
环孢素对癫痫大鼠海马氧化应激及线粒体能量代谢的影响*
2014-11-08 02:27王胜军薄其玉赵秀鹤曹丽丽迟兆富
中国病理生理杂志 2014年1期
王胜军, 薄 宁, 薄其玉, 赵秀鹤, 曹丽丽, 迟兆富
环孢素(cyclosporin;或环孢素A,cyclosporin A,CsA)可抑制T细胞的活化从而抑制免疫系统的活性,被广泛用于器官移植排斥的治疗。环孢素还可与线粒体膜通透性转换孔(mitochondrial permeability transition pore,mPTP)的亲环蛋白D结合,阻断线粒体膜通透性转换(mitochondrial permeability transition,MPT),调节线粒体功能及细胞存亡[1-2]。线粒体是细胞的“动力工厂”,线粒体呼吸链复合物(mi-tochondrial respiratory chain complexes)通过氧化磷酸化以ATP的形式为细胞提供能量。大量研究表明,痫性发作可使线粒体结构受损、能量代谢障碍,而线粒体结构和功能异常可加重神经元损伤,引起神经元兴奋性增高[3-5]。
癫痫发作尤其是癫痫持续状态(status epilepticus,SE)时脑组织产生大量活性氧(reactive oxygen species,ROS),作用于线粒体,产生毒性作用[6]。丙二醛(malondialdehyde,MDA)是脂质过氧化的终产物,能反映组织自由基的水平;超氧化物歧化酶(superoxide dismutase,SOD)是机体内重要的自由基清除酶,对ROS有清除作用。ROS可造成mPTP开放,导致线粒体功能障碍,因此,抑制ROS产生可有效减轻神经元的损伤[7]。
研究发现,环孢素可减少癫痫中神经元损伤并减少癫痫发作频率[8],但环孢素是否参与调节癫痫大鼠海马神经元氧化应激及线粒体能量代谢仍未见报道。本文利用匹鲁卡品(pilocarpine)诱导的癫痫持续状态动物模型来评价环孢素对癫痫大鼠海马氧化应激和线粒体损伤的调节作用及分子机制。
材料和方法
1 材料
雄性Wistar大鼠48只,6~8周龄,体重(250±50)g,由山东大学实验动物中心提供。环孢素、匹鲁卡品、丙稀酰胺、泛醌、鱼藤酮、抗霉素A、十二烷基麦芽苷、细胞色素 C和泛醇2购自 Sigma。MDA、SOD和ATP试剂盒购自南京建成生物工程研究所。
2 动物模型的制作与分组
48只大鼠随机分为空白对照组(control组)、亲环素对照组(CsA组)、匹鲁卡品致癫痫持续状态组(SE组)和环孢素干预组(SE+CsA组),每组12只。SE组大鼠首先皮下注射氢溴酸东莨菪碱1 mg/kg,用以拮抗匹鲁卡品的外周胆碱能反应,30 min后腹腔注射匹鲁卡品340 mg/kg,观察大鼠的行为学变化,达到Racine 4~5级标准的大鼠被认为癫痫模型制作成功,大鼠癫痫发作持续1 h后给予腹腔注射地西泮10 mg/kg终止发作。SE+CsA组大鼠为匹鲁卡品注射前30 min腹腔注射环孢素(10 mg/kg)。CsA组大鼠仅注射环孢素而不注射匹鲁卡品。对照组大鼠注射等量生理盐水代替匹鲁卡品,各组动物分别在腹腔注射24 h后处死。
3 方法
3.1 脑组织样本制备 大鼠迅速断头取脑,分离海马,冰浴中研磨,600×g离心15 min,取上清液为海马组织成分。海马组织加入裂解液,冰浴中充分研磨,800×g离心5 min,取上清液12 000 ×g离心10 min,沉淀为线粒体成分,用保存液悬浮沉淀,采用Bradford法检测线粒体蛋白含量。
3.2 线粒体膜通透性转换的检测 线粒体膜通透性转换增大时线粒体膜内外渗透失衡,导致吸光度明显减少。线粒体蛋白加入测定液(230 mmol/L甘露醇,70 mmol/L蔗糖,3 mmol/L HEPES缓冲液,25℃,pH 7.4),在540 nm波长处用分光光度计测定吸光度值。
3.3 MDA和SOD的检测 利用MDA可与硫代巴比妥酸缩合,在530 nm处有最大吸收峰,检测其活性。利用超氧阴离子自由基与氧化羟胺形成亚硝酸盐,与显色剂作用后在550 nm处有最大吸收峰检测SOD活性,按照试剂盒说明书进行操作。
3.4 ATP含量的检测 利用肌酸激酶催化三磷酸腺苷和肌酸生成磷酸肌酸,用磷钼酸比色法检测海马组织中ATP含量,实验操作按照试剂盒说明书进行。
3.5 线粒体呼吸链复合物I、III活性的检测 参照Gao等[5]报道的方法,将线粒体成分溶于分析介质,应用分光光度计检测活性。线粒体呼吸链复合物I活性检测:溶于介质的线粒体反复冻融3次,加入BSA(2.5 g/L)、抗霉素 A(2 mg/L)、NADH(0.13 mmol/L)、氰化钾(2 mmol/L)和泛醌(50 μmol/L),混匀后在340 nm处检测吸光度。线粒体呼吸链复合物III活性检测:溶于分析介质的线粒体成分加入细胞色素 C(15 μmol/L)、鱼藤酮(2 mg/L)、十二烷基麦芽苷(0.6 mmol/L)和泛醇 2(35 μmol/L),在550 nm处检测吸光度的变化。
4 统计学处理
数据以均数±标准差(mean±SD)表示,多个样本均数比较采用单因素方差分析(One-way ANOVA),两两比较采用Newman-Keuls检验。统计分析采用SPSS 13.0软件,以P<0.05为差异有统计学意义。
结 果
1 环孢素可抑制癫痫大鼠海马组织线粒体通透性转变
SE组大鼠海马组织的MPT值明显高于对照组(P<0.05);CsA+SE组大鼠海马组织的MPT值显著低于SE组(P<0.05);CsA对照组与正常对照组MPT值之间的差异无统计学意义,见表1。
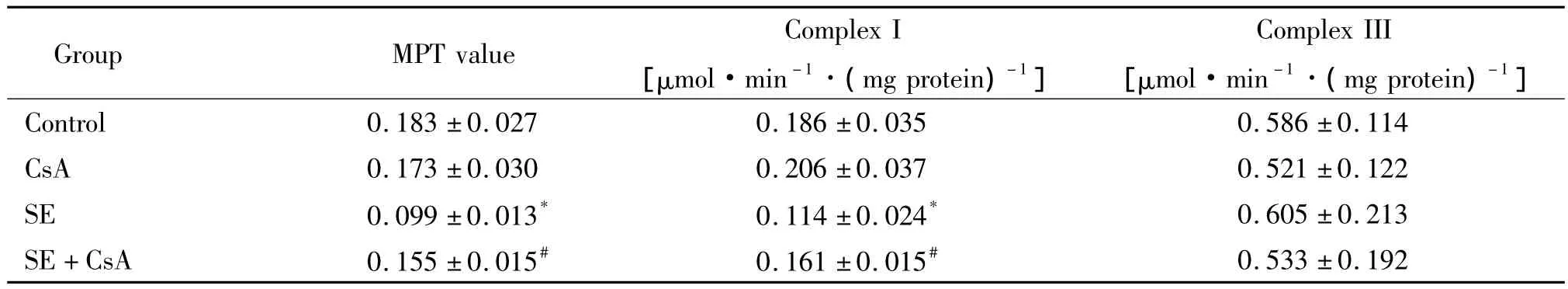
表1 环孢素对癫痫大鼠海马组织线粒体膜通透性转换及呼吸链复合物I、III活性的调节Table 1.Effect of cyclosporin A(CsA)on mitochondrial permeability transition(MPT),mitochondrial respiratory chain complex I and III in rat hippocampus after status epilepticus(SE)(Mean±SD.n=6)
2 环孢素能减少癫痫大鼠海马组织的脂质过氧化水平并提高SOD活性
SE组大鼠海马组织的MDA明显高于对照组,SOD明显低于对照组(P<0.05);CsA+SE组大鼠海马组织的MDA含量显著低于SE组,SOD活性显著高于SE组(P<0.05);CsA对照组与正常对照组之间的 MDA含量和 SOD活性无统计学差异,见表2。
3 环孢素增加癫痫大鼠海马组织的ATP含量
SE组大鼠海马组织的ATP含量明显高于对照组(P<0.05);CsA+SE组大鼠海马组织的ATP含量明显低于SE组(P<0.05);CsA对照组与正常对照组的ATP含量值无统计学差异,见表2。

表2 环孢素对癫痫大鼠海马组织MDA、SOD和ATP水平的调节Table 2.Effect of cyclosporin A(CsA)on MDA,SOD and ATP in rat hippocampus after status epilepticus(SE)(Mean±SD.n=6)
4 环孢素调节癫痫大鼠海马组织线粒体呼吸链复合物的活性
SE组大鼠海马组织呼吸链复合物I的活性低于对照组(P<0.05);CsA+SE组大鼠海马组织呼吸链复合物I的活性高于SE组(P<0.05);CsA对照组与正常对照组的呼吸链复合物I活性差异无统计学意义。各实验组与对照组间的呼吸链复合物III活性差异无统计学意义,见表1。
讨 论
mPTP是一种横跨于线粒体外膜和内膜之间的跨膜多蛋白孔道,其分子组成尚未完全清楚,生理状态的mPTP能通过小于1 500 kD分子量的非特异性物质[2,9]。在受到特定刺激后mPTP开放,使大分子质量物质非选择性地自由通过线粒体膜,造成线粒体内离子平衡紊乱、线粒体肿胀、氧化磷酸化解耦联、ATP水平下降,同时还可引起线粒体膜电位去极化,使细胞色素C、凋亡诱导因子等凋亡活性物质释放入胞质,诱导细胞死亡。另外,线粒体呼吸功能紊乱,组成线粒体抗氧化防御系统的还原型烟酰胺二核苷酸流失,使线粒体产生大量的ROS,进一步加重线粒体的损伤[9]。mPTP开放受亲环蛋白D、电压依赖性离子通道与腺嘌呤核苷酸转位酶的调节,环孢素可通过结合亲环蛋白D来抑制mPTP开放,从而引起膜通透性的转变[1,9]。本研究利用分光光度法检测线粒体渗透性转换能力的变化,间接反映mPTP的开放情况。本研究结果表明,匹鲁卡品诱发的大鼠癫痫持续状态可导致海马线粒体膜通透性转换孔的大量开放,表现为线粒体通透性转变值减小,而环孢素干预后可使神经元线粒体通透性转变值仍保持较高的水平,提示它可明显抑制mPTP的开放。
癫痫发作时产生的大量谷氨酸,可使神经元钙离子浓度增加,顺着电化学梯度进入线粒体内,导致线粒体内钙超载,从而诱导mPTP开放。此外,癫痫造成的氧化应激也是调节mPTP开放的一个重要因素。虽然脑组织存在多种的抗氧化防御系统,但癫痫发作时可产生大量ROS[6]。ROS一方面攻击神经元核DNA,激活核酸修复酶聚(ADP核糖)聚合酶,加重细胞能量消耗及激活线粒体凋亡诱导蛋白和炎症因子[10-11]。另一方面ROS可以通过氧化mPTP上的硫醇而触发mPTP开放,损伤线粒体DNA导致呼吸链复合物亚基表达的改变,影响线粒体的能量代谢[12]。研究发现,线粒体抗氧化酶 MnSOD部分缺失可增加脑的兴奋性,MnSOD表达下调的转基因小鼠癫痫的发生率明显增加,提示线粒体的氧化应激可能是癫痫形成的重要机制[13]。ROS可造成线粒体结构数目、能量代谢及生物合成异常,导致神经元功能障碍,促进癫痫的形成与反复发作[5,7-8]。线粒体既是ROS易损伤的细胞器,也是ROS产生的重要场所,ROS可诱导线粒体mPTP开放,进而加重线粒体肿胀及线粒体呼吸链功能,促进ROS的产生[6,9]。本研究显示,癫痫发作后海马组织中脂质过氧化产物MDA含量明显升高,同时自由基清除酶SOD的活性明显降低,这与以往的报道一致[7,14]。本研究发现,环孢素可明显抑制癫痫发作造成的海马组织的氧化损伤,提示减少mPTP开放可有效减少癫痫发作诱发的ROS增加,阻断ROS产生与线粒体损伤间的恶性循环。
线粒体呼吸链复合物位于线粒体内膜上,由5个复合物组成,分别为NADH氧化还原酶(复合物I)、琥珀酸氧化还原酶(复合物II)、细胞色素还原酶(复合物III)、细胞色素C氧化酶(复合物IV)和ATP合成酶(复合物 V)[3]。线粒体呼吸链复合物 I、III是线粒体产生ROS的重要部位[3,15]。近些年来的研究表明,神经元线粒体的功能损伤影响神经元的生存及可塑性的改变,与癫痫发生及反复发作的维持密切相关[3,5-7]。本研究发现,线粒体呼吸链复合物I在癫痫发作后活性明显下降,呼吸链复合物III活性无明显变化,环孢素可明显减轻SE对神经元线粒体呼吸链复合物I活性的损害。我们以前的研究还发现,线粒体呼吸链复合物IV活性在匹罗卡品诱导的癫痫模型急性期表现为代偿性增高,而在慢性期则活性下降,而且线粒体基因组参与编码的复合物亚基表达明显下降,提示线粒体DNA损伤可能是慢性期呼吸链复合物活性降低及慢性癫痫形成的重要原因之一[3,5-6]。线粒体DNA的保护机制和修复机制均不完善,比核DNA更易受到自由基的攻击,造成线粒体DNA的降解,使每个线粒体中DNA的拷贝数下降,线粒体编码的呼吸链亚单位的合成受损,导致线粒体功能的严重破坏[13]。
总之,环孢素可减轻癫痫持续状态大鼠海马氧化应激,保护线粒体呼吸链功能。对此进行深入研究有助于我们对癫痫致神经元损伤及癫痫反复发作形成机制的理解。
[1] Osman MM,Lulic D,Glover L,et al.Cyclosporine-A as a neuroprotective agent against stroke:its translation from laboratory research to clinical application[J].Neuropeptides,2011,45(6):359-368.
[2] Martin LJ.Biology of mitochondria in neurodegenerative diseases[J].Prog Mol Biol Transl Sci,2012,107:355-415.
[3] Pieczenik SR,Neustadt J.Mitochondrial dysfunction and molecular pathways of disease[J].Exp Mol Pathol,2007,83(1):84-92.
[4] Acharya MM,Katyare SS.Structural and functional alterations in mitochondrial membrane in picrotoxin-induced epileptic rat brain[J].Exp Neurol,2005,192(1):79-88.
[5] Gao J,Chi ZF,Liu XW,et al.Mitochondrial dysfunction and ultrastructural damage in the hippocampus of pilocarpine-induced epileptic rat[J].Neurosci Lett,2007,411(2):152-157.
[6] 刘朝巍,张 涛,杨 卓.氧化应激损伤线粒体参与癫痫病理过程[J].中国病理生理杂志,2008,24(1):198-200.
[7] Han YX,Lin YT,Xu JJ,et al.Status epilepticus stimulates peroxisome proliferator-activated receptor γ coactivator 1-α/mitochondrial antioxidant system pathway by a nitricoxide-dependentmechanism [J]. Neuroscience,2011,186:128-134.
[8] Jung S,Yang H,Kim BS,et al.The immunosuppressant cyclosporin A inhibits recurrent seizures in an experimental model of temporal lobe epilepsy[J].Neurosci Lett,2012,529(2):133-138.
[9] Elrod JW,Molkentin JD.Physiologic functions of cyclophilin D and the mitochondrial permeability transition pore[J].Circ J,2013,77(5):1111-1122.
[10]王胜军,迟兆富,王树华,等.PARP调节癫痫大鼠海马核转录因子κB及相关炎症因子的研究[J].中国病理生理杂志,2010,26(1):86-91.
[11] Chi LY,Wang SJ,Li XG.Poly(ADP-ribose)signal in seizures-induced neuron death[J].Med Hypotheses,2008,71(2):283-285.
[12]Barros DO,Xavier SM,Barbosa CO,et al.Effects of the vitamin E in catalase activities in hippocampus after status epilepticus induced by pilocarpine in Wistar rats[J].Neurosci Lett,2007,416(3):227-230.
[13] Liang LP,Patel M.Mitochondrial oxidative stress and increased seizure susceptibility in Sod2-/+mice[J].Free Radic Biol Med,2004,36(5):542-554.
[14]曹丽丽,张同霞,王胜军,等.抗坏血酸对癫痫大鼠海马分子伴侣介导的自噬激活的影响[J].中国病理生理杂志,2012,28(6):1140-1143.
[15] St-Pierre J,Buckingham JA,Roebuck SJ,et al.Topology of superoxide production from different sites in the mitochondrial electron transport chain[J].J Biol Chem,2002,277(47):44784-44790.
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
中医眼耳鼻喉杂志(2021年1期)2021-07-22
医药前沿(2020年9期)2020-07-04
创新作文(小学版)(2019年10期)2019-09-25
商品与质量(2018年52期)2018-12-06
中成药(2018年7期)2018-08-04
中成药(2018年3期)2018-05-07
中国医药指南(2017年3期)2017-11-13
中成药(2017年5期)2017-06-13
植物营养与肥料学报(2012年5期)2012-10-26
