
苜蓿银纹夜蛾核型多角体病毒Gp64蛋白的优化表达
2014-10-29 12:20徐智鹏张佑红张楠翟莉莉黄双成陈杏洲
湖北大学学报(自然科学版) 2014年1期
徐智鹏,张佑红,张楠,翟莉莉,黄双成,陈杏洲
(武汉工程大学化工与制药学院,绿色化工过程教育部重点实验室,湖北 武 汉430074)
0 引言
苜蓿银纹夜蛾核型多角体病毒是杆状病毒科核型多角体病毒属的典型代表,能感染30多种鳞翅目昆虫,该病毒的基因组是大小为134kb的环状单一双链DNA分子,编码约90种蛋白质.由于昆虫杆状病毒DNA具有大量的非复制必需区,能容许基因缺失或替换,具有较大的可操作性,能够容纳大片段外源DNA的插入,因此可以用作外源基因的表达载体[1-3].感染宿主后的核型多角体病毒有两种表型:一种为包涵体病毒(OV),另一种则为非包涵体病毒(BV)[4-5].目前只在BV中发现Gp64蛋白,其集中在杆状病毒BV颗粒的一端,是第三类过滤性的膜融合蛋白,病毒感染的早期和晚期均能表达这个蛋白,是病毒通过吸附、包吞作用进入宿主细胞的关键功能蛋白之一[6-7],它在出芽病毒吸附内吞进入细胞的过程中作pH依赖型的膜融合蛋白,通过与胞饮体膜融合而使病毒粒子进入到细胞内.最近的研究也报道了这个蛋白可以促进杆状病毒进入成熟间叶细胞和胚胎干细胞等哺乳动物细胞,这意味着杆状病毒可作为有效的基因传递载体用于基因治疗.本研究克隆了来自苜蓿银纹核型多角体病毒的Gp64蛋白编码基因,将去掉了囊膜编码区的截短基因导入大肠杆菌进行原核表达,并采用不同表达条件对表达水平进行了优化.本研究的结果将为深度解析GP64基因在苜蓿银纹核型多角体病毒感染过程中的功能奠定基础[8-9].
1 材料和方法
1.1 实验材料 pET28a(+)Gp64重组质粒由本实验室自行构建,大肠杆菌BL21(DE3)由本实验室保存.
1.2 试验方法
1.2.1 重组菌的培养 挑取含有pet28a(+)Gp64重组菌的保种液在含卡那霉素(50μg/mL)的LB固体培养基上划线,培养12h后挑取单菌落接种到20mL含卡那霉素(50μg/mL)的LB培养基中,于37℃,200r/min振荡培养过夜,活化细菌.测定其OD600nm=1.529.
1.2.2 不同培养基条件下的表达 将活化好的细菌分别按照5%的接种量接种到10mL LB-kan、SOC-kan、SOB-kan和LB-kan+10%甘油液体培养基培养3h后,对重组菌进行诱导表达.测其细菌OD600nm值.收集1mL非诱导和诱导后的大肠杆菌菌液,12 000r/min离心1min后加入100μL pH8.0的PBS重悬,加入50μL上样缓冲液煮沸10min后进行SDS-PAGE.利用SDS-PAGE分析Gp64蛋白在各个培养基中的相对表达量[10].
1.2.3 不同接种量条件下的表达 将活化好的细菌分别按照1%、5%、7%、10%和15%的接种量到10mL LB-kan培养基中于37℃培养3h后,对重组菌进行诱导表达.测其细菌OD600nm值.利用SDSPAGE(电泳样品处理同前)分析Gp64蛋白在各个培养基中的相对表达量.
1.2.4 不同温度条件下的表达 将活化好的细菌接种到40mL LB-kan培养基中,在37℃下培养3h后加入相同终浓度的IPTG,等分成4份,分别放置16℃、22℃、30℃和37℃条件下进行诱导表达;同时将活化好的细菌分别接种到10mL LB-kan培养基中于22℃、30℃培养3h后加入相同终浓度的IPTG后在30℃下进行诱导表达.测其细菌OD600nm值.利用SDS-PAGE(电泳样品处理同前)分析Gp64蛋白在不同培养温度和诱导温度下的相对表达量.
1.2.5 不同诱导时间条件下的表达 将活化好的细菌按10%接种到60mL LB-kan培养基中,在37℃下培养3h后入IPTG至终浓度为0.3mmol/L后均分6份.分别在30℃诱导表达1h、3h、5h、7h、9h和12h.测其细菌OD600nm值.利用SDS-PAGE(电泳样品处理同前)分析Gp64蛋白在不同诱导时间下的相对表达量[11].
1.2.6 不同诱导剂浓度条件下的表达 将活化好的细菌按10%接种到60mL LB-kan培养基中,培养3h后,均分为6份.分别加入IPTG 至终浓度分别为0.1mmol/L、0.3mmol/L、0.5mmol/L、0.7mmol/L、0.9mmol/L、1.2mmol/L后进行诱导表达.测其细菌OD600nm值.利用SDS-PAGE(电泳样品处理同前)相对定量分析Gp64蛋白在不同终浓度诱导剂的条件下的表达量[12].
2 结果和分析
2.1 培养基对重组菌生长和蛋白表达的影响 根据实验方案分别在不同培养基中培养3h后和诱导5h后测OD600nm发现,使用LB培养基能得到最大的生物量.如表1所示,最大的菌体OD600nm值为2.459.根据表1分析可以得出LB培养基最适合该重组菌的生长繁殖.
SDS-PAGE电泳图(图1)显示使用LB培养基能够得到最高的蛋白表达量.
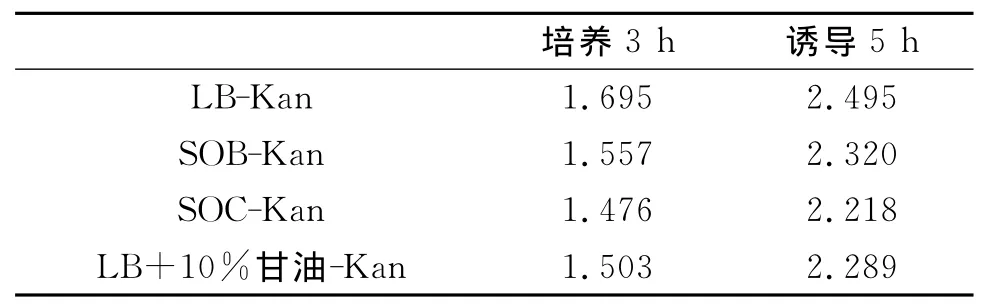
表1 不同培养基在培养3h和诱导5h后的OD600nm值
2.2 接种量对重组菌生长和蛋白表达的影响 按照实验方案分别使用不同接种量,在培养3h和诱导5h后测OD600nm发现并不是接种量越高,测的OD600nm值越大.如表2所示,该表说明了当接种量为10%的时候在培养3h和诱导5h后能得到最大的细菌生物量,最大OD600nm值为2.578.不论是培养过程还是诱导过程,培养基的成分对细菌生长都存在一个极限值,在培养相同时间下,达到极限值之前随着接种量的增大而增大,达到极限值后就会随着接种量的增大而变小.

图1 培养基对Gp64蛋白表达的影响
OD600nm和电泳图片(图2)显示在接种量为10%时候蛋白表达量最高.分析结果可得,并不是接种量越多生成目的蛋白越多,因为接种量相对过多时,细菌之间存在生长竞争,反而会抑制其代谢,使目标蛋白产率降低.
2.3 温度对重组菌生长和蛋白表达的影响 在不同培养温
度下培养3h后测细菌菌液OD600nm值发现在37℃的时候有最大值1.621.在不同的温度下诱导5h后测OD600nm发现,在30℃诱导5h后得到最大值2.598.SDS-PAGE电泳图(图3)显示在37℃条件下培养,30℃诱导的条件下表达的蛋白量最高.分析可得到重组细菌的最佳生长繁殖温度为37℃,但最佳表达温度为30℃.

表2 不同接种量下培养3h和诱导5h后的OD600nm
2.4 不同诱导时间对蛋白表达的影响 在培养相同时间后,得到相同的菌液量.分别加入终浓度为0.3mmol/L IPTG后经不同诱导时间诱导后测定OD600nm值.图4显示了在12h内细菌生物量都在随诱导时间的增加而增加,12h后基细菌量维持在一个量左右且在慢慢减小,但在培养5h时细菌生殖速率最快.SDS-PAGE电泳图片(图5)显示在30℃诱导5h表达量最高.当培养5h左右,菌体增长速率最快,此时菌体代谢最旺盛,蛋白的表达效率最高.
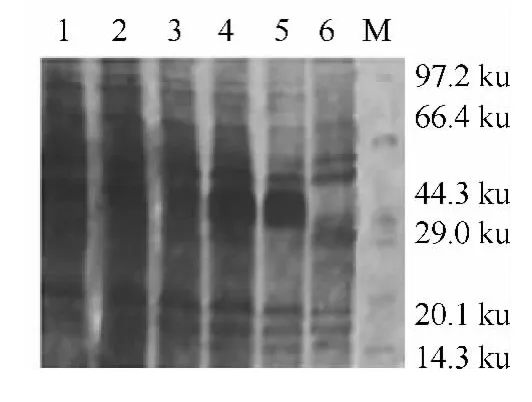
图2 接种量对Gp64蛋白表达的影响
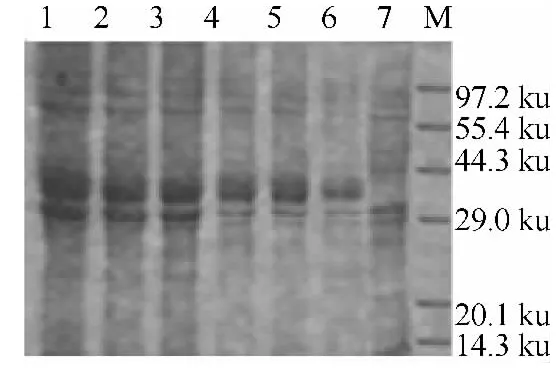
图3 温度对Gp64蛋白表达的影响
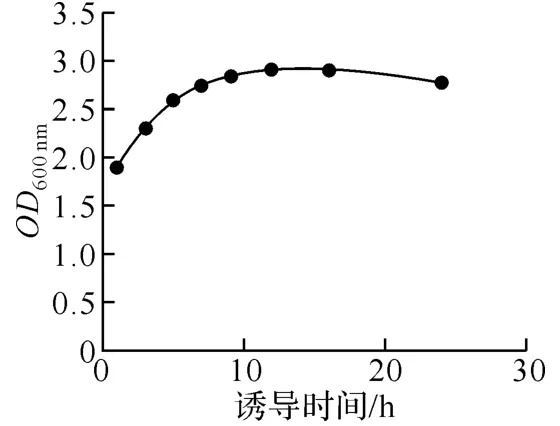
图4 不同诱导时间下OD600nm值
2.5 不同诱导剂浓度对蛋白表达的影响 在按上述实验方法操作后,分别加入不同体积的IPTG诱导表达.测定其细菌OD600nm值如图6所示,该图显示在IPTG为0.3mmol/L左右的时候有最大的生物量.当IPTG的浓度过低或过高都会抑制细菌的生长繁殖.分析细菌OD600nm和观察电泳图片(图7)发现在诱导剂IPTG浓度为0.3mmol/L的时候蛋白的表达量最大.
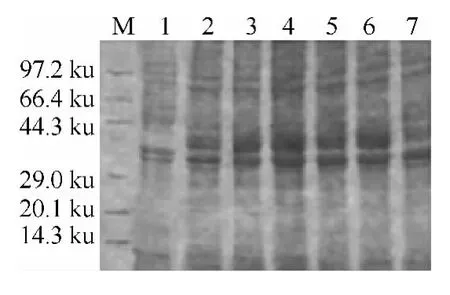
图5 诱导时间对Gp64蛋白表达的影响
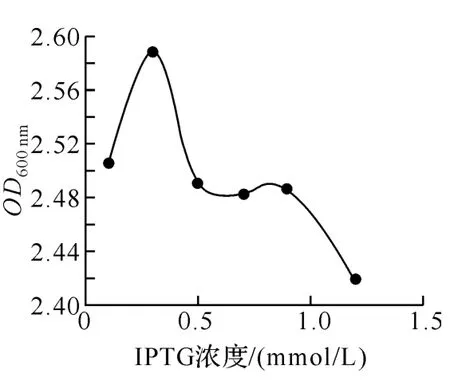
图6 不同的诱导剂浓度下的OD600nm

图7 IPTG浓度对Gp64蛋白表达的影响
3 讨论
利用基因工程技术在E.coli中高水平表达外源基因从而获得大量异源蛋白,是现代分子生物学的巨大成就之一.大肠杆菌作为基因工程表达的首选宿主,具有生产周期短、成本低、产量高等优点.但外源基因在表达过程中,往往会遇到表达效率与产率低等困难.影响外源基因在大肠杆菌中表达的因素很多.本研究就培养基、pH、接种量、诱导温度、诱导时间和诱导剂浓度进行优化,最终确定了大肠杆菌表达Gp64蛋白的最佳条件为:pH为自然条件下的LB培养基,37℃培养3h,30℃诱导表达5h,IPTG终浓度为0.3mmol/L.在此之前构建全长Gp64基因表达Gp64蛋白,尽管通过测序检测该序列的正确性,但是依旧无法正常诱导表达全长Gp64蛋白,可能是因为其跨膜区域的存在,影响了表达蛋白的正常折叠,而无法形成正常的蛋白.同时进行稀有密码子分析发现,在其跨膜区的位置稀有密码子大量出现,可能导致蛋白翻译在此无法正常进行,进而无法正常编码蛋白质[13-14].所以在切掉全长Gp64基因的跨膜区,重新构建重组子,表达截短Gp64蛋白.表达Gp64蛋白为研究Gp64蛋白及其受体在病毒感染过程中的定位、分布及其功能研究奠定了工作基础.Gp64蛋白截短后改变了蛋白的部分结构,部分功能也发生了改变,但得到的重组截短Gp64蛋白仍然保留其免疫原性,可以通过制备抗体或蛋白质体外相互作用等来开展进一步的研究,对后续的研究工作或结果不会产生较大的负面影响.
[1]Kost T A,Condreay J P,Jarvis D L.Baculovirus as versatile vectors for protein expression in insect and mammalian cells[J].Nature Biotechnology,2005,23(5):567-575.
[2]Chejanovsky N.Using the baculovirus insect cell system to study apoptosis[J].Baculovirus and Insect Cell Expression Protocols,2007(388):435-446.
[3]Stewart M E,Bonne N.Baculovirus expression of beak and feather disease virus(BFDV)capsid protein capable of self-assembly and haemagglutination[J].Journal of Virological Methods,2007,141(2):181-187.
[4]Wang S,Fang L.Construction and immunogenicity of pseudotype baculovirus expressing GP5and M protein of porcine reproductive and respiratory syndro-me virus[J].Vaccine,2007,25(49):8220-8227.
[5]Cox M M.Progress on baculovirus-derived influenza vaccines[J].Current Opinion in Molecular Therapeutics,2008,10(1):56-61.
[6]Jan Kadlec,Silvia L,Nicola G A.The postfusion structure of baculovirus gp64supports a unified view of viral fusion machines[J].Nature,2008(10):1024-1030.
[7]Li Z F,Gary W.Functional analysis of the transmembrane(TM)domain of the autographa californica multicapsid nucleopolyhedrovirus GP64protein:substitution of heterologous TM domains[J].Journal of Virology,2008(8):3329-3341.
[8]Li Z F,Gary W.Baculovirus GP64disulfide bonds:the intermolecular disulfide bond of autographa californica multicapsid nucleopolyhedrovirus GP64is not essential for membrane fusion and virion budding[J].Journal of Virology,2010(8):8584-8595.
[9]金晶,蔡亦红,类延花.HCMV pp65截短蛋白原核表达条件优化[J].微生物学杂,2005,25(3):28-32.
[10]苏友禄,冯娟,孙秀秀.赤点石斑鱼神经坏死病毒 MCP基因原核表达条件优化[J].农业科学与技术,2008,9(6):59-63.
[11]陈胜男,马素贞,翟少伦.犬细小病毒VP2基因原核表达条件优化与蛋白纯化[J].新疆农业大学学报,2011,34(1):59-61.
[12]潘滨,吴建,李桂新.烟草曲茎病毒复制相关蛋白基因原核表达条件优化[J].浙江大学学报:农业与生命科学版,2007,33(1):24-28.
[13]李国辉,唐琦,胡朝阳.家蚕核型多角体病毒囊膜蛋白 GP64的原核表达[J].微生物学杂志,2011,31(4):90-94.
[14]Lindley KM,Su JL,Paula K.Production of monoclonal antibodies using recombinant baculovirus displaying gp64-fusion proteins[J].Journal of Immunological Methods,2000,234(2):123-135.
猜你喜欢
江苏农业科学(2022年2期)2022-02-15
宁夏医学杂志(2020年3期)2021-01-21
江西农业大学学报(2018年5期)2018-11-22
上海医药(2016年23期)2016-12-22
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
广西林业科学(2016年2期)2016-03-20
现代农业(2016年6期)2016-02-28
中国医药生物技术(2015年4期)2015-12-26
哈尔滨医药(2015年2期)2015-12-01
中国当代医药(2015年9期)2015-03-01
