
外来入侵植物胜红蓟种群构件生物量结构特征
2014-10-28 02:53闫小红
井冈山大学学报(自然科学版) 2014年3期
闫小红,周 兵,王 宁
外来入侵植物胜红蓟种群构件生物量结构特征
*闫小红,周 兵,王 宁
(井冈山大学生命科学学院,江西,吉安 343009)
胜红蓟是广泛分布于我国长江流域及以南地区的主要外来入侵植物之一。通过样方调查,从构件水平研究了胜红蓟各构件的结构特征及其生物量间的关系模型,并进行了定量分析,探讨了其与入侵性的关系。结果表明,胜红蓟种群各构件生物量之间的关系为茎(3.62 g)>叶(2.18 g)>根(1.36 g)>花序(0.60 g),总生物量平均值为7.76 g,植株高度平均值为61.62 cm,平均花序数为79.15个。各构件生物量所占比率表现为茎(47.11%)>叶(29.04%)>根(17.54%)>花序(6.31%)。胜红蓟种群各构件生物量之间呈显著的正相关关系,均可用线性函数模型较好地表达。胜红蓟种群各构件较强的表型可塑性极大地提高了其入侵能力,而在繁殖器官相对较低的生物量分配显示了在其花期开展防控的针对性和实效性。
外来入侵植物;胜红蓟;生物量结构
生物入侵导致入侵地原有生物地理分布和生态系统的结构与功能发生改变,生物多样性急剧下降,影响了农业生产和人类健康,造成了巨大经济损失[1-3],因此备受世界关注,成为当今世界生态学研究的热点问题之一。一般认为,成功的入侵植物常具有较强的适应性、繁殖力和散布力[4-5]。胜红蓟(L.)是菊科(Asteraceae)藿香蓟属(sp.)一年生草本植物,原产中南美洲、墨西哥一带,自19世纪通过人工引种进入中国之后,迅速扩散,现广泛分布于我国长江流域及以南地区[6]。在低山、丘陵及平原地带的农田、路旁、荒地、桔园、茶园等普遍生长[7],严重影响了作物的生长和发育,造成减产[8-12]。由于其较强的化感抑制效应[13-15]、光竞争能力[16]和种子繁殖能力[17-18]等特性,胜红蓟具有较强的入侵性,在入侵地常形成优势群落,抑制其它植物的生长,严重降低了本地生态系统的生物多样性。同时,胜红蓟还是多种作物病害的寄主[19-20],为病害的发生提供了条件。因此,研究其生物学特性,揭示其入侵机制,探讨其防控和利用技术成为当前胜红蓟研究的热点。而目前针对其生物学特性及防治方面的研究相对较少[21],尤其是关于其生物量分配方面的研究未见报道。为此,我们通过研究胜红蓟种群个体构件的生物量分配状况及它们之间的关系,探讨其生物量分配特征与入侵性的关系,为揭示胜红蓟的入侵机制,预防其继续扩散,减少其危害提供理论基础。
1 研究地区与研究方法
1.1 样地概况
研究区设在江西省吉安市青原区,该地处吉安市东南部,属赣江中游,罗霄山脉中段,25°58′32″~27°57′50″ N,113°46′~115°56′ E。属亚热带季风湿润性气候,气候温和、光照充足、雨量充沛,年平均气温为17.1 ~ 18.6 ℃,年平均降水量为1487 mm。具有四季分明、雨热同季、无霜期长等气候特点。研究样地胜红蓟占绝对优势,覆盖度达90%以上。
1.2 研究方法
1.2.1 取样
在全面踏查的基础上,选择人为干扰较小且具有代表性的胜红蓟种群分布地段,随机设立15个样方,样方面积为1 m × 1 m,样方间距离不小于3 m。调查每个样方内胜红蓟种群个体数量,并随机选取10株大小不等的个体,15个样方共150株。测量各植株的高度,将植株连根系全部挖出,一起带回实验室,用水浸泡去掉根部泥土,冲洗干净后将植株的根、茎、叶、花等器官分离,分别装袋,在70 ℃下烘干至恒重,用电子天平逐株逐项称重。
1.2.2 数据处理
采用Excel和SPSS 19.0进行相关数据统计分析。对各构件生物量之间的关系建立定量化描述模型[22-24],所有定量关系均进行直线函数= a + b,幂函数= ab,指数函数= aebx模型的回归分析,相关性最高者为其最佳描述模型。
2 结果与分析
2.1 胜红蓟种群各构件生物量特征
胜红蓟种群各构件生物量特征见表1。
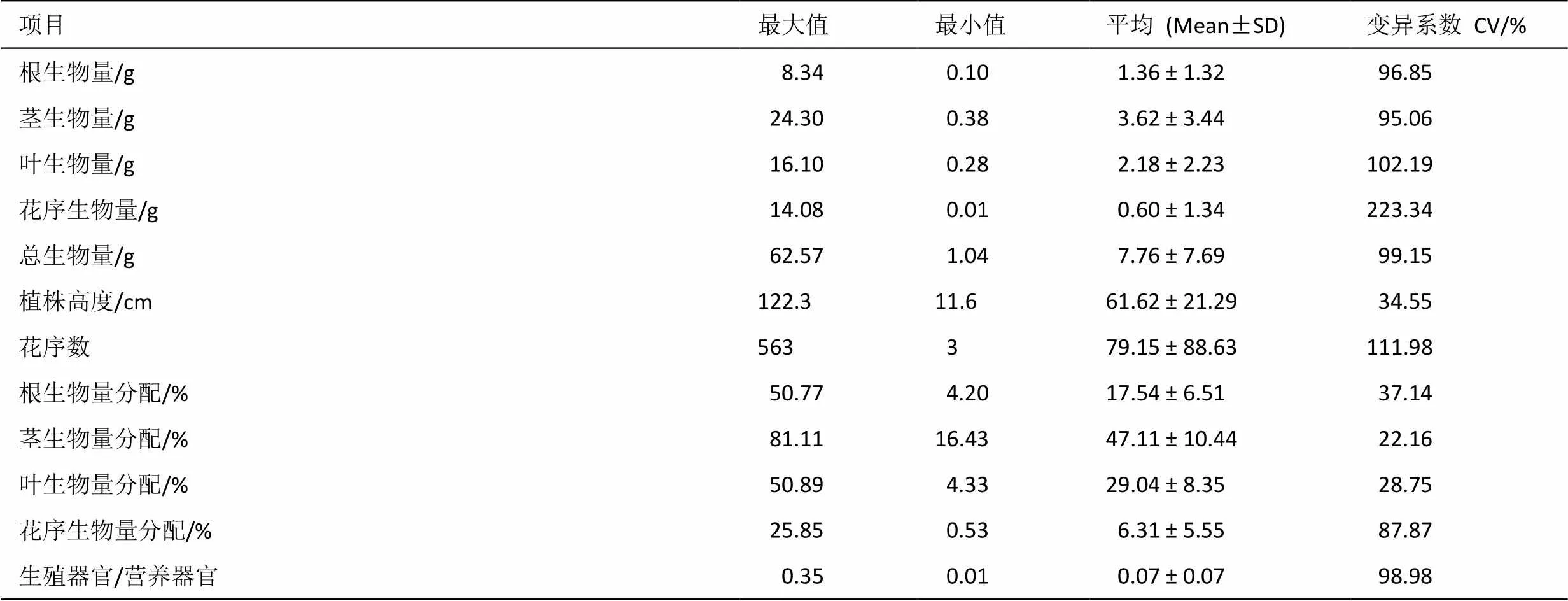
表1 胜红蓟种群各构件生物量特征
结果可见,各构件生物量之间关系表现为茎(3.62 g) >叶(2.18 g) >根(1.36 g) >花序(0.60 g),相对应的变异系数分别为 95.06%、102.19%、96.85%和223.34%,总生物量、植株高度和花序数平均值分别为7.76 g、61.62 cm和79.15个,相对应的变异系数分别为99.15%、34.55%和111.98%;胜红蓟种群根、茎、叶、花序等构件生物量及总生物量最大值和最小值之间的比值分别为83.4、63.95、57.50、1408.00和60.16,表明胜红蓟种群各功能构件具有较大表型可塑性。根、茎、叶和花序生物量分配比例大小关系为茎(47.11%) >叶(29.04%) >根(17.54%) >花序(6.31%),变异系数均相对较小,说明各构件生物量分配比例具有稳定性,各构件生物量分配比率的变异系数大小表现为花序>根>叶>茎,表明茎的生物量分配比例比其它构件分配比例更稳定,其次是叶。
2.2 胜红蓟根、茎、叶和花序生物量与植株高度的关系
回归分析和相关性检验结果表明,在胜红蓟种群繁殖生长时期,营养器官茎生物量与植株高度间存在极显著正相关关系(< 0.01),根生物量与植株高度间存在显著正相关关系(< 0.05)。不同函数拟合根、茎、叶各构件与植株高度关系的拟合方程见表2,其中,线性函数模型能较好的拟合各构件生物量与株高间的关系,相对应的函数方程分别为y = 3.114x + 57.36、y = 2.383x + 53.00和y = 1.050x + 59.33,相关系数分别为0.192、0.385和0.110,幂函数能较好的拟合花序生物量与株高间的关系,相对应的函数方程为y = 54.71x-0.03。植株高度随根、茎和叶生物量增加而升高,随花序生物量增加而降低;反映制约作用规律性的值分别为3.114、2.383、1.050和-0.03,表明根、茎、叶和花序生物量对植株高度的影响制约程度表现为根>茎>叶>花序(图1)。

表2 胜红蓟植株高度与各构件生物量间关系模型

图1 胜红蓟植株高度与各构件生物量间的关系
2.3 胜红蓟种群各构件生物量间的关系
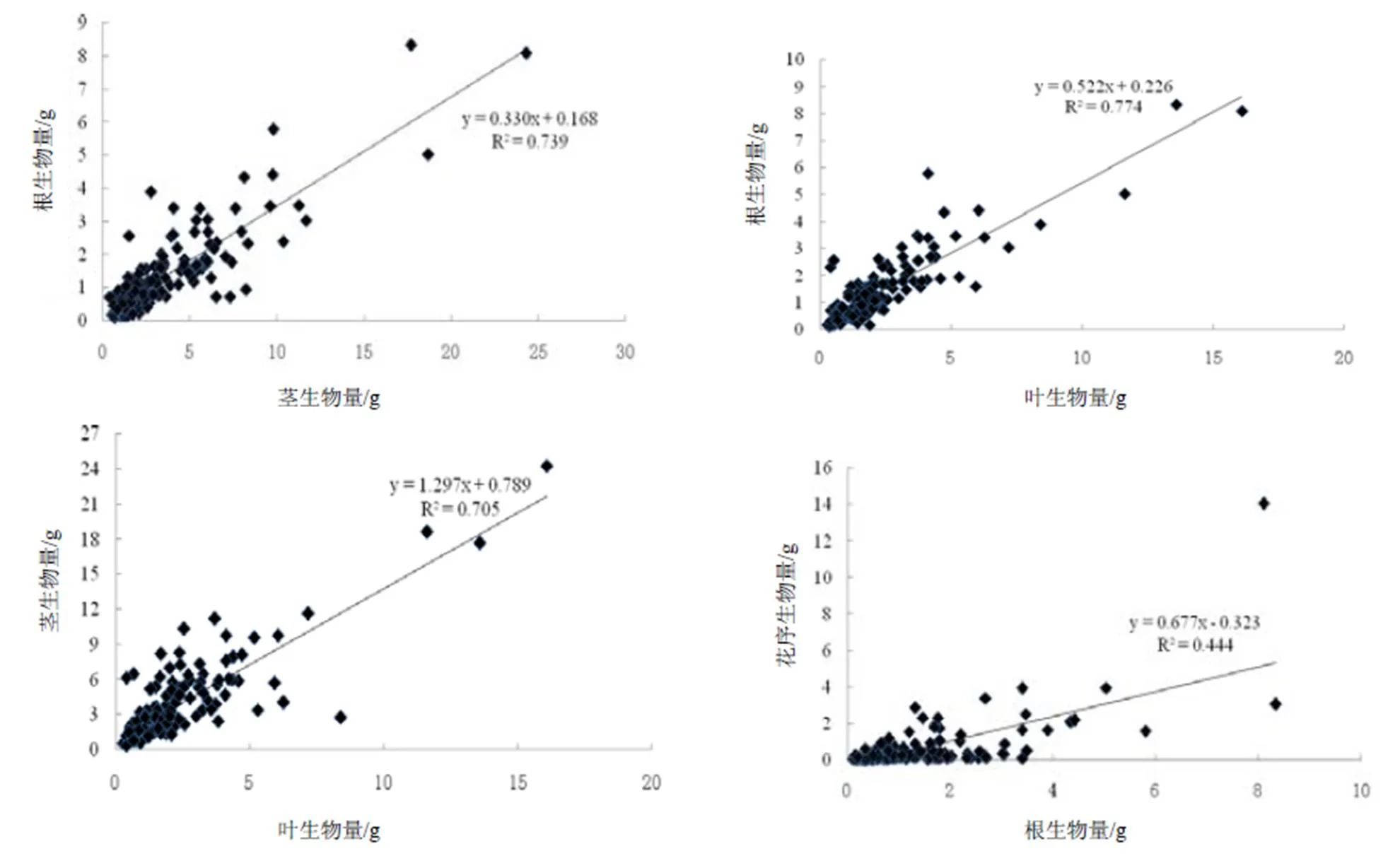
回归分析和相关性检验结果表明,胜红蓟种群根、茎、叶和花序各构件生物量间存在着极显著的正相关关系(< 0.01),不同函数的拟合方程见表3,其中,线性函数模型最能表现它们之间的关系。
胜红蓟种群营养器官之间存在着极显著正相关关系(< 0.01)。根生物量随茎生物量和叶生物量,茎生物量随叶生物量均呈线性函数形式增加,拟合曲线见图2,相对应的拟合方程分别为= 0.330+ 0.168、= 0.522+ 0.226和= 1.297+ 0.789,相关系数分别为0.860、0.880和0.840。各方程的值分别为0.330、0.522和1.297,表明茎、叶生物量对根生物量的影响制约程度表现为叶>茎。
胜红蓟种群营养器官与生殖器官花间也存在极显著的正相关关系(<0.01)。表现为花序生物量随根、茎和叶生物量均呈线性函数模型形式增加,具有较好的拟合效果(图2),相对应的拟合方程分别为= 0.677- 0.323、= 0.270- 0.375和= 0.431- 0.338,相关系数分别为0.666、0.691和0.714。各方程的值分别为0.677、0.270和0.431,表明根、茎、叶生物量对花序生物量影响制约程度表现为根>叶>茎。
胜红蓟种群不同构件生物量与同一构件生物量间,根和花序生物量随茎生物量呈线性函数增加,其值分别为0.330和0.270,表明根和花序生物量受茎生物量影响制约程度表现为根>花序;根和花序生物量随叶生物量呈线性函数增加,其值分别为0.522和0.431,表明根和花序生物量受叶生物量影响制约程度也表现为根>花序。

表3 胜红蓟各构件生物量间关系模型
3 讨论
植物种群和个体构件生物量是植物与环境因素共同作用的结果,既反映了植物种群对环境条件的适应能力和生长发育规律,也反映了环境条件对植物种群的影响程度[25-27]。本研究发现,胜红蓟种群各构件生物量之间关系表现为茎(3.62 g)>叶(2.18 g)>根(1.36 g)>花序(0.60 g),其变异系数分别为 95.06%、102.19%、96.85%和223.34%,而根、茎、叶和花序各构件生物量最大值和最小值之间的比值分别为83.4、63.95、57.50和1408.00,表明胜红蓟具有较强的表型可塑性,这增加了该种群的适应能力,有利于其适应新的生境,增强了其入侵能力。外来入侵植物牛膝菊()[22]、小飞蓬()[23]和钻形紫菀()[28]在生物量分配方面表现出与胜红蓟相似的较强表型可塑性。
在生物量分配比例方面,胜红蓟种群各构件表现为茎(47.11%)>叶(29.04%)>根(17.54%)>花序(6.31%),相对应的变异系数表现为茎(22.16)<叶(28.75)<根(37.14)<花序(87.87),表明胜红蓟在茎和叶上投入的资源比较多,且比较稳定。在茎和叶上面的大量资源投入有利于胜红蓟尽可能多的获得生存空间和光资源,有利于其营养物质的积累,保证了其营养生长和有性生殖的营养需求。虽然胜红蓟在花序上的生物量分配比例较低,但胜红蓟以种子繁殖为主,同时具有较强的营养繁殖特性,在节上可产生不定根,扦插也能生根成活,且种子萌发率极高,刚成熟的种子萌发率可达70%以上[21]。在调查的样方中,同时出现开花结籽的成熟植株和新生的幼苗,可见胜红蓟可在一个生长期内完成营养生长、开花结实、传播种子及萌发幼苗,与其它外来入侵植物小飞蓬和牛膝菊相似[22-23]。通过这种方式,胜红蓟在短期内产生大量的个体,形成优势种群,排挤其它植物,占据更广泛的生境,提高了其入侵能力。
植物生长和物质分配通常可用线性函数、幂函数或指数函数表达。研究结果显示,胜红蓟各构件生物量之间极显著正相关关系,均可用线性函数模型表达。这反映出植物生长过程中对各构件资源分配的协调性,一个构件的旺盛生长,必然有利于其它构件生物量累积,最终使整个个体增高、生物量增大[23]。根、茎生物量与株高之间呈显著或极显著正相关,而花序生物量与株高之间呈负相关,表明繁殖器官的生物量投入不利于其植株高度的增加,植株高度的增加更多的依赖于其营养器官物质的积累和供给。从模拟函数来看,根、茎、叶和株高之间均可用线性函数模拟,表明在观察期间的胜红蓟以同速生长为主,可能是因为其处于初花期,营养生长占主要优势,这与胜红蓟将大部分资源分配给营养构件是相一致的。
总之,胜红蓟各构件生物量的特征显示了其较强的入侵能力。当然,要明确这一特点,需要综合考虑胜红蓟各方面的生物学特性,比如适生特性、种子繁殖特性、传播特性以及各构件形态学和生理学特性等,这有待于深入研究。同时,胜红蓟主要依靠种子繁殖,而其在繁殖器官的生物量分配相对较少,因此,在其开花结实期进行防控一方面可以降低其结实量,另一方面可有效降低防控成本,更具针对性和实效性。
[1] Didham R K, Tylianakis J M, Hutchison M A, et al. Are invasive species the drivers of ecological change?[J]. Trends in Ecology and Evolution, 2005, 20(9): 470-474.
[2] Gurevitch J, Padilla D K. Are invasive species a major cause of extinctions?[J]. Trends in Ecology and Evolution, 2004, 19(9): 470-474.
[3] Pimentel D, McNair S, Janecka J, et al. Economic and environmental threats of alien plant, animal, and microbe invasions[J]. Agriculture, Ecosystems and Environment, 2001, 84: 1-20.
[4] 史刚荣, 马成仓. 外来植物成功入侵的生物学特征[J]. 应用生态学报, 2006, 17(4): 727-732.
[5] Pysek P, Richardson D M. Traits associated with invasiveness in alien plants: Where do we stand?[A]. In: Nentwig W, ed. Biological Invasions, Section II[C]. Berlin: Verlag-Springerc,2007.
[6] 徐海根, 强胜. 中国外来入侵物种编目[M]. 北京: 中国环境科学出版社, 2004.
[7] 郝建华, 强胜. 外来入侵性杂草——胜红蓟[J]. 杂草科学, 2005(4): 54-58.
[8] 孔垂华, 胡飞, 骆世明. 胜红蓟(L.)对作物的化感作用[J]. 中国农业科学, 1997, 30(5): 95.
[9] 陈建军, 孔垂华, 胡飞, 等. 胜红蓟化感作用研究Ⅷ. 植株对花生和相关杂草的田间化感效应[J]. 生态学报, 2002, 22(8): 1196-1201.
[10] 罗战勇, 李淑玲, 谭铭喜. 广东省烟田杂草的发生于分布现状调查[J]. 广东农业科学, 2007(5): 59-63.
[11] 赵之亭, 范志伟, 刘丽珍. 胜红蓟对大豆生长和产量的影响[J]. 热带农业科学, 2009, 29(9): 4-6.
[12] 郭艾英, 杨晴, 张风娟, 等. 三叶鬼针草和胜红蓟的叶浸提液对旱稗种子萌发的影响[J]. 河北科技师范学院学报, 2010, 24(3): 28-31.
[13] 孔垂华, 黄寿山, 胡飞.挥发油对真菌、昆虫和植物的生物活性及其化学成分[J]. 生态学报, 2001, 21(4): 584-587.
[14] Kong C H, Hu F, Xu T, et al. Allelopathic potential and chemical constituents of volatile oil from[J]. Journal of Chemical Ecology, 1999, 25: 2347-2356.
[15] Kong C H, Hu F and Xu X H. Allelopathic potential of volatiles fromunder stress[J]. Journal of Chemical Ecology, 2002, 28: 1185-1194.
[16] Banyikwa F F, Rulangorang Z K. Growth analysis of ground-nut () in competition with[J]. Turrialba, 1985, 35(3): 215-219.
[17] 林忠宁, 刘明香, 韩海东, 等. 胜红蓟种子发芽检验标准化研究[J]. 种子, 2008, 27(4): 106-108.
[18] 郝建华, 吴海荣, 强胜. 部分菊科入侵种种子(瘦果)的萌发能力和幼苗建群特性[J]. 生态环境学报, 2009, 18(5): 1851-1856.
[19] 王向阳. 广西胜红蓟黄脉病样中发现多种菜豆金色花叶病毒属的病毒[J]. 植物病理学报, 2007, 37(6): 679-682.
[20] Kashina B D, Mabagala R S, Mpunam A A. First report ofL. andBurm F. as new hosts of tomato yellow leaf curl-Tanzania virus[J]. Plant Protection Science, 2003, 39(1): 18-22.
[21] 吴海荣, 胡学难, 强胜, 等. 广州地区胜红蓟物候学观察与调查研究[J]. 杂草科学, 2010, (3): 18-21.
[22] 齐淑艳, 徐文铎, 文言. 外来入侵植物牛膝菊种群构件生物量结构[J]. 应用生态学报, 2006, 17(12): 2283-2286.
[23] 潘玉梅, 唐赛春, 蒲高忠, 等. 外来入侵植物小飞蓬种群构件生物量结构特征[J]. 广西植物, 2009, 29(4): 476-480.
[24] 郭水良, 盛海燕. 北美车前种群密度制约的统计分析[J]. 植物研究, 2002, 22(2): 236-240.
[25] 姜立志, 王东, 刘树楠, 等. 光照和氮素对喜旱莲子草形态特征和生物量分配的影响[J]. 水生生物学报, 2010, 34(1): 101-107.
[26] 王伯荪, 李鸣光, 彭少麟. 植物种群学[M]. 广州: 广东高等教育出版社, 1995: 8-27.
[27] Gurevitch J, Howard T G, Ashton I W, et al. Effects of experimental manipulation of light and nutrients on establishment of seedlings of native and invasive woody species in Long Island, NY forests[J]. Biological Invasions, 2008, 10: 821-831.
[28] 潘玉梅, 唐赛春, 岑艳喜, 等. 钻形紫菀开花期种群构件的生物量分配[J]. 热带亚热带植物学报, 2010, 18(2): 176-181.
MODULE BIOMASS STRUCTURE TRAITS OF THE ALIEN INVASIVE PLANT
*YAN Xiao-hong, ZHOU Bing, WANG Ning
(School of Life Sciences, Jinggangshan University, Ji’an, Jiangxi 343009, China)
is one of the alien invasive plants distributied abroadly in Yangtse river drainage area and the south area of it in China. Biomass structure characteristic and correlation models among different modules ofpopulation were studied at module level by sampling surveys. Their quantitative analysis and the relation between the characteristics and its invasiveness were also done in this paper. The results showed that the modules biomass ofwas decreased in the sequence of stem, leaf, root, capitulum, and the biomass proportion of each module to the whole plant biomass was decreased in the same sequence. The biomass of each module had significant positive correlations, and all the correlations could be well expressed by linear model. Stronger phenotypic plasticity of eachmodule greatly enhanced its invasive ability, and lower reproductive biomass allocation indicated the pertinence and efficiency for its control during flowering phase.
alien invasive plant;; biomass structure
Q948
A
10.3969/j.issn.1674-8085.2014.03.024
1674-8085(2014)03-0101-06
2014-02-09;
2014-04-12
国家自然科学基金项目(31360090);江西省自然科学基金项目(20114BAB214011);江西省教育厅科技计划项目(GJJ11717)
*闫小红(1977-),女,内蒙古赤峰人,实验师,硕士,主要从事植物生理生态学方面的研究(E-mail: yanxiaohong325@126.com);
周 兵(1977-),男,湖北黄梅人,副教授,博士,主要从事植物生态学方面的教学与研究(E-mail: zhoubing113@126.com);
王 宁(1979-),男,山东淄博人,副教授,博士,主要从事生态学方面的教学与研究(E-mail: wangning13@126.com).
猜你喜欢
今日农业(2022年15期)2022-09-20
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14
今日农业(2020年19期)2020-12-14
红土地(2018年7期)2018-09-26
南风(2017年34期)2018-01-16
科学种养(2017年6期)2017-06-13
科学种养(2017年5期)2017-05-17
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
当代畜禽养殖业(2014年10期)2014-02-27
