
比格犬雌激素受体β四个剪接异构体的发现
2014-08-14 07:57:42李建瑛许永华任秀梅赵彦斌白杰英孙兆增胡仲明
中国比较医学杂志 2014年11期
许 琴,董 翔,李建瑛,许永华,任秀梅,赵彦斌,白杰英,孙兆增,曾 林,胡仲明
(1. 军事医学科学院实验动物中心,北京 100071; 2. 兰州军区乌鲁木齐总医院动物实验科,乌鲁木齐 830000)
作为标准实验动物,比格犬在医药、公共卫生、生命科学以及军事医学研究领域得到了广泛的应用。但随着实验动物化时间的延长以及长期的封闭群繁殖饲养,我国比格犬种群中出现了不同程度的繁殖障碍。为解决上述生产中遇到的问题,对比格犬生殖生理进行研究很有必要。雌激素通过其受体在下丘脑-垂体-卵巢轴对雌性动物的生殖机能进行着周期性的调控。雌激素受体有三种亚型α、β、γ(Estrogen receptor α, β, γ, ERα, ERβ, ERγ),其中ERγ仅存在于鱼类[1]。有研究表明在不同生殖周期中,ERα、ERβ的表达水平,表达时间及组织表达部位不同[2],并且ERα敲除小鼠雌性不育,雄性繁殖力严重下降,ERβ敲除小鼠雌性与雄性能够繁殖,但产仔率降低,窝产仔数明显减少,并且已有研究表明ERβ是一种雌激素依赖性肿瘤的抑制因子[3-5],为了解ERβ在比格犬下丘脑-垂体-性腺轴发挥的调控作用,研究团队自2011年以来,对比格犬生殖内分泌轴上ERβ的剪接异构体存在情况进行了筛查[6-7],发现了ERβ的四个剪接异构体,现总结报告如下。
1 材料和方法
1.1 实验材料
1.1.1 主要仪器
PCR扩增仪(PTC-2000,BIO-RAD),高速冷冻离心机(3-18K,SIGMA),-80℃超低温冰箱(NU-9483E,NUAIRE),微量分光光度计(NanaDrop 2000,Thermo),核酸电泳仪(ESP1001, amersham pharmacia biotech),紫外凝胶成像仪(ACI,UVP),180℃干烤手术器械一套,移液器一套(Eppendof)。
1.1.2 主要试剂
Trizol购自 Invitrogen 公司,DEPC购自 Promega 公司,ReverTra-Ace反转录试剂盒购自TOYOBO公司,pMD18-T Vector, LA Taq酶,dNTPmix,2×GC buffer购自大连宝生物公司TaKaRa产品,标准分子量核酸DNA Marker DL-2000 购自康为世纪公司,DNA片段凝胶回收试剂盒,质粒提取试剂盒为博迈德公司BioFlux产品,其它试剂均由军事医学科学院条件处提供,为进口分装或国产分析纯试剂。
1.1.3 菌种及实验动物
大肠杆菌DH5α感受态细胞购自博迈德生物公司。
雌性比格犬11只(9只间情期犬,编号为1-9号;2只发情期犬,编号为F1,F2),18~20 kg,由军事医学科学院实验动物中心提供【SCXK (军) 2012-001】,取材在军事医学科学院实验动物中心实验室内进行【SYXK(军)2012-005】。
1.2 实验方法
1.2.1 组织标本的获取
取材器械包高压灭菌后,180℃干烤8 h以上备用,组织冻存管0.1% DEPC水处理后高压灭菌备用。实验动物用速眠新Ⅱ注射液(1.5 mL/支),按照0.08~0.1 mL /kg体重肌肉注射,待动物麻醉瘫软后,颈动脉放血处死动物,迅速留取下丘脑、垂体、子宫、卵巢组织标本,于冻存管中液氮冻存。
1.2.2 引物设计
根据NCBI网站公布的比格犬雌激素受体-β mRNA CDS区序列(犬ESR2 mRNA 24转录本,gene 1968 bp,CDS区1593 bp,Gene ID:403639)设计两对引物:真核ERβ-F:CCCAAGCTTATGGATATCAA AAACTCTCCATCTAGCC;真核ERβ-R:CGCGGA TCCTCACTGAGATGTGGTCTTCTGGGAGC,用于扩增全长序列。经限制性内切酶分析后,在犬ERβ一对引物的上游引入Hind Ⅲ,下游引入BamH Ⅰ内切酶位点,为构建真核表达重组子,在引物的下游均去除了终止密码子TGA。
筛查比格犬ERβ可能存在的两两外显子缺失的剪接异构体,设计引物如下:
1.2.3 下丘脑、垂体、卵巢、子宫组织总RNA的提取
采用Trizol一步法提取比格犬下丘脑、垂体、卵巢、子宫总RNA。微量分光光度计测定A260、A280的吸光度值,准确定量总RNA的浓度和纯度。取1 μg RNA用于1%琼脂糖电泳,以检测RNA的完整性。将RNA储存于-80℃冰箱备用。按照ReverTra Ace(TOYOBO)试剂盒操作说明书,将总RNA反转录为cDNA,反转录产物保存于-20℃冰箱中。
1.2.4 PCR扩增ERβ基因
ERβ基因扩增:模板为7丘(即7号犬下丘脑)cDNA,扩增体系25 μL如下:PCR 反应程序: 95℃预变性5 min;95℃变性30 s;56℃退火30 s;72℃延伸2 min;33个循环;最后延伸8 min。PCR产物进行1%琼脂糖电泳查看结果。
ERβ可能的两个外显子缺失筛查PCR反应:模板为ERβ全长PCR产物,引物为针对可能的外显子缺失所设计,反应体系同ERβ基因扩增体系。

引物名称引物序列(5、-3、)产物长度针对的外显子Exon 1-3FATGGATATCAAAAACTCTCCA1/2/两者Exon 1-3RGCATATGTAATCATTATG555 bp(1-555)Exon1-4FCCTGTAAACAGAGAGACACTG2/3/两者Exon1-4RGGTCACGTGGCCACCATT408 bp(352-759)Exon2-5FTGGTCGTGTGAAGGATGTAAGGCC3/4/两者Exon2-5RGAGCTCCACAAAGCCTGGAAT477 bp(490-966)Exon3-6FGTGAAGTGTGGCTCCCGGAGA4/5/两者Exon3-6RCCCCTCATCCCTGTCCAG459 bp(643-1101)Exon4-7FTTCACCGAGGCCTCCATG5/6/两者Exon4-7RCTTCCGGCTGCTCTCCGCCTCCTG411bp(865-1275)Exon 5-8FGACCACCCGGGCAAGCTCATC6/7/两者Exon 5-8RCAGCAGCAGGTCGTAGACTGGGAC432 bp(1045-1476)Exon6-8FTGCGTAGAAGGAATTCTGGAA7/8/两者Exon6-8RCCCGCGGAGTGTGTGCGC402 bp(1105-1506)Exon7-8FAAGGCCATGGTCCTCCTCAACTCC7/8/两者Exon7-8RTCACTGAGATGTGGTCTTCTG393 bp(1201-1593)
PCR反应程序为:95℃预变性5 min;95℃变性30 s;56℃退火30 s;72℃延伸30 s;33个循环;最后延伸8 min。PCR产物进行1.5%琼脂糖电泳查看结果。
1.2.5 PCR扩增产物的纯化
PCR扩增产物的纯化按照康为世纪公司DNA胶回收试剂盒回收相应目的片段,按照试剂盒说明书进行。纯化后的PCR产物与pMD-18T载体连接,16℃连接过夜。连接产物参照博迈德公司DH5α感受态细胞操作步骤进行转化。菌液PCR鉴定阳性克隆。初步鉴定为阳性的菌液,提取质粒送北京中科西林测序部测序鉴定。
2 实验结果
2.1 比格犬下丘脑、垂体、卵巢、子宫组织总RNA的提取
比格犬下丘脑,垂体,卵巢,子宫组织总RNA提取(图1),ERβ的扩增及菌液鉴定(图2)。

注:①F2犬下丘脑,垂体,卵巢,子宫,M: DL2000 DNA 标准分子量;② 3号犬卵巢与子宫,M: DL10000 DNA 分子量标准;③7号犬垂体与下丘脑。

注:①7号犬下丘脑,垂体;② F2犬子宫,卵巢,下丘脑,垂体;③PCR扩增产物菌液鉴定7号犬垂体与下丘脑;①②③中M均为DL2000 DNA 分子量标准。
比格犬下丘脑、垂体、卵巢、子宫组织总RNA的浓度测定结果见表1。
表1 下丘脑、垂体、卵巢、子宫总RNA浓度
由图1及表1中结果可见,提取的组织总RNA 1%琼脂糖电泳中28s和18s两条核糖体RNA条带清晰可见,比较完整,浓度测量纯度较高,满足后续基因扩增的要求。经过预实验确定,以F2丘(F2犬下丘脑cDNA)为模板扩增ERβ mRNA,以F2垂(F2发情期犬垂体)、7丘(7号犬下丘脑)cDNA为模板进行ERβ可能存在的两个外显子缺失剪接异构体的筛查。
2.2 ERβ基因测序结果
以7号犬下丘脑,垂体,2号发情犬(F2)下丘脑、垂体为模板进行扩大体积扩增,均在1000 bp~2000 bp之间有两条带,经比对两条带都是比格犬ERβ 24转录本 mRNA序列,产物长度分别为1593 bp、1293 bp(比对结果此文未列出)。分析ERβ1593与ERβ1293的mRNA序列,后者在ERβ1593第一外显子 ATG起始密码子之前增加63 bp碱基,为载体序列,并且在第656-955位缺失300 bp,所缺失部分恰好是ERβ1593的第四外显子的完整部分,推测是ERβ1593的一个剪接异构体。
2.3 ERβ可能的两两外显子缺失筛查结果
2.3.1 引物筛查及可能的外显子缺失异构体片段的筛查
所有引物以相同的模板,根据引物的Tm值,设计温度梯度进行温度梯度PCR,根据扩增产物对引物进行筛查(图3)。
由扩增电泳图可知,Exon1-4在48℃~58℃,Exon2-5在51℃~61℃,Exon4-7在52℃~64℃的温度梯度内的扩增产物均为单一条带,或主带之外有非常弱的非特异扩增带(如1-4),但不能后用于后续的回收纯化连接及转化,所以不能用于后续的可能的外显子缺失型的剪切异构体的筛查,Exon1-3,Exon3-6,Exon5-8,Exon6-8可用于后续的筛查。用后面的这四对引物,由其最适扩增条件进行新一轮扩增(图4-①)。
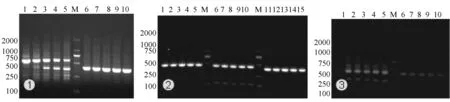
注:①引物1-3,1-4温度梯度筛查:1~5由引物Exon1-3扩增,6~10由引物Exon1-4扩增;②引物2-5,3-6,4-7温度梯度筛查:1~5由引物Exon2-5扩增,6~10由引物Exon3-6扩增,11~15由引物Exon4-7扩增;③引物5-8,6-8温度梯度筛查:1~5由引物Exon5-8扩增,6~10由引物6-8扩增。M. DM2000 DNA 标准分子质量。

注:1-3,3-6,5-8,6-8为引物代码,M. DM2000 DNA 标准分子质量。
Exon1-3扩增产物主带为555 bp,与预期结果一致,主带之上、之下各有一条可回收的非特异带:1-3上、1-3下;Exon3-6扩增产物主带为459 bp,与预期结果一致,主带之下有一条可回收的非特异带:3-6下;Exon5-8扩增产物主带为432 bp,与预期结果一致,主带之下有一条明显的非特异带:5-8下;Exon6-8扩增产物主带为402 bp,与预期结果一致,主带之下也有一条明显的非特异带:6-8下。将所扩增出的条带进行回收纯化(见图4-②),并测序比对鉴定。
2.3.2 ERβ外显子缺失异构体片段的测序比对
由四对引物回收纯化后的产物,测序后在NCBI网站上进行比对,均为比格犬ERβmRNA 部分CDS序列,经DNDMAN软件比对和手动校对后,获得了比格犬ERβ的4个剪接异构体的部分序列。
Exon3-6引物所辖定的主带序列与3-6下产物125 bp运用DNNMAN软件比对后,有部分错码,经手动校对,结果发现部分第4外显子与部分第5外显子缺失,缺失334 bp,缺失段为第4外显子289 bp(阴影标示)和第5外显子45 bp(下划线标示),是比格犬ERβ的一种剪接异构体形式,比对结果见图5,图6。

图5 部分第4与部分第5外显子缺失的DNAMAN比对

图6 部分第4与部分第5外显子缺失的手动比对
Exon3-6主带与3-6下194 bp产物运用DNAMAN软件比对后有部分乱码,经手动校对发现265 bp缺失,缺失段为部分第四外显子(163 bp,阴影标示部分)与部分第五外显子(102 bp,下划线标示部分)。结果发现是另一种部分第4与部分第5外显子组合缺失型的比格犬ERβ剪接异构体形式,比对结果见图7,图8。

图7 部分第4与部分第5外显子缺失的DNAMAN比对

图8 部分第4与部分第5外显子缺失的手动比对
Exon3-6主带与3-6下159 bp产物运用DNAMAN软件比对后也出现部分乱码,经手动校对发现300 bp缺失,缺失段为完整的第4外显子序列,结合ERβ主基因的扩增,及ERβ1293的出现可判定ERβ第四外显子缺失是比格犬ERβ基因的又一种剪接异构体形式,图9,图10。

图9 第四外显子完整缺失DNAMAN软件比对

图10 第四外显子完整缺失的手动比对

图11 第七外显子完整缺失的DNAMAN软件比对
Exon5-8主带(432 bp)与5-8下回收产物(251 bp)运用DNAMAN比对时也出现部分序列乱码,经手工校对后发现了比格犬ERβ的第4种剪接异构体,选择性剪接发生在第7外显子上,缺失181 bp,是第7外显子编码序列的完整缺失,比对结果见图11,图12。

图12 第七外显子完整缺失的手动比对

图13 第7外显子完整缺失(引物Exon6-8)
Exon6-8主与6-8下比对也发现了第七外显子(181 bp)的完整缺失,是ERβ基因的第七外显子完整缺失型剪切异构体,比对结果见图13。经5`,3`RACE获得了比格犬ERβ第七外显子缺失剪切异构体的全长编码序列ERβ1257,因第七外显子的缺失使主基因发生了移码突变,翻译提前终止(测序结果此文未列出)。
结合DNAMAN软件比对及手动校对结果,应用所涉及的两两筛查引物,本实验共筛查出了比格犬ERβ的四个剪接异构体,分别为:第4外显子完整缺失型剪接体;第4外显子缺失289 bp与第5外显子缺失45 bp(总计缺失334 bp)的组合缺失型剪接体Ⅰ;第4外显子缺失163 bp与第5外显子缺失102 bp(总计缺失265 bp)的组合缺失型剪接体Ⅱ;第7外显子完整缺失型剪接体。对第4外显子完整缺失及第7外显子完整缺失的剪接异构体,研究小组通过5、-3、RACE获得了全长序列,并构建了真核表达重组子,对其表达及功能进行了系列的研究[8-9],而对于第4与第5外显子部分缺失的两种组合型缺失剪接异构体,本研究仅发现了部分编码序列,并非全长CDS序列。
3 讨论
比格犬ERβ位于8号染色体上(XM_861041.2,gene ID:403639),全长1968 bp,CDS区1593 bp,编码530个氨基酸(aa)[10]。本实验在扩增比格犬ERβ全长序列时在主带之下有一条明显条带,将两条带均回收纯化克隆,得到两个产物,一为ERβ1593,另一为ERβ1293,前者是比格犬野生型的ERβ,后者是比格犬ERβ的一种剪接异构体,其第四外显子完整缺失。
针对ERβ的每一个外显子,两两顺次组合设计引物(每对引物包含两个完整的外显子),进行扩增,对只能扩增出单一条带的引物进行剔除,而对除目的条带外,还能扩增出比较明显的可以回收的非特异条带(也可能是ERβ的一种剪接异构体)的引物,进行扩大体积扩增并回收纯化克隆产物,测序后在NCBI网站比对,结果确定是目的基因的,再与主带进行比对分析,如此我们得到了第4外显子完整缺失的ERβ剪接异构体(缺失300 bp),部分第4与部分第5外显子组合缺失的两个组合型缺失的ERβ剪接异构体(各缺失334 bp,265 bp),以及第七外显子完整缺失的ERβ剪接异构体(缺失181 bp),根据实验分析,及其它种属的相关文献报道[11-16],比格犬ERβ还存在其它形式的剪接异构体,但因在不同的生理条件或者病理条件、不同的发育时期、不同的组织或不同的细胞内,其表达时期及表达水平不同,所以对这些情况还需进行扩大碱基序列再次排查,该项工作课题组其它研究成员正在进行。
本研究中扩增出的比格犬ERβ mRNA CDS区第七外显子(181 bp)完整缺失,导致ERβ阅读框移位,使其在终止密码子之前引入了10个非ERβ编码的氨基酸残基。该发现与Fitzgerald SD在ERα上的发现相似[17]。García Pedrero等[18]在多种正常组织和肿瘤中分离出了ERα的一种剪切异构体ER△E7,ER△E7第七外显子完整缺失,失去了与p160辅激活因子(如SRC-1,AIB1)的雌激素依赖性的相互作用,ER△E7能以激素依赖的形式与 ERα 和ERβ 形成异源二聚体。本研究发现第七外显子完整缺失的ERβ剪接异构体,所缺失的相应氨基酸编码序列为409-469aa,是野生型ERβ配体结合区298-469aa中的部分序列,部分配体结合序列的缺失,是否引起该剪接异构体配体结合能力及信号转导途径的改变,以及缺失序列之后掺入的十个氨基酸残基与野生型ERβ的配体结合区是否具有同源性等,对此还需进一步进行实验验证。
参考文献:
[1] Gamba L, Cubedo N, Ghysen A,etal. Estrogen receptor ESR1 controls cell migration by repressing chemokine receptor CXCR4 in the zebrafish posterior lateral line system[J]. Proc Natl Acad Sci U S A, 2010 , 107(14):6358-6363.
[2] Kuiper GG, Carlsson B, Grandien K,etal.Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta [J]. Endocrinology, 1997,138(3):8632-8701.
[3] Couse JF, Curtis Hewitt S, Korach KS. Receptor null mice reveal contrasting roles for estrogen receptor alpha and beta in reproductive tissues [J]. J Steroid Biochem Mol Biol, 2000, 74:287-296.
[4] Krege JH, Hodgin JB, Couse JF,etal. Generation and reproductive phenotypes of mice lacking estrogen receptor β[J]. Pro Natl Acad Sci USA, 1998, 95: 15677 - 15682.
[5] Ogawa S, Chester AE, Hewitt SC,etal. From the cover: abolition of male sexual behaviors in mice lacking estrogen receptors alpha and beta (alpha beta ERKO) [J]. Proc Natl Acad Sci USA, 2000, 97:14737-14741.
[6] 孔德强,李爱学,曾林,等. 一种比格犬雌二醇β受体剪切异构体的克隆和序列分析[J]. 中国比较医学杂志, 2011, 21(7):44-47.
[7] 任秀梅,赵彦斌,孙兆增,等.比格犬一种新雌激素β 受体可变剪切体的克隆及鉴定[J]. 中国比较医学杂志,2012,22(7):36-39.
[8] 甘艺,钟睿,任秀梅,等. 比格犬ERβ1293基因真核载体的构建及其在HEK293T细胞中的表达和定位[J]. 中国比较医学杂志,2014,24(2):7-10,15.
[9] 钟睿,甘艺,赵彦斌,等. 比格犬雌激素β受体RNA干扰实验细胞及PCR检测引物的筛选[J]. 中国比较医学杂志,2014,24(2):42-45,51.
[10] www.ncbi.nlm.nih.gov
[11] Poola I, Abraham J, Liu A. Estrogen receptor β splice variant mRNAs are differentially altered during breast carcinogenesis[J]. J Steroid Biochem Mol Bio,2002,l82:169-179.
[12] Poola I, Abraham J, Baldwin K,etal. Identification of ten exon deleted ERβ mRNAs in human ovary, breast, uterus and bone tissues: alternate splicing pattern of estrogen receptor β mRNA is distinct from that of estrogen receptor α. FEBS Lett,2002, 516:133-138.
[13] Friend K, Ang L, Shupnik M. Estrogen regulates the expression of several different estrogen receptor mRNA isoforms in rat pituitary[J]. Proc Natl Acad Sci USA,1995, 93:5925-5930.
[14] Gunawan A, Kaewmala K, Uddin MJ,etal. Association study and expression analysis of porcine ESR1 as a candidate gene for boar fertility and sperm quality[J]. Anim Reprod Sci, 2011,128(1-4):11-21.
[15] Griffin C, Flouriot G, Buck VS,etal. Identification of Novel Chicken Estrogen Receptor-α Messenger Ribonucleic Acid Isoforms Generated by Alternative Splicing and Promoter Usage [J]. Endo, 1998, 139(11):4614-4625.
[16] Oliveira AG, Dornas RA, Mahecha GA,etal. Occurrence and cellular distribution of estrogen receptors ERα and ERβ in the testis and epididymal region of roosters[J]. Gen Comp Endocrinol, 2011, 170(3): 597-603.
[17] Fitzgerald SD, Allred D,etal. Inhibition of estrogen receptor action by a naturally occurring variant in human breast tumors[J]. Cancer Res,1992, 52:483-486.
[18] García Pedrero, Zuazua Pedro, Martínez-Campa Carlos,etal. The Naturally Occurring Variant of Estrogen Receptor(ER) ER△E7 Suppresses Estrogen-Dependent Transcriptional Activation by Both Wild-Type ERα and ERβ [J]. Endocrinology, 2003, 144(7):2967-2976.
猜你喜欢
小星星·阅读100分(高年级)(2024年2期)2024-04-23 19:23:30
系统仿真技术(2022年4期)2023-01-17 13:01:44
云南化工(2021年8期)2021-12-21 06:37:38
小读者·阅世界(2020年5期)2020-06-01 07:54:30
故事会(2009年8期)2018-09-03 00:00:00
中老年健康(2017年9期)2017-12-13 07:16:39
中成药(2017年6期)2017-06-13 07:30:34
国外医药(抗生素分册)(2016年4期)2016-07-12 14:25:19
中国民族医药杂志(2016年7期)2016-05-09 07:49:19
信息记录材料(2016年4期)2016-03-11 15:22:30
