
吗啡成瘾大鼠边缘下区脑电活动的无线遥测及分析*
2014-11-08 02:27朱再满白家明潘群皖
中国病理生理杂志 2014年1期
李 晶, 朱再满, 白家明, 潘群皖△, 李 敏, 虞 冉
阿片类药物成瘾是一种以强迫性用药、觅药为核心症状的慢性、复发性脑病[1]。近年来,有研究显示[2],内侧前额皮层(medial prefrontal cortex,mPFC)参与阿片成瘾的形成和复吸过程,其作用机制与mPFC的学习记忆功能密切相关。边缘下区(infralimbic cortex,IL)位于 mPFC 的腹侧[3-4],是条件性恐惧和药物成瘾记忆消退的重要脑区。有关IL区如何参与阿片成瘾的形成过程,IL区脑电活动的改变对成瘾过程有何影响目前尚不清楚。本实验通过制备吗啡成瘾大鼠条件性位置偏爱(conditioned place preference,CPP)模型,在 CPP箱内视频监控大鼠CPP行为的改变,同时利用无线遥测脑电系统实时观察大鼠IL区脑电活动的相应变化,以期从神经电生理学视角探索IL区在大鼠吗啡成瘾过程中的作用及其脑电活动改变对吗啡成瘾过程的影响,为阿片成瘾和复吸治疗另辟新径。这种针对阿片成瘾大鼠CPP行为变化过程中IL区脑电活动的实时记录与分析,迄今少见报道。
材料和方法
1 动物、主要药品及仪器
1.1 动物 选用成年雄性SD大鼠[合格证号为SCXK(苏)2009-0001]24只,SPF级,体重290~320 g,购自苏州爱尔麦特科技有限公司。自由进食、饮水,12 h光照,室温控制22~25℃。将大鼠随机分成2组(吗啡戒断组和生理盐水对照组)。
1.2 主要药品 盐酸吗啡注射液(皮下注射),沈阳第一制药公司,批号为110508-2,由安徽省医药股份有限公司提供。
1.3 仪器装置 大鼠脑立体定位仪(第二军医大学);CPP视频分析装置(上海吉量软件科技有限公司);生理无线遥测脑电系统(成都泰盟软件有限公司)。
2 方法
2.1 脑立体定位埋植电极手术 以1%戊巴比妥钠(50 mg·kg-1)腹腔注射麻醉大鼠,将其俯卧位(使门齿杆低于耳间连线3 mm,调整前囟与后囟至同一水平面)固定在脑立体定位仪上。剪毛、消毒,切开头皮,剥离骨膜,暴露骨缝,参照大鼠脑立体定位图谱[5]定位左侧IL区(前囟前3.2 mm,中缝向左旁开0.6 mm,硬膜下5.2 mm),定位点钻孔。记录电极使用直径0.3 mm的漆包镍铬丝,尖端裸露2 mm,植入定位孔硬膜下5.2 mm,同时放置参考电极于头皮下,采用自凝牙托粉封固。术后恢复7 d,连续5 d给予青霉素(400 kU·kg-1)抗感染,每天1次。
2.2 吗啡成瘾大鼠CPP模型的制备 模型制备前期,对大鼠进行CPP行为预测试。打开CPP装置的通道隔板,使大鼠在两箱中自由活动15 min,每日1次,共3 d,比较大鼠在两箱中各自平均停留时间,同时采用CPP视频系统分析软件绘制大鼠活动轨迹图。比较结果表明,大鼠均偏爱黑箱,因此模型制备阶段,设非天然偏爱箱(白箱)为伴药箱。模型制备期,用隔板关闭黑、白箱间通道。戒断组大鼠每日皮下注射吗啡及生理盐水(NS)各1次[6],注射吗啡后投入白箱45 min,注射NS后投入黑箱45 min,2次注射间隔8 h,CPP训练共8 d,吗啡首日剂量10 mg·kg-1,以后逐日递增 10 mg·kg-1,至 6 d 达到 60 mg·kg-1,7~8 d维持该剂量,对照组大鼠以相同方法注射NS。模型制备结束后打开通道,于戒断期1~3 d,通过CPP视频系统对大鼠的CPP行为进行测试,计测其15 min内在白箱中停留时间,同时绘制大鼠活动轨迹图。经组间比较及组内给药前后对照,明确大鼠CPP的形成。
2.3 大鼠IL区脑电的遥测及分析 记录方法参照文献[7],于造模前及戒断期,在CPP装置的视频箱内,使用无线遥测脑电系统实时计测各组大鼠在不同行为状态下左侧IL区的脑电活动。行为状态类型包括:白箱停留、黑箱停留、白-黑箱穿梭及黑-白箱穿梭。利用无线遥测脑电系统的快速傅里叶变换(fast Fourier transform,FFT)功能处理分析原始脑电,原始脑电图的横坐标表示脑电信号的采样时点,纵坐标表示脑电波幅。高通及低通滤波分别为0.1 Hz和 100 Hz,采样率 500 Hz。
3 统计学处理
采用SPSS 17.0统计软件分析,数据以均数±标准差(mean±SD)表示。采用独立样本t检验及重复测量的方差分析。若方差不齐时,采用非参数检验。以P<0.05为差异有统计学意义。
结 果
1 吗啡成瘾大鼠CPP模型检测
如图1和表1所示,注射8 d吗啡的戒断组大鼠,戒断1~3 d在白箱中停留时间明显长于同阶段注射NS的对照组大鼠(P<0.01)。与造模前白箱停留时间相比,造模后对照组大鼠在白箱中停留时间无显著变化(P>0.05);戒断组大鼠戒断1~3 d在白箱中停留时间均显著延长(P<0.01),并且戒断组大鼠在白箱中停留时间随戒断天数增加逐渐延长,戒断3 d在白箱中停留时间最长,表明戒断3 d位置偏爱最明显。上述结果提示,戒断1~3 d戒断组大鼠对白箱产生稳定的CPP,模型检测验证制模成功。

Figure 1.The maps of activity tracks of the rats in withdrawal group before and after modeling.A:before modeling;B:withdrawal 1 d;C:withdrawal 2 d;D:withdrawal 3 d.图1 戒断组大鼠造模前后活动轨迹图
2 大鼠在黑、白箱中停留或经黑、白箱间穿梭时,IL区遥测脑电的变化
造模前,戒断组大鼠在黑、白箱中停留或经黑、白箱间穿梭时IL区遥测脑电与对照组大鼠同一行为状态下的IL区脑电无明显差异(P>0.05)。
造模后戒断3 d,戒断组与对照组大鼠相比,IL区遥测脑电表现为:当戒断组大鼠停留于白箱时,δ波明显减少(P<0.05),β波(以β2波增加为主)明显增加(P<0.01),θ波及α波组间比较无显著差异(P>0.05),见图2;当戒断组大鼠停留于黑箱时,IL区脑电各波与对照组比较差异无统计学意义(P>0.05),见图3;当戒断组大鼠由黑箱向白箱穿梭时,IL区脑电δ波显著增加(P<0.01),α波(包括α1波和α2波)显著减少(P<0.01),β波(包括 β1波和β2波)显著减少(P<0.01),仅θ波组间差异不显著(P>0.05),见图4;当戒断组大鼠由白箱向黑箱穿梭时,IL区脑电各波与对照组相比无明显改变(P>0.05),见图 5。

表1 大鼠在白箱中停留时间比较Table 1.Comparison of the spending time of the rats in the white box(s.Mean ±SD.n=12)

Figure 2.The telemetry electrical activity of left IL when rats stayed in white box on withdrawal 3 d.Mean±SD.n=12.*P <0.05,**P <0.01 vs control group.图2 戒断3 d大鼠停留于白箱时左侧IL遥测脑电活动
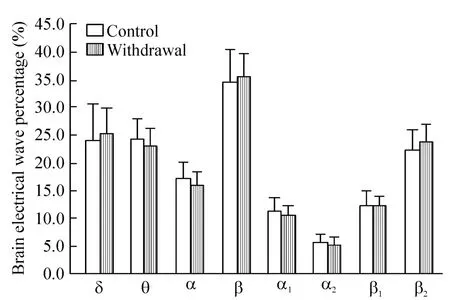
Figure 3.The telemetry electrical activity of left IL when rats stayed in black box on withdrawal 3 d.Mean±SD.n=12.图3 戒断3 d大鼠停留于黑箱时左侧IL遥测脑电活动
讨 论

Figure 4.The telemetry electrical activity of left IL when rats shuttled from black box to white box on withdrawal 3 d.Mean± SD.n=12.*P < 0.05,**P < 0.01 vs control group.图4 戒断3 d大鼠黑-白箱穿梭时左侧IL遥测脑电活动
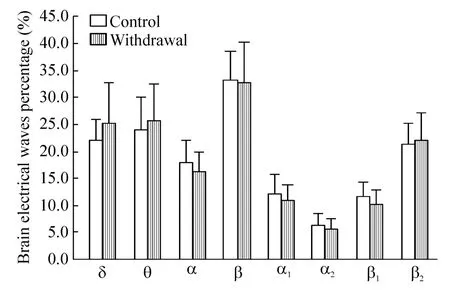
Figure 5.The telemetry electrical activity of left IL when rats shuttled from white box to black box on withdrawal 3 d.Mean±SD.n=12.图5 戒断3 d大鼠白-黑箱穿梭时左侧IL遥测脑电活动
阿片类药物成瘾者具有强烈的心理渴求(觅药动机),在药物耐受、依赖和强迫戒断后,一旦药物相关环境线索暴露,觅药动机引燃,即可产生复吸。相关研究显示,在阿片成瘾的形成中,用药行为由最初的可控性过渡到强迫性用药,该过程与mPFC的功能异常密切相关,mPFC是中脑腹侧背盖区多巴胺神经元的主要投射区,参与药物成瘾相关的学习、记忆过程[8]。在mPFC中,位于腹侧的IL区是调控药物成瘾的重要脑区之一,主要行使认知、行为决策、情绪表达调节等功能[9]。因此本实验将IL区作为研究焦点,探讨吗啡成瘾大鼠CPP表现最强烈时期(戒断3 d)IL区脑电的变化,这项实验将为阿片成瘾及复吸机制的深入研究提供参考。
在IL区参与阿片成瘾形成机制的长期探索中,部分研究认为IL区具有觅药抑制功能,如IL区注射 α-氨基-3-羟基-5-甲基-4-异恶唑丙酸(α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid,AMPA)和β2肾上腺素能受体激动剂可抑制大鼠海洛因觅药行为恢复[9],选择性失活IL区神经元则增加海洛因诱导的大鼠CPP恢复[10];进一步研究表明IL区也具有促进觅药功能,如Bossert等[11]报道IL区至伏隔核壳部的谷氨酸能投射纤维激活引发线索诱导的大鼠海洛因觅药复燃,注射γ-氨基丁酸(γaminobutyric acid,GABA)受体激动剂至IL区使其失活,可消退复燃现象。从上述文献分析提示IL区神经元的功能或许具有双重性:即一部分IL区神经元抑制觅药;而另一部分IL区神经元促进觅药。IL区的这种双重性体现在本实验中:(1)戒断3 d,当吗啡成瘾大鼠(与对照组相比)由黑箱向白箱穿梭时,IL区脑电活动出现一过性δ波明显增加,α波(α1、α2)及 β 波(β1、β2)明显减少,结果提示 IL 区神经元功能受到短时压抑。IL区是控制冲动行为的重要脑区[12],阿片类药物的长期使用可损害IL区的结构和功能,使药物成瘾者对冲动行为控制能力下降。对吗啡戒断大鼠前额联络皮层的脑电分析研究显示[13],吗啡摄入可引起前额皮层部分神经元功能受到抑制、兴奋性降低,引起该区脑电慢波增加。考虑本实验中受抑制的部分IL区神经元,可能参与对大鼠觅药行为的抑制性调控(IL抑制区),当其功能受到压抑时,对大鼠觅药冲动的控制力削弱,则大鼠向白箱穿梭觅药行为表达增强。(2)当吗啡成瘾大鼠进入白箱并停留时,IL区脑电呈现δ波显著减少,β波(主要是β2)显著增加。有研究发现[14-15],药物相关环境线索暴露可激活中脑腹侧背盖区→杏仁核→IL区的投射通路,通过IL区激活的部分神经元集群进一步编码和调控阿片奖赏效应与药物相关环境线索联合的学习过程。在本实验中,吗啡成瘾大鼠停留于白箱(伴药箱)时,药物相关环境线索的充分暴露可能激活了具有促进觅药功能的IL区神经元,使其脑电呈现慢波(δ波)减少,快波(β波)增加的趋势。(3)当吗啡成瘾大鼠由白箱向黑箱穿梭或停留于黑箱时,IL区脑电各波与对照组比较无显著差异。表明在上述行为状态下,吗啡成瘾大鼠IL区脑电活动未发生特异性改变;黑箱(非伴药箱)作为非药物相关环境线索,在IL区脑电活动的改变过程中未产生显著影响。
综合上述实验结果分析,吗啡成瘾大鼠穿梭觅药与停留于药物相关环境线索中引发的IL区脑电活动改变可能存在不同机制;IL区神经元功能在大鼠吗啡成瘾过程中或许起双重调控作用。本研究发现,在给予大鼠吗啡注射8 d后,具有抑制觅药功能的IL区神经元出现功能障碍,不能有效遏制觅药动机及其行为;具有促进觅药功能的IL区神经元在吗啡作用下出现激活,可增强动物觅药渴求。伴随不同神经元功能的改变,IL区相应部位产生特征性的生物电变化,例如吗啡成瘾大鼠由黑箱向白箱穿梭时,抑制觅药功能的IL区神经元表现兴奋性降低,促进觅药功能的IL区神经元少量激活,综合脑电活动改变为慢波活动增多;当吗啡成瘾大鼠进入白箱并停留时,少量激活的IL区促进觅药功能的神经元在充分场景暴露下进一步激活,这种神经元的兴奋引起的脑电快波掩盖了IL区抑制觅药功能神经元的慢波活动,因此成瘾大鼠白箱停留脑电表现为快波增多。
[1] 薛丽芬,薛言学,何 佳,等.下边缘皮质中神经元周围基质网络(PNNs)的降解联合消退训练能够抑制吗啡奖赏记忆的复燃[J].中国药物依赖性杂志,2012,21(2):105-110.
[2] 郝 悦,杨静玉,吴春福.内侧前额叶皮层损毁对小鼠位置偏爱及学习记忆的影响[J].中国药理通讯,2004,21(3):7-8.
[3] 王益琦,刘惠芬.觅药行为消退的研究进展[J].中国药物依赖性杂志,2011,20(5):334-339.
[4] 李培培,张丽丽,韦 美,等.条件性恐惧大鼠边缘下区Cdk5激酶活性、caspase-3表达以及突触结构的变化[J].心理学报,2011,43(5):544-552.
[5] Paxinos G,Watson C.The rat brain in stereotaxic coordinates[M].6th ed.San Diego:Academic Press,2007:50-58.
[6] 顾珊智,李生斌.各种实验因素对建立大鼠吗啡依赖模型的影响[J].西安交通大学学报(医学版),2004,25(5):446-448.
[7] 潘群皖,黄 磊,朱再满,等.海洛因诱导的条件性位置偏爱大鼠额叶联络皮层脑电的无线遥测及其分析[J].中国病理生理杂志,2012,28(9):1589-1596.
[8] 杨 烨,张春燕,王 斌,等.吗啡成瘾及戒断大鼠前额叶皮质蛋白质组变化的初步研究[J].中国应用生理学杂志,2011,27(2):184-186.
[9] 王益琦,陈俊丰,庄丁丁,等.边缘下皮层在海洛因觅药行为消退和恢复中的作用[J].中国药理学与毒理学杂志,2012,26(3):449.
[10] Ovari J,Leri F.Inactivation of the ventromedial prefrontal cortex mimics re-emergence of heroin seeking caused by heroin reconditioning[J].Neurosci Lett,2008,444(1):52-55.
[11] Bossert JM,Stern AL,Theberge FR,et al.Role of projections from ventral medial prefrontal cortex to nucleus accumbens shell in context-induced reinstatement of heroin seeking[J].J Neurosci,2012,32(14):4982-4991.
[12]曾 红,郭斯萍.易感人格的工作记忆特征对药物成瘾的调节机制[J].心理科学进展,2011,19(3):420-426.
[13]李 晶,潘群皖,朱再满.吗啡依赖大鼠戒断期额叶联络皮层脑电活动的分析[J].中国药物依赖性杂志,2012,21(4):273-277.
[14]Kalivas PW,Volkow ND.The neural basis of addiction:a pathology of motivation and choice[J].Am J Psychiatry,2005,162(8):1403-1413.
[15] Bossert JM,Stern AL,Theberge FR,et al.Ventral medial prefrontal cortex neuronal ensembles mediate context-induced relapse to heroin[J].Nat Neurosci,2011,14(4):420-422.
猜你喜欢
遥测遥控(2022年1期)2022-02-11
环球时报(2021-07-22)2021-07-22
儿童故事画报·智力大王(2020年1期)2020-04-28
民用飞机设计与研究(2019年2期)2019-08-05
民用飞机设计与研究(2019年2期)2019-08-05
中国生物医学工程学报(2019年6期)2019-07-16
电子制作(2019年11期)2019-07-04
中央民族大学学报(自然科学版)(2017年3期)2017-10-22
自动化学报(2017年11期)2017-04-04
中国生物医学工程学报(2017年6期)2017-02-10
