
艾美耳球虫子孢子从宿主细胞中逸出机制的初步研究*
2014-11-10 01:50:58闫鑫磊计永胜田秀玲索静霞刘贤勇
寄生虫与医学昆虫学报 2014年4期
闫鑫磊 计永胜 田秀玲 索静霞 刘贤勇 索 勋
(中国农业大学动物医学院,北京100193)
顶复门原虫包括疟原虫、弓形虫、隐孢子虫、艾美耳球虫等多种寄生虫,可感染包括人在内的几乎所有脊椎动物,由此引发的疟疾、弓形虫病、隐孢子虫病及球虫病给人类社会造成了巨大的损失 (Sibleyetal., 2011)。作为专性胞内寄生病原,顶复门原虫在细胞内的繁殖周期均需经过入侵、胞内增殖及从宿主细胞中逸出3个过程,虫体逸出后可以继续入侵周围细胞开始下一个胞内周期 (Plattneretal., 2008)。在上述3个过程中,逸出作为两次胞内周期的连接点对于顶复门原虫的生存和繁殖具有举足轻重的作用(Roikoetal., 2009)。因此,对该过程的研究有助于了解这类寄生原虫的致病机理,为开发新型有效的抗寄生虫药物提供科学依据。
刚地弓形虫作为顶复门原虫的模式生物,有关其逸出的研究相对成熟。研究表明,刚地弓形虫的逸出是一个主动过程,依赖于虫体或宿主细胞的离子浓度变化,尤其是与钙离子浓度密切相关(Moudyetal., 2001; Arrizabalagaetal., 2004a)。同时,虫体自身的蛋白也参与了逸出过程,如微线体(Microneme)蛋白在逸出过程中可大量分泌(Carruthersetal.,1999),穿孔素样蛋白(TgPLP)能够穿透带虫空泡膜 (Kafsacketal., 2009),敲除刚第弓形虫的钠氢交换通道蛋白TgNHE1 (Arrizabalagaetal., 2004b) 或钙依赖性蛋白激酶1TgCDPK1 (Rezzonicoetal., 1995) 可以影响虫体的运动能力等。
顶复门原虫的另一种重要病原艾美耳球虫主要寄生于畜禽肠道,其从肠上皮细胞逸出的过程是诱发畜禽球虫病的主要原因之一。然而,有关艾美耳球虫逸出的研究较少,更鲜有报道涉及其分子机制。本课题组于近期通过体外研究发现,激活后的淋巴细胞可在非接触的情况下诱导柔嫩艾美耳球虫子孢子从被感染的鸡肾原代细胞(PCKs)中逸出,提示宿主体内可能存在着一条通过诱导胞内病原逸出介导的清除胞内病原的免疫途径 (Dongetal., 2011)。因此,阐明艾美耳球虫的逸出机制不仅可以明晰球虫的致病机理,而且可以为明确逸出介导的机体胞内病原清除途径提供更加可靠的证据。
在弓形虫逸出的研究中,乙醇是一种重要的、常见的诱导工具 (Arrizabalagaetal., 2004)。本研究以乙醇作为逸出诱导剂对球虫子孢子逸出的机制进行了初步解析,为今后对于该过程机制的进一步研究提供了有力的参考。
1 材料与方法
1.1 细胞与虫株
MDBK细胞由本实验室保存,于37℃,5% CO2条件下培养。稳定表达黄色荧光蛋白 (YFP) 的转基因柔嫩艾美耳球虫EtM2e株由本实验室构建 (Liuetal., 2013)。
1.2 子孢子提取
柔嫩艾美耳球虫EtM2e株孢子化卵囊用研磨器研磨至约90%卵囊破碎,3 000 r/ min离心8 min后用孢子囊消化液(10%鸡胆汁,0.25%胰蛋白酶)按1×107个卵囊/毫升混匀并于42℃水浴锅中作用30~50 min至90%子孢子释放,2 500 r/min离心10 min洗去孢子囊消化液后加入DE-52纤维素柱中纯化。收集到的子孢子用血细胞计数板进行计数后用于相关实验。
1.3 逸出实验
MDBK细胞培养于24孔细胞培养板中,每孔加入2×105个EtM2e株子孢子,培养12 h后用PBS缓冲液洗去未入侵的子孢子。用4%乙醇处理20 min或8%乙醇处理3 min后每孔抽取500 μL含有游离子孢子的培养基,利用EtM2e株子孢子稳定表达的黄色荧光蛋白(YFP)为参数,流式细胞仪分析其中10 μL中所含的游离子孢子数量。
1.4 逸出阻断实验
按前述方法洗去未入侵的子孢子后将培养基换成含有终浓度均为10 μmol/L的CytoD或BAPTA-AM(Sigma-Aldrich公司)培养基,37℃条件下孵育30 min,而后用8%乙醇作用3 min,逸出检测方法如1.3所述。
1.5 微线体蛋白分泌诱导/阻断实验
诱导实验:子孢子以2 500 r/min离心5 min,以6×107/mL浓度重悬于DMEM完全培养基中(90% DMEM培养基+10%胎牛血清),将待测样品加入96孔细胞培养板中(46 μL/孔),每孔分别加入4 μL无水乙醇或4 μL DMEM完全培养基于37℃细胞培养箱中孵育3 min。作用后样品以12 000 r/min于4℃条件下离心15 min分离上清与沉淀,两部分均用于Western Blot检测。
阻断实验:子孢子在含有BATPA-AM(终浓度为10 μmol/L)的完全培养基中于37℃环境下作用30 min后用8%乙醇处理3 min,其余实验方法同诱导实验。
1.6 柔嫩艾美耳球虫Mic2蛋白(EtMic2)Western Blot检测
将1.6所述上清和沉淀样品进行SDS-PAGE后转移至PVDF膜上进行Western Blot检测。上清检测组孵育的一抗为本实验室保存的抗EtMic2单抗,1∶200稀释,二抗为HRP-羊抗小鼠IgG,1∶2 000稀释;沉淀作为内参孵育的一抗为抗黄色荧光蛋白多抗,1∶2 000稀释,二抗为HRP-羊抗兔IgG,1∶2 000稀释。
1.7 数据统计
实验数据用SPSS 13.0统计软件进行分析,各组数据均用平均值±标准差表示,采用T检验比较各组间数据差异,将P<0.05做为差异具有统计学显著性的标准。
2 结果
2.1 乙醇诱导柔嫩艾美耳球虫子孢子从MDBK细胞中逸出
用4%乙醇处理感染子孢子的MDBK细胞20 min,收集含有游离子孢子的细胞培养液利用流式细胞仪分析。经4%乙醇处理后,细胞培养液中的游离子孢子数量远远大于未经乙醇处理组(图1-A);将乙醇浓度提高至8%后,作用3 min便达到4%乙醇作用20 min的效果(图1-B)。

图1 乙醇可诱导柔嫩艾美耳球虫子孢子从MDBK细胞中逸出Fig. 1 Ethanol could induce rapid egress of E. tenella sporozoites from infected MDBK cellsA: 4%乙醇诱导20 min 子孢子逸出情况;B: 8% 乙醇诱导3 min子孢子逸出情况。图中所示为均值±标准差。A: 4% ethanol for 20 min; B: 8% etahnol for 3 min. Each bar represents the mean with SEM (n=5).
2.2 子孢子的逸出过程与其运动能力相关
利用肌动蛋白抑制剂CytoD处理感染子孢子的细胞充分抑制虫体运动能力后,用8%乙醇处理细胞3 min。通过流式细胞仪检测结果可以看出,与未用CytoD处理组相比,经CytoD处理后的子孢子在乙醇诱导下逸出的能力显著下降(图2)。
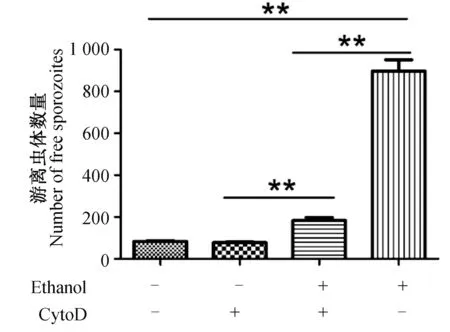
图2 细胞松弛素D对逸出的影响检测Fig.2 Determination of the effect of cytochalasin D on rapid egress of sporozoites图中所示为均值±标准差;“+”表示加入该种试剂,“﹣”表示无该种试剂作用。Each bar represents the mean with SEM (n =6).
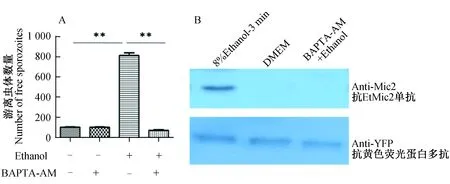
图4 虫体内钙离子对子孢子的逸出起关键作用Fig. 4 Intracellular Ca2+ of the parasite plays a vital function to egressA: 子孢子感染的MDBK细胞用终浓度为10 μmol/L的BAPTA-AM于37 ℃条件下孵育30 min后用8%乙醇作用3 min;B: 用8%乙醇处理10 μmol/L BAPTA-AM孵育30 min后的子孢子。A: Intracellular sporozoites were treated with 8% ethanol for 3 min either in the presence or absence of 10 μmol/L BAPTA-AM for 30 min. Each bar represents the mean with SEM (n=4); B: Free sporozoites were treated with 8% ethanol for 3 min at 37 ℃ either in the presence or absence of 10 μmol/L BAPTA-AM for 30 min.
2.3 乙醇可诱导子孢子微线体蛋白的大量分泌
微线体蛋白的分泌量与顶复门原虫的运动能力呈现相关性,在刚地弓形虫逸出的研究中常以微线体蛋白的分泌指示逸出现象的发生。本研究选择EtMic2蛋白来反映虫体在乙醇刺激下微线体的活动情况。通过Western Blot结果可以看出,经8%乙醇处理后子孢子EtMic2蛋白发生大量分泌(图3)。

图3 乙醇可诱导虫体微线体蛋白的分泌Fig 3 Ethanol could stimulate the discharge of microneme from sporozoites以沉淀中虫体表达的黄色荧光蛋白作为内参。Sporozoites expressing YFP from parasites whole-cell lysate was used as a control.
2.4 虫体内Ca2+在逸出过程中起关键作用
使用内源性Ca2+阻断剂BAPTA-AM作用感染子孢子的细胞后,用8%乙醇处理被感染的细胞3 min。与未被阻断的子孢子相比,Ca2+阻断后子孢子在乙醇诱导下的逸出能力完全丧失(P<0.01);同时,虫体内Ca2+被阻断后虫体在乙醇诱导下微线体蛋白的分泌能力完全受到抑制(图4)。
3 讨论
本研究发现乙醇诱导的子孢子逸出与虫体肌动蛋白介导的虫体运动相关,虫体发生逸出时伴随微线体蛋白的大量分泌,更重要的发现是上述过程均受到虫体内Ca2+的直接调控。上述研究结果填补了球虫子孢子逸出机制研究的空白,初步建立了子孢子逸出研究的模型,为今后的进一步研究提供了重要的参考。
顶复门原虫逸出过程需要多种虫体蛋白的参与,例如微线体蛋白的分泌会大量增加。弓形虫微线体蛋白2(TgMic2)在自然逸出或是乙醇刺激的情况下分泌量显著增加 (Nagamuneetal., 2008a)。本研究得到类似结果,即以球虫微线体蛋白 EtMic2释放作为标准指示逸出的发生,结果显示在未经刺激的情况下无法检测到EtMic2的分泌,而当8%乙醇诱导3 min后该蛋白的分泌量明显升高,说明该蛋白可指示子孢子逸出的发生。因此,该发现也提示我们可以将EtMic2作为艾美耳球虫在逸出的体内研究中的一个重要候选标记物。
本研究利用乙醇为工具初步阐明了Ca2+在艾美耳球虫子孢子逸出中的作用和重要性。相关研究表明弓形虫逸出依赖于虫体胞浆内Ca2+的瞬间升高,这些Ca2+来源于虫体内质网 (Melzeretal., 2008);通过钙离子载体A13287人为将弓形虫内Ca2+浓度迅速升高可导致多种微线体蛋白的大量分泌 (Carruthersetal.,1999), 而在虫体内原性Ca2+被螯合的情况下分泌则完全受阻 (Nagamuneetal., 2008b)。有关疟原虫的研究发现Ca2+在其的逸出过程中同样起到关键作用:提高宿主细胞亦或是虫体自身Ca2+浓度均能导致恶性疟原虫Plasmodiumfalciparum裂殖子从红细胞中快速逸出 (Chandramohanadasetal., 2009; Cruzetal., 2011);而在逸出过程中微线体的分泌则必须依赖虫体自身Ca2+浓度的升高 (Gargetal., 2013)。这两种顶复门原虫的模式生物Ca2+与逸出关系的研究与本研究基本相似,这提示我们Ca2+在顶复门原虫逸出过程中的作用可能具有保守性。
综上所述,本研究初步剖析了柔嫩艾美耳球虫逸出的机制,为进一步明确球虫致病的分子机制提供了新的研究方向,并为研发新型抗球虫药物和高效球虫疫苗提供了一定的科学依据。
猜你喜欢
动物医学进展(2024年5期)2024-05-09 08:09:50
锦绣·下旬刊(2022年1期)2022-05-16 07:30:45
现代畜牧科技(2021年3期)2021-07-21 08:42:04
中国工作犬业(2021年3期)2021-03-27 15:10:41
中国预防兽医学报(2020年4期)2020-01-15 04:06:32
河南畜牧兽医(2020年23期)2020-01-10 03:19:52
家庭百事通·健康一点通(2019年8期)2019-08-29 02:50:49
畜牧与饲料科学(2016年9期)2016-11-01 21:07:43
河南畜牧兽医(2015年13期)2015-08-15 00:46:58
中国人兽共患病学报(2015年9期)2015-05-09 01:03:45
