
木本果树开花相关基因的研究进展
2014-08-29 08:57魏丹凤曾黎辉
亚热带农业研究 2014年2期
魏丹凤,曾黎辉
(福建农林大学园艺学院,福建 福州 350002)
木本果树开花相关基因的研究进展
魏丹凤,曾黎辉
(福建农林大学园艺学院,福建 福州 350002)
开花是植物从营养生长向生殖生长的转变过程。拟南芥中存在多条调控开花时间的信号途径如光周期途径、春化途径、赤霉素途径和自动途径等。与草本植物相比,木本植物成花调控研究进展较缓慢。本文综述了木本果树中开花综合基因,即综合不同成花调控途径信号的关键基因,如CO、FLC、SVP、FT、LFY、TFL、AP1、SOC1等的研究进展,以期为进一步探索木本植物成花的分子调控机制提供参考。
木本果树; 开花途径; 开花综合基因
木本多年生植物成花调控包括童期调控和成年期花芽的诱导。木本果树实生苗一般需要经历5-10年的童期之后,才具备开花能力;成年树开花诱导具有周期性,这些因素制约着果树及林木的遗传改良。植物成花过程由外部环境因子和内部生长发育状态共同决定,对果树开花的研究主要是借鉴拟南芥等模式植物的研究成果。拟南芥花芽分化主要受光周期途径、春化途径、赤霉素(GA)途径和自主开花途径调节(图1),以及最近提出的年龄相关(age-related)途径[1,2]。这些成花途径相互作用,有多个基因参与影响植物开花,其中FLOWERINGLOCUST(FT)、SUPPRESSOROFOVEREXPRESSIONOFCONSTANS1(SOC1)、APETALA1(AP1)和LEAFY(LFY)等综合基因(integrator gene)综合多条诱导途径信号,决定确切的开花时间;TERMINALFLOWER1(TFL1)、FLOWERINGLOCUSC(FLC)和SHORTVEGETATIVEPHASE(SVP)是成花调控网络的抑制基因,也是连接不同调控途径的综合基因。以上基因在拟南芥、水稻以及其他高等植物之间具有很高的保守性。目前,已将苹果FT基因[3]和龙眼FT、AP1基因[4]转入拟南芥中表达影响开花;而拟南芥FT、LFY、AP1基因同样可以调控柑橘的成花转变[5];此外,拟南芥基因在苹果[6](LFY)、杨树[7,8](FT/LFY)、梨[9](FT)、枳橙[10](AP1/LFY)的表达也已获得成功。但是木本植物具有童期、开花季节性等特殊的成花特性说明其可能有与草本植物不同的开花调控途径和作用网络。
本文侧重综述了木本果树中几个综合基因的研究进展,以期为进一步探索木本植物成花的分子调控机制提供参考,为人工调控果树花芽分化和缩短童期提供新方法。
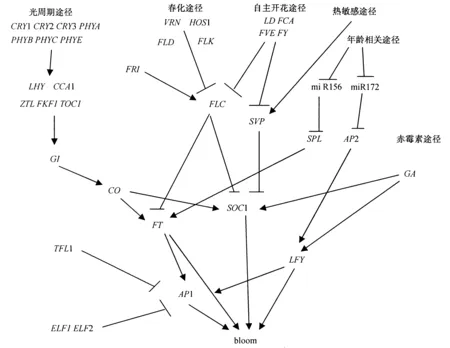
箭头表示促进;横线表示抑制。
1 CO基因
拟南芥中光周期途径通过激活开花整合基因FT、SOC1促进植物的开花。CONSTANS(CO)是光周期途径中的关键基因[11],可以整合光信号和生物钟信号,节律性地激活下游基因FT的表达从而诱导开花。CO的过量表达促进拟南芥在任何光周期下提早开花。CO基因位于GIGANTEA(GI)基因的下游、FT和SOC1的上游。短日照通常使拟南芥CO基因表达下降导致晚花,长日照下CO基因表达上升可以诱发FT表达和开花。目前已知的昼夜节律类基因如EARLYFLOWERING3 (ELF3)、FLAVINBINDINGKELCH-REPEATF-BOX1 (FKF1)、TIMINGOFCABEXPRESSION1 (TOC1)、ZEITLUPE(ZTL)、LUXARRHYTHMO(LUX)和LOVKELCHPROTEIN2 (LKP2)均能激活CO的表达。ELF3基因介导GI基因在光下的节律表达,GI和FKF1可通过影响CDF1调控CO表达,CDF1(CYCLINGDOFFACTOR1)蛋白通过结合到CO基因启动子上抑制CO表达。长日照下GI代替CO与CDF1结合,FKF1使CDF1蛋白降解,故CO表达上升[12]。另一个CO转录抑制因子CDF2也被FKF1作为目标减弱表达。除CDF蛋白外,还有主要在长日照下影响CO、FT表达的抑制因子REDANDFAR-REDINSENSITIVE2(RFI2)及短日照下抑制CO表达的DAYNEUTRALFLOWERING(DNF)[13]。近期从葡萄中分离出CO的同源基因VvCOL1、VvCO[14]。VvCOL1可能参与芽休眠诱导与维持的光周期转录调控。VvCO在潜伏芽的作用途径表明葡萄可能与拟南芥具有相似的光周期调控路径[14]。然而,富士苹果中CO同源基因MdCOL1、MdCOL2的表达模式异于拟南芥,表现出器官特异性,对生殖器官的生长发育有重要作用[15]。
2 FLC和SVP基因
温度对植物成花诱导具有重要作用,与温度有关的开花途径主要是春化(低温)作用。植物春化特性由多基因控制,且基因之间存在复杂的作用关系。在拟南芥中,与春化作用相关的基因主要有VERNALIZATION1(VRN1)、VERNALIZATION2(VRN2)、VERNALIZATIONINDEPENDENT3(VRN3)、FRIGIDA(FRI)和FLC。目前拟南芥FLC的表达机制较清楚,而十字花科之外的研究很少。FLC基因对成花具有抑制作用,表达水平越高,开花时间则越晚。在春化途径中,VRN类基因和HIGHEXPRESSIONOFOSMOTICALLYRESPONSIVEGENES1 (HOS1)基因能抑制FLC基因的表达。VRN1、VRN2可以维持对FLC的抑制,VIN3只在低温时表达,并且随低温处理时间的增加表达水平升高。FRI基因参与FLC的表达而延迟植物开花,FRI的存在可增强FLC的抑制作用,但自动途径的FCA基因可以逆转FRI对FLC的促进作用。各个途径通过FLC基因相互联系,自主途径可能与春化途径共同通过染色体甲基化、乙酰化等方式控制FLC的表达。与野生型相比拟南芥的fca、fy、fpa、ld、fld、flk、fve突变体能提高FLC基因的表达水平,在长日照和短日照下均表现为晚花。LD与FCA、FPA、FVE、FY、FPA、FLK等基因可能通过转录后修饰和染色体组蛋白修饰抑制FLC基因表达,共同促进开花[16]。此外,年龄相关途径则通过miR156、miR172及其靶基因SQUAMOSAPROMOTERBINDINGLIKE(SPL)、APETALA2 (AP2)的表达响应低温春化调控开花时间[1,2]。
FLC基因通过春化和自动途径调控植物开花已经在枳的研究中得到证明[17]。PtFLCmRNA在普通枳和早实枳中存在不同的选择性剪切本,这些剪切本在普通枳和早实枳的各个发育阶段有不同的表达量,由此推测枳由童期向成年树的转变可能与PtFLC的特异性剪切有关[18]。将枳PtFLC不同剪切序列构建表达载体在早实枳及椪柑中超量表达,结果表明PtSOC1严格响应PtFLC的负调控[19]。在拟南芥中,ELF5调控FLC的表达。研究发现,童期枳PtELF5的表达水平高于PtFLC,成年树二者的表达受季节和花发育调控,推测PtELF5是在枳早花中发挥重要作用的基因[20,21]。最近研究发现了FLC基因在植物中影响开花以外的功能。从山核桃花芽中鉴定到2个FLC的同源基因。与1、2年生植物相比,该基因可能在山核桃中发生了功能分化,它们可能在不同时间抑制雌、雄花器官的发育[22]。此外,相关研究发现,FLC除在花调控中发挥作用,对种子的萌发过程也有重要影响[23]。
SVP是响应环境温度以调控植物开花的另一个重要基因。SVP是MADS-box转录因子,位于自动途径FCA、FVE基因下游,其功能是维持营养生长,能够抑制开花但不改变光周期和春化作用对开花时间的影响[24]。SVP接受环境温度信号,调控FT的表达,此外,还接受自动途径以及GA诱导途径信号,在芽和叶片中直接调控SOC1的表达。近期研究表明,SVP是热敏感途径的关键基因[25]。SVP不调控FLC,但在营养生长阶段,SVP与FLC形成复合体,抑制FT的表达,二者协调作用,抑制开花。此外,SVP还与AGL24以及SOC1形成复合体,阻止拟南芥花分生组织的过早分化,促进花器官的正常发育。目前对木本果树SVP功能的研究主要集中在落叶果树休眠的开始和解除,包括猕猴桃[26]、枳壳[27]、悬钩子[28]、桃[29,30]等。桃有6个SVP同源基因,这6个基因至少有4种不同的表达模式[29],其中PpDAM5和PpDAM6的表达与芽的休眠密切相关[30];猕猴桃中发现了4个拷贝的SVP同源基因,在芽休眠和开花过程中具有不同功能[26]。
3 FT 与TFL基因
FT基因是光周期途径关键调控因子CO的直接靶基因,在叶片和维管组织特异表达,与LFY基因共同激活AP1的转录促进拟南芥开花,是可以长距离运输的开花素信号分子[31]。FT基因在木本植物成花中也有重要作用,杨树开花时FT同源基因的表达量上升[32,33]。转基因FT过量表达能使杨树[7]、梨树[9]早花。在杨树中发现了2个FT同源基因FT1和FT2,FT1响应低温调控生殖生长,FT2响应温暖的气温和长日照抑制芽转变,调控营养生长[7]。在龙眼中分离到2个与拟南芥FT高度同源的基因DlFT1和DlFT2,二者在拟南芥的异位表达使得开花时间出现变化,推测DlFT1使叶分生组织向花转变,促进开花,DlFT2则抑制开花[4]。柑橘童期长达6-10年。将从温州蜜柑中分离的FT同源基因CiFT1转入枳后持续表达,该35S::CiFT1转基因枳表现早花[34]。FT基因在植物生长阶段扮演着很多角色。龙眼DlFT2基因可能参与叶的形成[4]。在针叶树中,FT基因不仅与开花有关,还与童期随季节周期生长有关[35]。将苹果MdFT1和MdFT2基因转入拟南芥,发现它们参与调控细胞增殖和新生组织形成,并与TCP和VOZ基因家族蛋白相互作用而影响叶、果实的发育[3]。对温州蜜柑成年树进行低温处理后,随着茎和叶分生组织总CiFT1的表达量升高,树体开花潜力也增强,低温促使CiFTs表达量升高从而诱导温州蜜柑开花[5]。从以上研究可以看出,FT在木本果树中的功能出现分化,但过量表达FT会促进木本果树缩短童期,提早开花。
TFL1与FT基因同属一个家族,功能上却相反。拟南芥中TFL1基因抑制茎端分生组织形成花原基,延迟植物由营养生长向生殖生长的转变[36]。TFL1同源基因在发育阶段早期的芽中表达量较高,但在花发育过程中不表达[37]。TFL1能够抑制AP1对开花的正调控,在甜橙中发现的TFL1同源基因CsTFL与LFY和AP1在柑橘中的表达负相关[38]。近来研究发现,木本植物中TFL1基因的功能可能与童期有关。蔷薇科的玫瑰和草莓发现有连续开花的变异品种,变异性状都与TFL1有关,玫瑰是由于反转录转座子插入到其TFL1同源基因KSN中,使得KSN转录受阻,而草莓的连续开花变异是因为其TFL1编码区发生了2个碱基的缺失[39]。早花枳壳1年之中也能开花2-3次[40],FT、SOC1等基因的表达量增加,TFL1等抑制基因的表达量下降[18,40]。MdTFL1-1 和MdTFL1-2在苹果的童期和成年树营养组织中均表达,具有调控植物分生组织和抑制开花的作用[37]。将苹果MdTFL1基因在童期为5年的正常苹果植株中反义表达,转基因苹果在移栽温室8个月后即开始诱导开花[41]。抑制MdTFL基因的表达同样可以使苹果树缩短童期提早开花[42]。有人认为这可用于加快苹果育种[43]。拟南芥tfl1突变体腋生花序发育成顶花,茎端分生组织终止提前开花。利用转基因RNAi干扰使梨的PcTFL1-1、PcTFL1-2基因表达沉默,梨提早开花[44]。
4 花特性基因LFY、AP1和SOC1
LFY基因是花分生组织特性基因,不但控制花序分生组织向花分生组织的转变,还控制开花时间,在由营养生长向生殖生长转变过程中起主要的调控作用。目前已从多种木本果树如桃[45]、龙眼[46]、苹果[47]、银杏[48]、桉树[49]等中克隆到LFY的同源基因。
LFY基因的表达贯穿于花序和花发育的各个阶段,被认为是花发育的主要调控基因[50]。LFY基因的超量表达能使拟南芥花期提前[51]。把LFY基因转入杨树中,转基因植株的顶端开花时间显著提前,从8-20年缩短到几个月[8]。LFY与AP1基因对决定花分生组织特性是必需的。LFY基因在柑橘的过量表达能缩短童期而导致早花[52]。将LFY和AP1分别转入枳橙后出现早花,且产生可育花,转基因植株2-20个月后初次开花,最早1年后开始结果[10]。但从拟南芥获得LFY基因,转入苹果表达未出现早花现象,但植株表型有变化,猜测LFY基因在不同物种中可能具有不同的功能[6]。龙眼成花逆转时LFY同源基因LLFY的表达下降,表明LLFY与龙眼花芽分化及维持有关[53]。
在拟南芥中AP1编码MADS蛋白,既是花分生组织决定基因,又是花器官特征基因(花萼和花瓣特异调控基因),调节花分生组织的形成和花器官的确认与发生,突变会导致植物开花的延迟[54]。研究表明,AP1可以作为检测木本植物是否处于花发育阶段的标志[55]。甜橙成花转变时期CsAP1表达升高[38];相比营养枝,FT、AP1在苹果结果枝中的表达量更高[56],证明AP1基因与花器官生长有关。但是AP1和LFY的过量表达并不能使转基因苹果植株提前开花[6]。沙梨中AP1的同源基因PpAP1只在花芽或花中表达,转入拟南芥后呈现早花,证明PpAP1在花发育中起重要作用,但PpAP1在雄蕊少量表达说明它不是A功能基因[57]。从龙眼中分离到DlAP1-1和DlAP1-2基因,其中DlAP1基因序列高度保守,转入拟南芥后早于野生型开花,但长日照条件下35S::DlAP1-1转基因植株表现出晚花现象[3]。
SOC1基因也是开花途径整合子之一,属于MADS-box基因家族,受光周期途径、春化途径、自主途径和赤霉素途径等调控,促进营养分生组织向花序分生组织转变,而促进开花。光周期途径CO基因是SOC1基因的上游调节因子,但CO基因不直接调控SOC1的表达,而是通过作用于FT基因,间接激活SOC1基因,从而促进植物开花[58]。SOC1基因具有非常复杂的作用网络,在植物生长的各阶段都发挥着重要作用[58]。从芒果中克隆SOC1基因,该SOC1蛋白与橙SOC1蛋白亲缘关系最近,与葡萄、咖啡豆SOC1蛋白亲缘关系最远,因此推测SOC1基因在不同物种中的进化程度可能存在差异[59]。PTM5是山杨木中SOC1的同源基因,在维管组织中特异表达,与山杨木材的形成有关,该基因可能参与作用1年生植物和多年生植物之间的进化差异[60]。在4年生白桦茎尖中分离得到1条SOC1-likecDNABpSOC1,BpSOC1既参与白桦的营养生长,又参与生殖生长[61]。甜橙中SOC1的同源基因CsSL1和CsSL2在营养器官和花器官中均有表达,其中CsSL1在茎、叶、顶芽、花蕾和雄蕊处表达,CsSL2基因表达则更为广泛[62]。
5 展望
植物开花是遗传因子和环境因素协同作用的复杂过程,受错综复杂的网络信号途径调控。而木本植物大多数童期较长,始花期晚。柑橘的童期根据品种不同可持续5-10年不等,而一些林木经济作物如黄连木的童期则长达12年。嫁接等技术虽然能提早开花,但遗传性质未变,不能从本质上解决问题。尽管果树的遗传转化取得了一定的进展,但转化成功的案例普遍集中在少数品种。果树开花过程复杂,如柑橘结果树小年花诱导时期FT、AP1和LFY的表达量高于大年;但SOC1基因的表达量大小年无变化[63],说明人们对高等植物成花完全理解需要一个较长的研究过程。因此,借鉴模式植物的研究成果,利用现代分子生物学、基因组等技术研究开花途径关键基因的调控机理和调控网络,是真正实现木本植物成花遗传调控的基础。
[1]ZHOU C M,ZHANG T Q,WANG X,et al. Molecular basis of age-dependent vernalization in cardamine flexuosa[J]. Science,2013,340(10):1097-1100.
[2]BERGONZI S,ALBANI M C,THEMAAT E V L V,et al. Mechanisms of age-dependent response to winter temperature in perennial flowering ofArabisalpina[J]. Science,2013,340:1094-1097.
[3]MIMIDA N,KIDOU S I,IWANAMI H,et al. AppleFLOWERINGLOCUSTproteins interact with transcription factors implicated in cell growth and organ development[J]. Tree Physiology,2011(31):555-566.
[4]PATRICK W,PIMSIRI T,ALON S,et al. Isolation and characterization ofFLOWERINGLOCUSTsubforms andAPETALA1 of the subtropical fruit treeDimocarpuslongan[J]. Plant Physiology and Biochemistry,2013,71:184-190.
[5]NISHIKAWA F,ENDO T,SHIMADA T,et al. IncreasedCiFTabundance in the stem correlates with floral induction by low temperature in Satsuma mandarin (CitrusunshiuMarc.)[J]. Journal of Experimental Botany,2007,58(14):3915-3927.
[6]FLACHOWSKY H,HTTASCH C,MONIKA H,et al. Overexpression ofLEAFYin apple leads to a columnar phenotype with shorter internodes[J]. Planta,2010,231:251-263.
[7]HSU C Y,ADAMS J P,KIM H J,et al.FLOWERINGLOCUSTduplication coordinates reproductive and vegetative growth in perennialpoplar[J]. Proceedings of the National Academy of Sciences,2011,26(108):10576-10761.
[8]杨传平,刘桂丰,魏志刚.高等植物成花基因的研究[J].遗传,2002,24(3):379-384.
[9]MATSUDA N,IKEDA K,KUROSAKA M,et al. Early flowering phenotype in transgenic pears (PyruscommunisL.) expressing theCiFTgene[J]. Journal of the Japanese Society for Horticultural Science,2009,78:410-416.
[10]FRED G,GMITTER J,CHEN C X,et al. Citrus genomics[J]. Tree Genetics and Genomes,2012,8:611-626.
[11]KOBAYASHI Y,WEIGEL D. Move on up,it′s time for change—mobile signals controlling photoperiod-dependent flowering[J]. Genes and Development,2007,21(19):2371-2384.
[12]SAWA M,NUSINOW D,KAY S,et al.FKF1 andGIGANTEAcomplex formation is required for day-length measurement inArabidopsis[J]. Science,2007,318:261-265.
[13]MORRIS K,THORNBER S,CODRAI L,et al.DAYNEUTRALFLOWERINGrepressesCONSTANSto preventArabidopsisflowering early in short days[J]. The Plant Cell,2010,22(4):1118-1128.
[14]ALMADA R,CABRERA N,CASARETTO J A,et al.VvCOandVvCOL1,twoCONSTANShomologous geneareregulated during flower induction and dormancy in grapevine uds[J]. Plant Cell Reporter,2009,28:1193-1203.
[15]JEONG D H,SUNG S K,AN G. Molecular cloning and characterization ofCONSTANS-like cDNA clones of the Fuji apple[J]. Journal of Plant Biology,1999,42(1):23-31.
[16]WOLLENBERG A C,STRASSER B,CERDAN P D,et al. Acceleration of flowering during shade avoidance inArabidopsisalters the balance betweenFLOWERINGLOCUSC—mediated repression and photoperiodic induction of flowering[J]. Plant Physiol,2008,148(3):1681-1694.
[17]HOU X J,LIU S R,KHAN M R G,et al. Genome-wide identification,classification,expression profiling,and SSR marker development of the MADS-box gene family in citrus[J]. Plant Molecular Biology Reporter,2013,32(1):28-41.
[18]ZHANG J Z,LI Z M,MEI L,et al.PtFLChomolog from trifoliate orange (Poncirustrifoliata) is regulated by alternative splicing and experiences seasonal fluctuation in expression level[J]. Planta,2009,229:847-859.
[19]胡璐.PtFLC选择性剪切本的遗传转化及功能分析[D].武汉:华中农业大学,2013.
[20]ZHANG J Z,LI Z M,YAO J L,et al. Identification of flowering-related genes between early flowering trifoliate orange mutant and wild-type trifoliate orange (PoncirustrifoliataL.Raf.) by suppression subtraction hybridization (SSH) and macroarray[J]. Gene,2009,430(1-2):95-104.
[21]ZHANG J Z,AI X Y,SUN L M,et al. Molecular cloning and functional characterization of genes associated with flowering in citrus using an early-flowering trifoliate orange (PoncirustrifoliataL.Raf.) mutant[J]. Plant Molecular Biology,2011,76:187-204.
[22]杨希宏.山核桃CcFLC同源基因鉴定与表达分析[D].浙江:浙江农林大学,2012.
[23]CHIANG G C,BARUA D,KRAMER E M,et al. Major flowering time gene,FLOWERINGLOCUSC,regulates seed germination inArabidopsisthaliana[J]. Proceedings of the National Academy of Sciences,2009,106(28):11661-11666.
[24]LI D,LIU C,SHEN L,et al. A repressor complex governs the integration of flowering signals inArabidopsis[J]. Development Cell,2008,15(1):110-120.
[25]LEE J H,YOO S J,PARK S H,et al. Role ofSVPin the control of flowering time by ambient temperature inArabidopsis[J]. Genes and Development,2007,21(4):397-402.
[26]WU R M,WALTON E F,RICHARDSON A C,et al. Conservation and divergence of four kiwifruitSVP-like MADS-box genes suggest distinct roles in kiwifruit bud dormancy and flowering[J]. Journal of Experimental Botany,2012,63:797-807.
[27]LI Z M,ZHANG J Z,MEI L,et al.PtSVP,anSVPhomolog from trifoliate orange (PoncirustrifoliataL. Raf.),shows seasonal periodicity of meristem determination and affects flower development in transgenicArabidopsisand tobacco plants[J]. Plant Molecular Biology,2010,74:129-142.
[28]MAZZITELLI L,HANCOCK R D,HAUPT S,et al. Co-ordinated gene expression during phases of dormancy release in raspberry (RubusidaeusL.) buds[J]. Journal of Experimental Botany,2007,58:1035-1045.
[29]LI Z G,REIGHARD G L,ABBOTT A G,et al. Dormancy-associated MADS genes from theEVGlocus of peach [Prunuspersica(L.) Batsch]have distinct seasonal and photoperiodic expression patterns[J]. Journal of Experimental Botany,2009,60:3521-3530.
[30]YAMANE H,OOKA T,JOTATSU H,et al. Expressional regulation ofPpDAM5 andPpDAM6,peach (Prunuspersica) dormancy-associated MADS-box genes,by low temperature and dormancy-breaking reagent treatment[J]. Journal of Experimental Botany,2011,62:3481-3488.
[31]李昱,罗志鹏,赵淑清.拟南芥开花时间调控的整合途径[J].植物生理学通讯,2007,43(5):799-804.
[32]BÖHLENIUS H,HUANG T,CHARBONNEL-CAMPAA L,et al.CO/FTregulatory module controls timing of flowering and seasonal growth cessation in trees[J]. Science,2006,312:1040-1043.
[33]HSU C Y,LIU Y,LUTHE D S,et al.PoplarFT2 shortens the juvenile phase and Promotes seasonal flowering[J]. Plant Cell,2006,18:1846-1861.
[34]ENDO T,SHIMADA T,FUJII H,et al.Ectopic expression of anFThomolog fromCitrusconfers an early fowering phenotype on trifoliate orange (PoncirustrifoliataL.Raf.) [J]. Transgenic Research,2005,14:703-712.
[35]GYLLENSTRAND N,CLAPHAM D,KALLMAN T,et al. A Norway spruceFLOWERINGLOCUSThomolog is implicated in control of growth rhythm in conifers[J]. Plant Physiology,2007,144:248-257.
[36]常丽丽,吴连成,库丽霞,等.植物FLOWERINGLOCUST/TERMINALFLOWER1基因家族的研究进展[J].西北植物学报,2008,28(4):843-851.
[37]MIMIDA N,KOTODA N,UEDA T,et al. FourTFL1/CEN-like genes on distinct linkage groups show different expression patterns to regulate vegetative and reproductive development in apple (Malus×domesticaBorkh.)[J]. Plant Cell Physiol,2009,50:394-412.
[38]FLACHOWSKY H,LE ROUX P M,PEIL A,et al. Application of a high-speed breeding technology to apple (Malus×domestica) based on transgenic early flowering plants and marker-assisted selection[J]. New Phytol,2011,192:364-377.
[39]IWATA H,GASTON A,REMAY A. TheTFL1 homologueKSNis a regulator of continuous flowering in rose and strawberry[J]. The Plant Journal,2012,69:116-125.
[40]ZHANG J Z,AI X Y,SUN L M,et al. Transcriptome profile analysis of flowering molecular processes of early flowering trifoliate orange mutant and the wild-type [Poncirustrifoliata(L.) Raf.]by massively parallel signature sequencing[J]. BMC Genomics,2011b,12(1):1-20.
[41]KOTODA N,IWANAMI H,TAKAHASHI S,et al. Antisense expression ofMdTFL1,aTFL1-like gene,reduces the juvenile phase in apple[J]. Journal of the American Society for Horticultural Science,2006,131:74-81.
[42]KOTODA N,WADA M,MASUDA T,et al. The break-through in the reduction of juvenile phase in apple using transgenic approaches[J]. Acta Horticulturae,2003,625:337-343.
[43]PILLITTERI L J,LOVATT C J,WALLING L L,et al. Isolation and characterization of aTERMINALFLOWERhomolog and its correlation with juvenility inCitrus[J]. Plant Physiology,2004,135(3):1540-1551.
[44]FREIMAN A,SHLIZERMAN L,GOLOBOVITCH S,et al. Development of atransgenic early flowering pear (PyruscommunisL.) genotype by RNAi silencing ofPcTFL1-1 andPcTFL1-2[J]. Planta,2012,235:1239-1251.
[45]安丽君,李天红.桃成花基因PpLFY的克隆与表达及多克隆抗体制备[J].园艺学报,2008,28(8):1573-1580.
[46]官磊.龙眼LEAFY基因克隆与功能研究[D].福州:福建农林大学,2008.
[47]WADA M,CAO Q F,KOTODA N,et al. Apple has two orthologues ofFLORICAULA/LEAFYinvolved in flowering[J]. Plant Molecular Biology,2002,49:567-577.
[48]ZHANG J Y,CHEN L G,HU X Q,et al.LEAFYhomologous gene cloned in maiden hair tree (GinkgobilobaL.)[J]. Scientia Silvae Sinicae,2002,38(4):167-170.
[49]SOUTHERTON S G,STRAUSS S H,OLIVE M R,et al. Eucalyptus has a functional equivalent of theArabidopsisfloral meristem identity geneLFY[J]. Plant Molecular Biology,1998,37:897-910.
[50]MOYROUD E,KUSTERS E,MONNIAUX M,et al. LEAFY blossoms[J].Plant Science,2010,6(15):346-352.
[51]何新华,郭永泽,张利.金柑LEAFY同源基因克隆与全序列分析[J].广西农业生物科学,2007,26(4):273.
[52]PENA L,MARTIN-TRILLO M,JUAREZ J,et al. Constitutive expression ofArabidopsisLEAFYorAPETALA1 genes in citrus reduces their generation time[J]. Nature Biotechnology,2001,19:263-267.
[53]ZENG L H,GUAN L,WU S H.LLFY,a LonganLEAFYortholog,is associated with differentiation and maintenance of inflorescence bud[J]. International Society for Horticultural Science,2010,863:123-128.
[54]段艳欣,郭文武.木本植物开花调节基因的分离克隆及其童期控制[J].中国生物工程杂志,2004,24(10):22-26.
[55]JAYA E S,CLEMENS J,SONG J,et al.Quantitative expression analysis of meristem identity genes inEucalyptusoccidentalis:AP1 is an expression marker for flowering[J]. Tree Physiol,2010,30:304-312.
[56]KOTODA N,HAYASHI H,SUZUKI M,et al. Molecular characterization ofFLOWERINGLOCUST-like genes of apple (Malus×domesticaBorkh.)[J]. Plant and Cell Physiology,2010,51(4):561-75.
[57]LIU Y X,KONG J,LI T Z,et al. Isolation and characterization of anAPETALA1-like gene from pear (Pyruspyrifolia)[J]. Plant Molecular Biology Reporter,2013,31:1031-1039.
[58]LEE J,LEE I.Regulation and function ofSOC1,a flowering pathway integrator[J]. Journal of Experimental Botany,2010,61(9):2247-2254.
[59]罗聪.芒果SCoT分子标记与逆境和重要开花时间相关基因研究[D].广西:广西大学,2012.
[60]CSEKE L J,ZHENG J,PODILA G K. Characterization ofPTM5 in aspen trees:a MADS-box gene expressed during woody vascular development[J]. Gene,2003,318:55-67.
[61]刘菲菲,李慧玉,王姗.白桦BpSOC1基因的克隆及时序表达分析[J].东北林业大学学报,2011,39(4):1-4.
[62]TAN F C,SWAIN S M.Funetional charaeterization ofAP3,SOC1 andWUShomologues from citrus(Citrussinensis)[J]. Physiologia Plantarum,2007,131:481-495.
[63]SHALOM L,SAMUELS S,ZUR N,et al. Alternate bearing inCitrus:changes in the expression of flowering control genes and in global gene expression in ON-versus OFF-crop trees[J]. PLOS ONE,2012,7(10):1-16.
(责任编辑:陈幼玉)
Floweringintegratorgenesinwoodyfruittrees
WEI Dan-feng,ZENG Li-hui
(College of Horticulture,Fujian Agriculture and Forestry University,Fuzhou,Fujian 350002,China)
Flowering is an important process for plants to transfer from vegetative to reproductive development. Studies onArabidopsisrevealed several pathways were involved in flowering time control,such as photoperiod pathway,vernalization pathway,GA pathway,and autonomous pathway. Compared with herbs,the research progress of flowering regulation in woody plants is slowly and have a big divergence. This review summarizes recent progresses on flowering integrator genes such asCO,FLC,SVP,FT,TFL,AP1,SOC1 andLFYin woody fruit trees,so as to provide reference for further explore about flowering molecular mechanism of woody plants.
woody fruit tree; flowering pathways; flowering genes
2014-03-12
魏丹凤(1988- ),女,硕士研究生。研究方向:果树分子遗传。Email:15980248065@163.com。通讯作者曾黎辉(1973- ),女,教授,博士生导师。研究方向:果树遗传育种与分子生物学。Email:lhzeng@hotmail.com。
Q945
A
1673-0925(2014)02-0134-07
猜你喜欢
蔬菜(2022年1期)2022-11-11
麦类作物学报(2022年4期)2022-05-23
金山(2021年10期)2021-11-02
汉字汉语研究(2021年2期)2021-08-30
汉字汉语研究(2019年2期)2019-08-27
热带生物学报(2019年1期)2019-04-18
江苏农业科学(2018年11期)2018-06-29
现代园艺(2017年23期)2018-01-18
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
上海农业学报(2017年3期)2017-04-10
