化,最终分别生成叶黄素(Lutein)和新黄质(Neoxanthin)[10]。

图10 β-LCY基因的Real-time PCR结果Fig.10 Expression levels of β-LCY after infiltration

图11 β-OHase基因的Real-time PCR结果Fig.11 Expression levels of β-OHase after infiltration
类胡萝卜素是影响烟叶品质的重要成分之一,能够决定调制后烟叶的颜色。它的降解产物香气好,刺激性小,是影响烟叶香气质量的主要组分[11]。近期的研究还表明类胡萝卜素能够提高光合效率,抑制并清除自由基[12],这对烟叶品质的改善和降低烟气中自由基的伤害具有重要意义。因此,提高烟叶中类胡萝卜素的含量不仅能够提高烟叶的品质,还可以改善烤烟的香气风味。
本研究发现普通烟草的CRTISO基因沉默后,类胡萝卜素合成通路下游的基因ε-LCY、β-LCY和β-OHase的表达量降低,烟叶上开始出现黄色斑块,然而烟草本身似乎并未受到严重伤害。新鲜烟叶中的植物色素主要包括叶绿素、β-胡萝卜素、叶黄素、新黄质和紫黄质,而β-OHase负责催化α-和β-胡萝卜素的降解,该基因表达量的降低抑制了胡萝卜素降解反应的进行,可能会导致烟草中胡萝卜素的累积,使烟叶颜色发生改变。这为进一步研究烟草类胡萝卜素合成代谢途径中各基因的功能奠定了良好的基础。
综上所述,由于类胡萝卜素在烟叶中的转化、积累和降解与烟叶的品质和香气密切相关,而类胡萝卜素的合成代谢受光照、温度和遗传等多种因素的调节[13],因此通过分子手段选育类胡萝卜素含量高的优良烟草品种将是我们今后深入研究的方向。
[1] Kim S H,Kim Y H,Ahn Y O,et al.Downregulation of the lycopene ε-cyclase gene increases carotenoid synthesis via the β-branch-specific pathway and enhances salt-stress tolerance in sweetpotato transgenic calli [J].Physiologia Plantarum,2013,147(4): 432-442.
[2] Stahl W,Sies H.β-carotene and other carotenoids in Protection from sunlight [J].The American Journal of Clinical Nutrition,2012,96(5): 1179S-1184S.
[3] Cutler S R,Rodriguez P L,Finkelstein RR,et al.Abscisic acid: emergence of a core signaling network [J].Annual Review of Plant Biology,2010,61: 651-679.
[4]杨懿德,杨焕文.烟草类胡萝卜素的研究进展[J].西南农业学报,2006,B09: 526-530.
[5] Misawa N,Yamano S,Linden H,et al.Functional expression of theErwinia uredovoracarotenoid biosynthesis genecrt Iin transgenic plants showing an increase of β-carotene biosynthesis activity and resistance to the bleaching herbicide norflurazon [J].The Plant Journal,1993,4(5): 833-840.
[6] Mann V,Harker M,Pecker I,et al.Metabolic engineering of astaxanthin production in tobacco flowers [J].Nature Biotechnology,2000,18(8): 888-892.
[7] Kishimoto S,Ohmiya A.Carotenoid isomerase is key determinant of petal color ofCalendulaofficinalis[J].Journal of Biological Chemistry,2012,287(1): 276-285.
[8] Kato M,Ikoma Y,Matsumoto H,et al.Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit [J].Plant Physiology,2004,134(2): 824-837.
[9]陶能国,张继红.纽荷尔脐橙CRTISO基因片段的克隆与分析 [J].吉林农业大学学报,2008,30(1): 28-30.
[10]Cazzonelli C I,Pogson B J.Source to sink: regulation of carotenoid biosynthesis in plants [J].Trends in Plant Science,2010,15(5): 266-274.
[11]过伟民,李伟观,刘阳,等.烤烟类胡萝卜素含量与香气质量的关系[J].烟草科技,2010(1): 51-55.
[12]Darvin M E,Fluhr J W,Meinke MC,et al.Topical betacarotene protects against infra-red-light-induced free radicals [J].Experimental Dermatology,2011,20(2): 125-129.
[13]Quinlan R F,Shumskaya M,Bradbury L M,et al.Synergistic interactions between carotene ring hydroxylases drive lutein formation in plant carotenoid biosynthesis [J].Plant Physiology,2012,160(1): 204-214.
Cloning and functional analysis ofCRTISOgene inNicotiana tabacum
SHI Yanmei1,2,WANG Ran2,YANG Jun2,LUO Zhaopeng2,LI Feng2,WU Mingzhu2,WEI Chunyang2,LIN Fucheng2,QU Lingbo1,3,WEI Pan2
1.College of Chemistry and Molecular Engineering,Zhengzhou University,Zhengzhou 450001,China;
2.Zhengzhou Tobacco Research Institute,China National Tobacco Corporation,Zhengzhou 450001,China;
3.College of Chemistry and Chemical Engineering,Henan University of Technology,Zhengzhou 450001,China
CDS of carotenoid isomerase (CRTISO1716 bp) was obtained from cDNA ofNicotianatabacumby PCR to explore function ofCRTISO.Blast results showed that this gene shared 93% identities withSolanumtuberosumandSolanumlycopersicum.Key nucleotide sequence ofCRTISOwas inserted into tobacco rattle virus (TRV) vector and recombinant pTRV2-CRTISO vector was constructed.Expression level ofCRTISOwas suppressed through VIGS system.Plants infiltrated with pTRV-CRTISO grew abnormal leaves which had yellow patches.Results of Real-time PCR showed that the expression level of ε-LCY,β-LCYandβ-OHasewas depressed due to silence ofCRTISO.Findings of this study may contribute to further research in carotenoid biosynthesis and help to lay theoretical foundation for tobacco breeding.
Nicotianatabacum; CRTISO; Real-time PCR; VIGS
10.3969/j.issn.1004-5708.2014.06.022
Q81 文献标志码:A 文章编号:1004-5708(2014)06-0138-06
郑州烟草研究院科技项目“烟草类胡萝卜素含量的遗传调控和材料验证”(092013CZ0620); 郑州烟草研究院院长科技发展基金项目“普通烟草紫黄质脱环氧化酶基因的克隆和功能分析”(902012CA0120)
史艳梅(1985—),在读博士研究生,主要从事烟草基因功能研究,Tel:0371-67672076,Email:symzgh@163.com
魏 攀(1983—),博士研究生,工程师,主要从事烟草分子生物学研究,Tel:0371-67672073,Email:weipan83@126.com
2014-03-01
猜你喜欢
食品安全导刊(2021年20期)2021-08-30 06:40:36
祝您健康(2019年7期)2019-07-12 03:11:52
天然产物研究与开发(2018年9期)2018-10-08 03:25:38
新疆农垦科技(2016年10期)2016-06-15 20:29:33
中国果菜(2016年9期)2016-03-01 01:28:39
医学研究杂志(2015年8期)2015-06-22 14:00:54
食品与生物技术学报(2013年1期)2013-11-09 00:45:04
食品科学(2013年22期)2013-03-11 18:29:34
食品科学(2013年19期)2013-03-11 18:27:54
食品科学(2013年19期)2013-03-11 18:27:50
