
燕麦microRNAs及其靶基因的生物信息学预测
2014-09-10 09:05郭红媛贾举庆张怡晴舒国平陈潇洒
山西农业科学 2014年5期
郭红媛,贾举庆,段 娜,张怡晴,舒国平,陈潇洒
(山西农业大学农学院,山西太谷030801)
microRNAs(miRNA)是在真核生物中发现的一类内源性且具有调控功能的非编码单链RNA,大小约为22个核苷酸。其主要参与发育、病毒防御、信号转导等多种调控[1-3]。成熟miRNA识别靶基因位点后,可与靶基因位点紧密结合,破坏或阻遏蛋白质翻译机制。由于序列微小,因此,一般极难检测。现在已发现大量的miRNAs,新的miRNAs也正陆续被发现。miRNA通常可以控制多个靶位点,它们可以与多达1 000种甚至更多不同的靶位点结合,有时基因也可能受到多个miRNA调控,因此,难以了解其复杂的相互作用。
目前,研究miRNA表达量和表达位点可采用分子生物学和生物信息学等方法。其中,分子生物学法可以特异性地鉴定miRNAs,但依赖于极强的技术能力,且耗时耗力,成本高[4]。随着新一代测序技术的发展,small RNA测序技术为发现miRNAs提供了强有力的手段。此外,根据miRNAs在植物物种中的高度保守性,利用比较基因组学和生物信息学方法可以从大量已测序的EST和GSS等数据库中挖掘、鉴定miRNAs,了解miRNA群或其结构信息并获取更多的细胞功能分析。
目前,利用生物信息学方法已在玉米、小麦、高粱等植物中有效地预测到大量的miRNA[5-8]。燕麦是一种粮、经、饲、药兼用的世界分布型植物,因其营养价值高而且具有抗病、耐瘠、抗旱等优良性状而引起了众多研究者的关注[9-11],公共数据库中也积累了大量的EST和GSS序列,这为研究燕麦提供了很好的可利用资源。目前,对燕麦miRNA的鉴定以及对靶基因调控方面的研究尚未见报道。
本研究根据miRNA序列的保守性,用大麦、小麦、二穗短柄草等植物miRNAs序列在燕麦EST数据库和GSS数据库中进行检索和筛选,预测得到燕麦miRNA及其靶基因,补充了燕麦miRNA数据,并为燕麦miRNA对抗旱性调控等研究方面奠定了一定的基础。
1 材料和方法
1.1 序列来源
根据序列在进化方面的保守性,采用同源性搜索方法,从miRNA数据库miRBase(http://www.mirbase.org/index.shtml)下载已登记的1 910条miRNA序列。在美国国家生物技术信息中心(NCBI)收录的燕麦EST数据库和概览序列(GSS)数据库中,将1 910条miRNA序列去重复后用来检索潜在的燕麦miRNA序列。
1.2 序列检索方法
从miRNA数据库中下载已有植物miRNA序列,用多序列比对软件ClustalW2(http://www.ebi.ac.uk/Tools/msa/clustalw2/)比对后去除重复序列;用这些序列进行BLAST(http://blast.ncbi.nlm.nih.gov/Blast.cgi)在线比对,检索NCBI中的燕麦EST和GSS数据库,将少于4个碱基错配的同源序列作为重复序列去除;再与NCBI数据库中进行BLASTX比对,去除燕麦蛋白质编码序列后,将剩余序列作为燕麦候选miRNA序列。
1.3 miRNA二级结构及靶基因的预测
将所得序列用Mfold 3.2软件(http://mfold.rit.albany.edu/?q=mfold/RNA-Folding-Form)预测其二级结构,根据对miRNA的筛选标准[12-13]进行预测、鉴定。用psRNATarget(http://plantgrn.noble.org/psRNATarget/)对符合miRNA二级结构标准的序列进行靶基因预测。
2 结果与分析
2.1 燕麦miRNAs的预测
从 miRBase下载山羊草(2)、二穗短柄草(464)、油棕(6)、高羊茅(15)、大麦(69)、水稻(713)、高粱(241)、甘蔗(36)、小麦(43)和玉米(321)已登记的miRNA序列,截至2013年10月共1 910条。对其进行多序列比对后,将重复序列只保留1条。用留下来的序列检索NCBI中的燕麦EST和GSS数据库,检索到的序列经多序列比对,重复序列保留1条。然后,将留下来的序列在燕麦的蛋白质数据库中进行BLASTX比对(以去除其中编码序列),结果共获得143条非编码的候选燕麦miRNA序列,对143条pre-miRNA的统计分析得到的参数(前体序列长度L>60 nt;GC含量为30%<CG<71%;二级结构生成自由能△G<-41.18 kJ/mol)进行进一步筛选,若符合所有条件的片段则作为候选,不符合这些条件的片段则弃之。之后,对候选序列用Mfold
3.2 进行二级结构预测并分析。
根据预测miRNA二级结构的标准,miRNA前体结构中具有茎环结构,且成熟的miRNA尽量位于茎环结构的臂上;miRNA与互补链间错配应少于5个碱基;miRNA互补链上不能有大的突起或缺口;以最小折叠自由能系数(MFEI)大于8.5来判断所预测的序列是否为符合二级结构的miRNA序列(图1)。
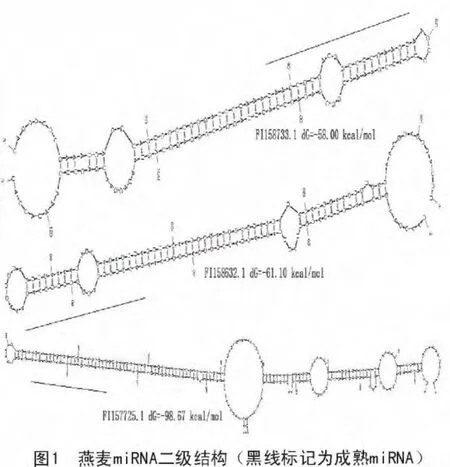
经分析、筛选,共有46条序列符合miRNA鉴定标准,表1列出了其中的10条。
2.2 燕麦miRNA靶基因的预测
经psRNATarget在线预测所得的46条符合鉴定标准的燕麦miRNA序列的靶基因,预测时选择期望值为3.0,允许不配对的最大能量(UPE)小于25.0,其余参数为默认值。结果共找到61个靶基因,部分序列的功能信息列于表2。

表1 预测到的燕麦miRNAs

表2 预测到的燕麦miRNA靶基因
3 讨论
本研究预测的燕麦miRNAs序列,首先其前体序列必须具有重要特征——发卡结构,而且预测的成熟miRNA序列要尽量位于其前体序列二级结构的“臂”上,避免处于“环”结构中;如不能避免,其与“环”结构最多有4 nt配对。为与其他类型的RNAs进行区分,又采用了2个鉴定miRNA的重要参数,即最小折叠自由能(MFE)和最小折叠自由能系数(MFEI),研究发现,MFE与前体序列的长度和GC含量有关,燕麦miRNA前体的MFEI平均值为1.06,高于 mRNA(0.62~0.66),rRNA(0.59)和 tRNA(0.64)[14],也高于前人的研究结论 0.85,结果证实,MFEI为预测miRNA的重要且可靠参数。
在预测到的靶基因中,绝大多数是编码与生长发育有关的蛋白质,参与新陈代谢,编码转录因子调控生长、发育以及其他生理代谢过程,如膜蛋白相关的物质运输、细胞信号转导等。这些靶基因分别属于不同的基因家族,部分靶基因编码的蛋白功能未知。本研究预测的miRNAs前体序列中有的具有多个成熟的miRNA序列,前体序列长度为107~315 nt。预测的成熟miRNAs的位置有的位于3'端,有的位于5'端。所预测的所有miRNA前体序列在长度和二级结构方面虽各不相同,但都分别可折叠成标准的miRNA二级结构。在预测的燕麦miRNA序列中大多是编码与生长发育相关的蛋白质,可能参与燕麦生长中的新陈代谢活动,有一些是与信号传导相关的,还有一些可能是编码转录因子的。但由于目前燕麦基因组信息在网上公布信息的不完全,本研究对部分预测的miRNA序列仍未能预测到其功能。目前,有许多软件可对RNA功能进行预测,如:TargetScan(http://www.targetscan.org/),PicTar(http://pictar.mdc-berlin.de/),MiRscan(http://genes.mit.edu/mirscan/)等。想要得到准确的预测结果,可以用多种软件反复预测、比较,但预测结果最终需经实验进一步验证。
miRNAs作为一种非编码的基因表达调控因子,虽已有在小麦、水稻、玉米、棉花、花生、烟草等[15-17]方面的研究,但目前对燕麦miRNAs生物学功能及调控机理的研究还不多。本研究预测了燕麦miRNAs序列及其靶基因,为进一步研究燕麦miRNAs的功能奠定了基础(可为开展具体的实验验证提供依据,从而对燕麦miRNAs生物学功能注释缩小范围)。相信随着生物信息学的发展,会有更多、更全面的基因组序列公布于网上,会发现更多的燕麦miRNAs。
[1]段成国,王春晗,郭惠珊.MicroRNA对植物生长发育和病毒侵染的调控[J].科学通报,2006,51(4):371-375.
[2]郭晓琴,孙红正.microRNA靶基因检测技术研究进展[J].河南农业科学,2013,42(4):11-13.
[3]耿锐梅,杨宏伟,曹长代,等.植物MicroRNA研究进展及其在植物与病原菌互作中的作用 [J].浙江农业科学,2011(5):1092-1094.
[4]苏小茜,张清政,姜枫,等.MicroRNA研究方法进展[J].上海畜牧兽医通讯,2008(5):2-4.
[5]李婧,熊莉丽,胡久梅,等.基于EST和GSS序列的玉米未知微RNA的数据挖掘[J].生物技术通报,2011(12):108-112.
[6]韩友生,栾福磊,朱洪亮,等.生物信息学方法挖掘小麦microRNAs及其靶基因[J].中国科学,2009,39(6):585-595.
[7]杜江峰,武永军,方晓峰,等.生物信息方法预测高粱miRNA及其靶基因[J].科学通报,2010,55(7):553-561.
[8]蔡斌,李成慧,彭日荷,等.葡萄microRNA的计算识别[J].华北农学报,2008,23(S2):213-216.
[9]周建萍,刘龙龙,崔林.山西省燕麦育种现状及资源特点[J].山西农业科学,2010,38(11):6-9.
[10]刘龙龙,崔林,刘根科,等.山西省燕麦产业现状及技术发展需求[J].山西农业科学,2010,38(8):3-5.
[11]刘龙龙,崔林,周建萍,等.利用核不育燕麦新种质选育新品种品燕 2号[J].山西农业科学,2012,40(5):445-446.
[12]Zuker M.Mfold web server for nucleic acid folding and hybridization prediction[J].Nucleic Acids Res,2003,31(13):3406-3415.
[13]张磊,曹江涛,崔萌萌.茄子microRNAs与其靶基因的生物信息学预测[J].遗传,2011,33(7):776-784.
[14]Zhang B H,Pan X P,Cox B,et al.Evidence that miRNAs are different from other RNAs[J].Cell Mol Life Sci,2006,63(2):246-254.
[15]潘玉欣,刘恒蔚.花生miRNA与其靶基因的生物信息学预测[J].中国油粮作物学报,2010,32(2):290-294.
[16]金伟波,李楠楠,吴方丽,等.水稻MicroRNA的预测及实验验证[J].中国生物化学与分子生物学报,2007,23(9):743-750.
[17]Frazier TP,Xie FL,Freistaedter A,et al.Identification and characterization of microRNAsand their target genesin bobacco(Nicotiana tabacum)[J].Planta,2010,232(6):1289-1308.
猜你喜欢
中老年保健(2022年5期)2022-11-25
天津医科大学学报(2021年1期)2021-12-05
现代畜牧科技(2021年10期)2021-11-19
今日农业(2021年7期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
天津医科大学学报(2019年3期)2019-08-13
中国博物馆(2018年2期)2018-12-05
延河(2017年7期)2017-07-19
现代检验医学杂志(2016年5期)2016-08-20
现代检验医学杂志(2015年4期)2015-02-06

- 山西农业科学的其它文章
- 一个新的小麦-中间偃麦草异代换系的分子细胞学鉴定