
北五味子总木脂素减轻内质网途径凋亡延缓小鼠脑衰老*
2014-08-13 12:42于春艳于春荣鞠文博姜恩平陈建光
中国病理生理杂志 2014年11期
于春艳, 于春荣, 李 贺, 鞠文博, 姜恩平, 4△,陈建光△
(1北华大学基础医学院,吉林 吉林 132013; 2北京昭衍新药研究中心股份有限公司毒理部,北京 100176; 3北华大学药学院,吉林 吉林 132013; 4广东医学院, 广东 东莞 523808)
随着人们年龄的增长、老龄人口的增多,研究衰老的机制以及如何缓解衰老引起学者们越来越多的关注。脑衰老是人衰老最明显的现象,脑衰老的过程极其复杂,研究认为脑组织结构与功能的改变由多种因素共同作用导致,学习和记忆能力的减退是最重要的表现[1]。北五味子[Schisandrachinensis(Turcz.)Baill]属木兰科植物,据报道其中有效成分五味子多糖可使衰老小鼠已退行性变的神经细胞恢复正常,且可使神经细胞胞体增大并可明显促进衰老小鼠神经细胞的发育[2]。北五味子是否具有缓解脑衰老作用尚需进一步研究。关于衰老的发生机制有许多学说,如自由基学说、端粒学说和细胞凋亡学说等[3]。研究发现,在细胞水平,衰老主要体现在细胞膜流动性降低、脂褐质色素累积、酶活性下降和蛋白质聚集体增加等[4]。内质网是细胞内最重要的负责蛋白质折叠的细胞器,未折叠蛋白或错误折叠的蛋白在内质网内堆积即发生内质网应激。持续的内质网应激会诱导细胞发生凋亡[5]。本课题组已利用D-半乳糖复制小鼠衰老模型,给予不同剂量的北五味子总木脂素(schisandra total lignin,SCL),结果显示北五味子总木脂素给药组延缓小鼠衰老,并增加衰老小鼠肌肉协调性、增强衰老小鼠学习记忆能力[6],此次实验观察代表蛋白聚集物堆积的泛素化蛋白(ubiquitin,Ub)的表达水平以及葡萄糖调节蛋白78(glucose-regulated protein 78,GRP78)、硫氧还蛋白二硫键异构酶(protein disulfide isomerase,PDI)和C/EBP同源蛋白(CCAAT/enhancer-binding protein homologous protein,CHOP)表达水平的变化,凋亡相关蛋白Bcl-2、Bax的表达情况,用以明确北五味子总木脂素抗衰老作用及其可能的作用机制。
材 料 和 方 法
1 试剂
北五味子总木脂素由北华大学林学院自制提纯,实验时用蒸馏水稀释溶解至所需浓度;D-半乳糖购自Sigma;SP试剂盒购于福州迈新试剂公司;Ub、GPR78、PDI、CHOP、Bcl-2、Bax及β-actin抗体购自Santa Cruz。
2 衰老小鼠模型的建立、分组与给药
健康昆明小鼠,雌雄各半,体重18~20 g,吉林大学实验动物部提供。合格证号为SCXK-(吉)2008-0003。清洁级动物室饲养,饮水不限,定时喂食。
参照文献[7]复制衰老小鼠模型。取小鼠50只,雌雄各半,经适应性饲养1周,按体重随机分为5组: 空白对照组、模型组、五味子总木脂素低剂量组、五味子总木脂素中剂量组和五味子总木脂素高剂量组,每组10只,正常饮食。除空白对照组外,其余各组每天1次颈、背部皮下注射D-半乳糖100 mg/kg,连续注射10周。同时,五味子总木脂素低剂量组、中剂量组、高剂量组分别按35 mg/kg、70 mg/kg、140 mg/kg蒸馏水配制药液,灌胃给药,每天1次,连续给药10周[8]。断髓处死小鼠,冰上低温解剖取脑,部分多聚甲醛固定,部分冻存于-80 ℃。
3 方法
3.1学习记忆能力测试(小鼠跳台实验) 学习训练于停止给药后次日进行。每组各取1只,平行操作。实验装置为一长方形反射箱,大小为32 cm×22.5 cm× 33 cm,用黑色塑料板分隔成6间。底面铺以铜栅,间距为0.5 cm,每间左角置一高度和直径均为4.5 cm 的橡胶平台。将小鼠置于跳台仪隔间内,适应环境3 min,然后立即通以32 V 交流电。其正常反应是跳回平台以躲避伤害性刺激,以双足同时接触铜栅为错误反应,训练5 min(避免电击死亡)。每只训练3 次。
记忆测试于学习训练24 h 后进行。将小鼠放置于跳台上,同时按动记时器,记录第1 次跳下平台时间,此为触电潜伏期。并记录5 min 内跳下平台受到电击的次数,即错误反应数,作为记忆指标。
学习记忆实验结束后断头处死,后立即于冰台上解剖取出全脑组织,用预冷的生理盐水反复冲洗,滤纸吸干,用于Western blotting实验的脑组织置于-80 ℃ 超低温冰箱保存待用,用于切片的脑组织置于4%多聚甲醛中固定。
3.2Western blotting检测Ub、PDI、GRP78、CHOP、Bcl-2和Bax蛋白的表达 用RIPA裂解液提取细胞总蛋白,以β-actin的水平作为等量蛋白质上样对照,取50 μg蛋白质样品进行SDS-PAGE后, 转至硝酸纤维素膜上;5%奶粉室温封闭2 h,用含0.01% Tween 20的TBS缓冲液 (TBST) 漂洗3次,每次10 min;加入相应的抗体Ub (1∶200)和PDI (1∶200)、GPR78 (1∶200)、CHOP(1∶200),Bcl-2(1∶200)和Bax(1∶200),4 ℃ 孵育过夜;TBST漂洗3次后加入相应的辣根过氧化物酶标记的Ⅱ抗 (1∶1 000),37 ℃摇床温育2 h;TBST漂洗3次,每次10 min;DAB显色,凝胶图像分析系统(上海天能科技有限公司GIS凝胶成像系统)拍照,同时以β-actin为内参照,进行蛋白表达分析,蛋白表达量用光密度比值表示。
3.3免疫组化方法检测Bcl-2和Bax蛋白的表达 应用SP 免疫组化染色法。抗Bcl-2多克隆抗体以1∶100稀释,抗Bax单克隆抗体以1∶100稀释,Ⅱ抗为羊抗兔/鼠IgG 抗体。显微镜(Olympus) 下观察组织中蛋白表达,并摄片。
4 统计学处理
采用SPSS 13.0统计软件进行统计分析,数据以均数±标准差 (mean±SD) 表示,多组间样本均数比较采用t检验。以P<0.05为差异有统计学意义。
结 果
1 小鼠跳台实验检测北五味子总木脂素对衰老小鼠学习记忆能力的影响
在学习测试中,模型组小鼠5 min 内错误次数较对照组增多(P<0.05),SCL低(L)、中(M)、高(H)剂量组小鼠5 min 内错误次数较模型组减少(P<0.05)。在记忆测试中,模型组小鼠首次跳下平台的潜伏期较空白对照组缩短(P<0.05),5 min内错误次数增多(P<0.05);与模型组比较,SCL低、中、高剂量组小鼠首次跳下平台的潜伏期延长(P<0.05),5 min 内错误次数减少(P<0.05),见表1。
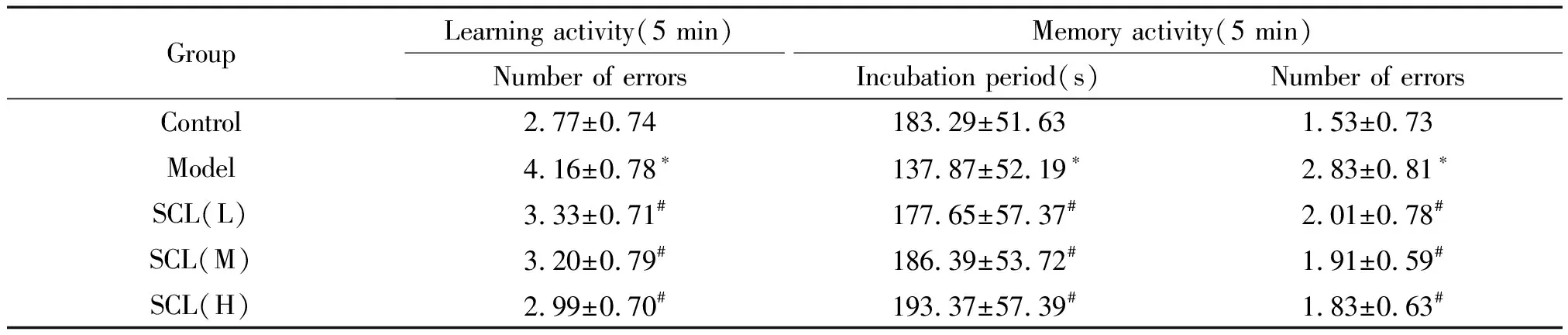
表1 北五味子总木脂素对衰老小鼠学习记忆能力的影响
2 北五味子总木脂素对衰老小鼠脑组织中Ub蛋白表达的影响
与空白对照组比较,模型组Ub表达增加(P<0.05);与模型组相比,SCL低、中、高剂量组Ub蛋白表达降低(P<0.05),见图1。
3 北五味子总木脂素对衰老小鼠脑组织中GRP78蛋白表达的影响
与空白对照组比较,模型组GRP78蛋白表达增加(P<0.05);与模型组相比,SCL低、中、高剂量组GRP78蛋白表达降低(P<0.05),见图2。
4 北五味子总木脂素对衰老小鼠脑组织中PDI蛋白表达的影响
与空白对照组比较,模型组PDI蛋白表达增加(P<0.05);与模型组相比,SCL低、中、高剂量组PDI蛋白表达降低(P<0.05),见图3。

Figure 1. The effect of SCL on Ub protein expression of the aging mouse brain tissues detected by Western blotting.1: control group; 2: model group; 3: SCL(L) group; 4: SCL(M) group; 5: SCL (H) group.Mean±SD.n=3. *P<0.05 vs control group; #P<0.05 vs model group.

Figure 2. Effect of SCL on GRP78 protein expression of the aging mouse brain tissue detected by Western blotting.Mean±SD.n=3.*P<0.05 vs control group; #P<0.05 vs model group.
5 北五味子总木脂素对衰老小鼠脑组织中CHOP蛋白表达的影响
与空白对照组比较,模型组CHOP蛋白表达增加(P<0.05);与模型组相比,SCL低、中、高剂量组CHOP蛋白表达降低(P<0.05),见图4。
6 北五味子总木脂素对衰老小鼠大脑皮质Bcl-2、Bax蛋白表达的影响
免疫组化结果显示,空白对照组神经细胞形态正常,细胞核清晰,胞浆丰富;模型组大部分神经细胞变性,神经元体积变小,胞核与胞浆界限模糊,细胞核固缩成三角形或不规则形;与模型组相比,SCL低、中、高剂量组神经细胞形态规则仍有不同程度的神经细胞变性,但变性细胞数量明显减少,见图5、6。

Figure 3. The effect of SCL on PDI protein expression of the aging mouse brain tissue detected by Western blotting.Mean±SD.n=3.*P<0.05 vs control group; #P<0.05 vs model group.
空白对照组神经细胞细胞浆内可见棕黄色颗粒,Bcl-2蛋白为阳性表达;模型组神经细胞胞浆内棕黄色颗粒含量降低,Bcl-2蛋白为阴性表达;与模型组相比,SCL低、中、高剂量组神经细胞细胞浆内可见棕黄色颗粒,Bcl-2蛋白为阳性表达,见图5。
空白对照组神经细胞胞浆内无棕黄色颗粒,Bax蛋白为阴性表达;模型组神经细胞胞浆内可见棕黄色颗粒,Bax蛋白为阳性表达;与模型组相比,SCL低、中、高剂量组神经细胞胞浆内无棕黄色颗粒,Bax蛋白为阴性表达,见图6。
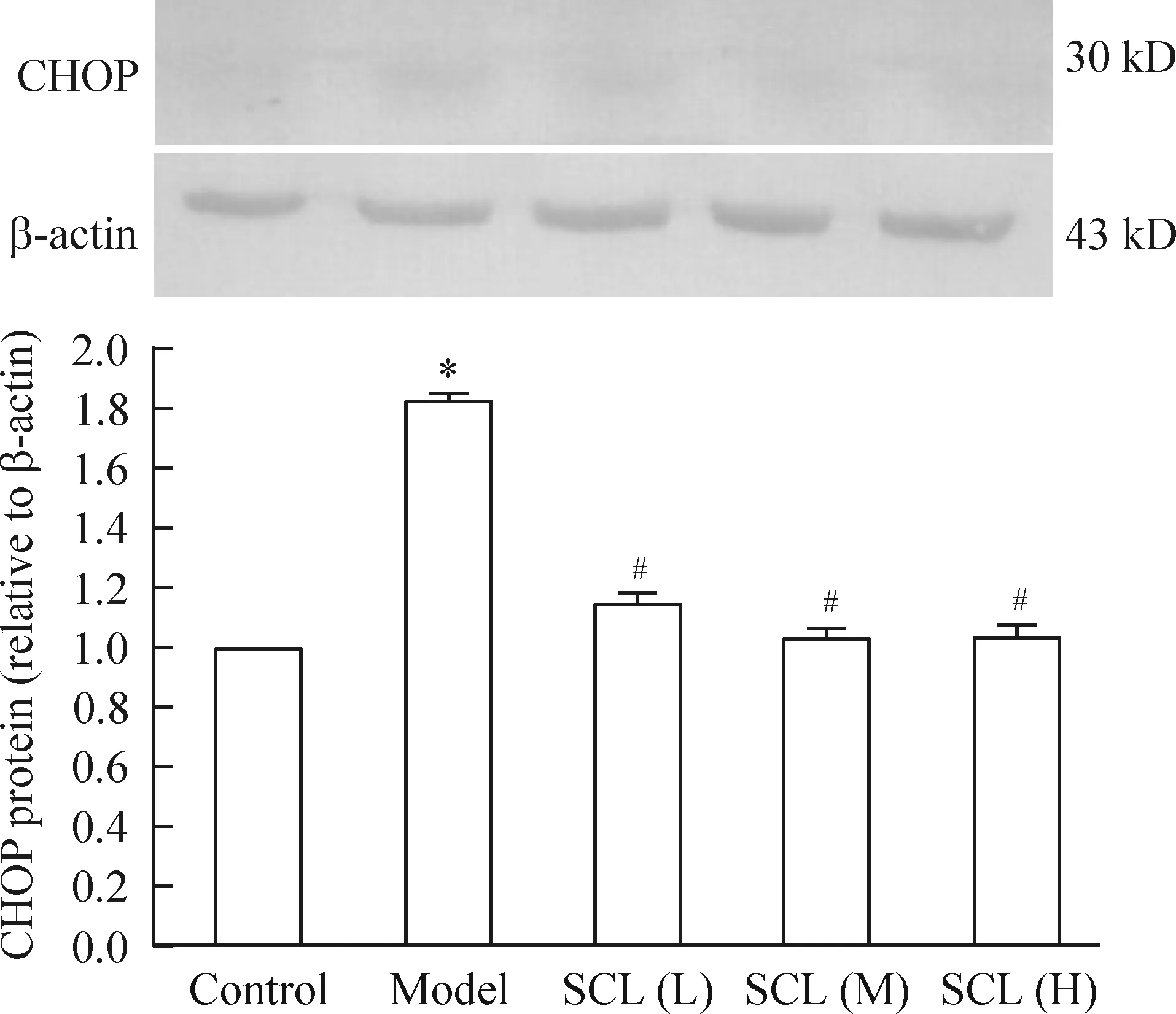
Figure 4. The effect of SCL on CHOP protein expression of the aging mouse brain tissues detected by Western blotting.Mean±SD.n=3.*P<0.05 vs control group; #P<0.05 vs model group.
7 北五味子总木脂素对衰老小鼠脑组织中Bcl-2和Bax蛋白表达的影响
与空白对照组相比,模型组线粒体抗凋亡蛋白Bcl-2表达降低(P<0.05),促凋亡蛋白Bax表达增加(P<0.05),Bcl-2/Bax比值降低(P<0.05);与模型组相比,SCL低、中、高剂量组Bcl-2蛋白增加(P<0.05),Bax蛋白表达降低(P<0.05),Bcl-2/Bax比值升高(P<0.05),见图7。
讨 论
北五味子药用其成熟、干燥的果实,其中含有木脂素、多糖、挥发油、脂肪酸和维生素等活性成分,具有增强心血管功能、抗氧自由基及免疫增强作用。本实验室前期研究结果显示,北五味子具有明显的镇静、催眠、促睡眠作用,并对剂量呈现一定依赖性[9-10]。北五味子总木脂素具有抗疲劳、增强学习记忆能力、增强免疫系统功能等作用[11]。五味子油对2型糖尿病大鼠胰岛B细胞具有保护作用[12]。北五味子总木脂素具有抑制过氧化氢(H2O2) 诱导大鼠肾上腺嗜铬细胞瘤PC12细胞凋亡的作用,与线粒体途径凋亡有关[13]。本课题从北五味子延缓衰老方面进行研究。

Figure 5. The effect of SCL on Bcl-2 protein expression of the aging mouse cerebral cortex observed by immunohistochemistry (×400).
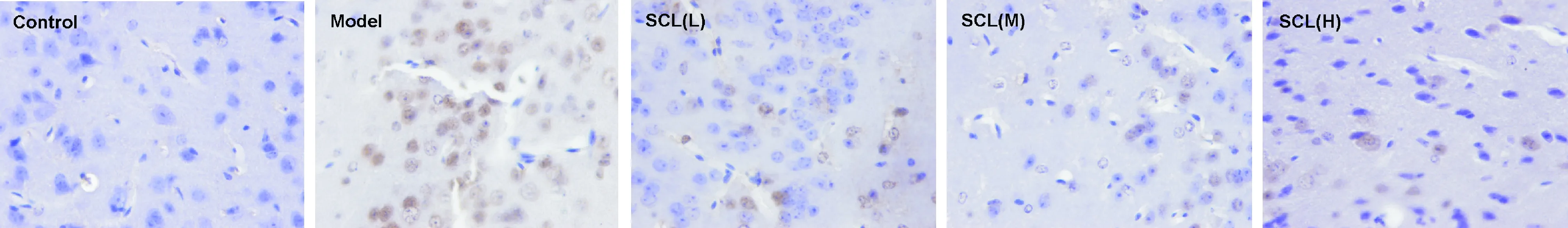
Figure 6. The effect of SCL on Bax protein expression in the aging mouse cerebral cortex observed by immunohistochemistry (×400).
D-半乳糖诱发动物衰老与氧化应激损伤和自由基毒性有关[14]。D-半乳糖可诱发体内自由基蓄积,使蛋白、脂质及核酸过氧化,引起线粒体结构损伤和功能紊乱,出现能量代谢障碍,导致细胞损伤。机体衰老以脑结构和脑功能的改变为重要特征。随着大脑老化,灰色物质萎缩,白色物质功能开始衰退[15]。萎缩现象在前额皮质和海马体处非常明显,前额皮质的功能是帮助人们进行高级思考和推理,而海马体在记忆过程中起着核心作用。学习记忆障碍是脑衰老的一种重要表现。我们的实验结果显示,D-半乳糖诱导小鼠表现为学习记忆能力障碍,形成衰老的行为学改变;通过免疫组化结果,观察到衰老小鼠大脑皮质神经细胞变性,形成衰老的形态学改变。北五味子总木脂素能改善衰老小鼠的学习记忆能力,延缓衰老的发生和发展。
目前研究发现,在衰老相关的疾病中,细胞活性氧生成过多可引起线粒体能量代谢障碍,从而导致一些神经退行性疾病的发生或恶化[16]。线粒体的完整性由Bcl-2超家族的几个成员控制。细胞内存在促凋亡和抗凋亡2个Bcl-2蛋白家族。在凋亡发生时,促凋亡蛋白Bax和Bak二聚化,从而插入线粒体外膜,启动内源性凋亡信号通路,使得生物膜上形成孔洞。抗凋亡蛋白Bcl-2可以中断这一相互作用,避免线粒体膜通透化,故细胞凋亡的发生取决于Bcl-2 和Bax的相对浓度,即Bcl-2 /Bax 值。免疫组化结果观察到衰老小鼠神经细胞胞浆中抗凋亡蛋白Bcl-2呈阴性表达,促凋亡蛋白Bax呈阳性表达,SCL低、中、高剂量组神经细胞胞浆中抗凋亡蛋白Bcl-2呈阳性表达,促凋亡蛋白Bax呈阴性性表达。Western blotting结果显示,衰老小鼠Bcl-2蛋白表达水平降低,Bax蛋白表达增加,Bcl-2/Bax降低;SCL低、中、高剂量组Bcl-2蛋白表达水平增加,Bax蛋白表达降低,Bcl-2/Bax增加,这些结果提示北五味子总木脂素通过减少神经细胞凋亡而发挥抗小鼠脑组织衰老的作用。

Figure 7. The effects of SCL on the protein expression of anti-apoptotic protein Bcl-2 and proapoptotic protein Bax, and Bcl-2 /Bax ratio in the aging mouse brain tissue detected by Western blotting.Mean±SD.n=3.*P<0.05 vs control group; #P<0.05 vs model group.
衰老不仅能引起机体各组织器官功能下降、免疫功能下降易感染以外,还引起阿尔茨海默(Alzheimer)病等相关疾病[17]。在Alzheimer病患者脑中可见β淀粉性蛋白沉积,由于错误折叠或变性蛋白质在神经细胞内堆积而导致中枢神经系统功能障碍,细胞凋亡速率是正常对照组的50多倍。异常蛋白或聚集的错误折叠的蛋白是被泛素化修饰的,所以常常用Ub代表蛋白发生异常或错误折叠[18]。
内质网是真核细胞的重要细胞器,提供了蛋白质合成、折叠、修饰然后分泌到细胞外,或表达于细胞膜表面的主要场所[19]。内质网内新生成蛋白质的正确折叠、成熟和稳定由内质网内的几种驻留分子伴侣来完成,其中包括了GRP78以及几种参与蛋白折叠的酶类,例如硫氧还蛋白超家族成员PDI。在各种生理和病理的情况下,包括缺氧、氧化损伤,都有可能会导致内质网的蛋白折叠负荷和能力的失衡,从而引起未折叠蛋白或错误折叠蛋白在内质网内堆积,此时即发生内质网应激。内质网应激发生时,其腔内的PDI蛋白表达增加、活化以恢复内质网蛋白折叠能力。内质网应激是进化保守机制,其整合了信号转导途径未折叠蛋白反应(unfolded protein response,UPR)。哺乳动物细胞的UPR反应是由3个跨膜蛋白所控制,都是存在于内质网膜上的应激感受因子,它们分别是PERK、IRE1和ATF6。UPR首要的作用就是减轻内质网蛋白负担。GRP78在内质网应激时与IRE-1α、PERK、ATF6解离,激活内质网应激信号通路[20]。
内质网应激是细胞对内外刺激的适应性反应。适度内质网应激诱导GRP78 等内质应激蛋白表达上调,有利于内质网处理未折叠蛋白或错误折叠蛋白,并促进钙稳态恢复;持续而严重的内质网应激可明显上调GRP78的表达,诱导内质网应激相关的CHOP 的表达及活化,触发内质网应激相关凋亡途径[21]。CHOP 又称生长停滞及DNA损伤蛋白153是内质网应激特异的转录因子,过量表达的CHOP 由胞浆转位至细胞核,进而通过下调抗凋亡蛋白Bcl-2,导致Bax 从胞浆转位至线粒体,促进细胞凋亡[22],或者诱导促凋亡蛋白Bim表达等途径促进细胞凋亡[23]。另外,CHOP 可通过上调内质网氧化酶1α( ER oxidase 1α,ERO1α) 诱导三磷酸肌醇受体介导的钙释放,激活钙/钙调蛋白依赖性蛋白激酶,进而触发线粒体凋亡途径诱导细胞凋亡。
我们的结果显示,衰老模型组Ub蛋白表达水平增加,内质网应激标志蛋白PDI表达增加,GRP78和内质网途径凋亡调控蛋白CHOP蛋白表达增加,Bcl-2蛋白表达水平降低,Bax蛋白表达增加,Bcl-2/Bax降低;SCL低、中、高剂量组Ub蛋白、PDI 、GRP78和CHOP蛋白表达降低,Bcl-2蛋白表达水平增加,Bax蛋白表达降低,Bcl-2/Bax增加,这提示北五味子总木脂素通过减少内质网应激诱导的凋亡而发挥抗小鼠脑衰老的作用。
细胞凋亡途径分为内源性与外源性,内源性凋亡有线粒体途径和内质网途径2条主要的信号途径。Bcl-2 蛋白家族是线粒体途径凋亡的关键分子,CHOP 被认为是内质网应激介导凋亡的特异性标志。内质网和线粒体之间存在结构和功能上的紧密联系,参与调控内质网到线粒体间通信的因素还需深入研究。
[参 考 文 献]
[1] Donmez G. Sirtuins as possible targets in neurodegenerative diseases[J]. Curr Drug Targets, 2013, 14(6):644-647.
[2] 林红梅, 韩红祥, 李岳桦, 等. 五味子木脂素与生态因子相关性研究[J]. 中国中药杂志, 2013, 38(24):4281-4286.
[3] 谭敦勇, 李楚杰.衰老分子机制研究的某些进展[J]. 中国病理生理杂志, 2000, 16(1):79-83.
[4] 席兴字. 细胞自噬与衰老[J]. 生命的化学, 2011, 31(2):268-271.
[5] Szegezdi E, Logue SE, Gorman AM, et al. Mediators of endoplasmic reticulum stress-induced apoptosis[J]. EMBO Rep, 2006, 7(9):880-885.
[6] 鞠文博. 北五味子总木脂素抗小鼠衰老过程中自噬作用的实验研究[D]. 吉林: 北华大学, 2013.
[7] Orgel LE. The maintenance of the accuracy of protein synthesis and its relevance to aging: a correction [J]. Proc Natl Acad Sci U S A, 1970, 67(3):1476.
[8] Bruce R, Troen MD. The biology of aging[J]. Mt Sinai J Med,2003, 70(1):3-22.
[9] 陈建光, 王春梅, 高晓旭, 等. 一种镇静安神促进睡眠的口服液及其制备方法: 中国, CN103202896A[P].2013-07-17.
[10] 石 绘, 万丽华, 李 贺,等. 北五味子木脂素对小鼠镇静催眠作用的实验研究[J]. 中国老年保健医学, 2012, 10(5):27-28.
[11] 李 贺, 陈建光, 王春梅, 等.一种具有抗衰老作用的保健食品及其制备方法: 中国, CN103141833A [P],2013-06-12.
[12] 安丽萍,王英平,王春梅,等. 五味子油对链脲佐菌素诱导的2型糖尿病大鼠的影响[J]. 吉林大学学报:医学版, 2012, 38(1):84-88.
[13] 姜恩平, 吴金义, 陈建光. 北五味子总木脂素对H2O2诱导PC12 细胞凋亡的影响[J]. 吉林大学学报:医学版, 2009, 35(5):466-469.
[14] 曲 娴,方文娟,李 冰,等. 维生素E对D- 半乳糖诱致衰老小鼠脑抗氧化能力、钙稳态和线粒体DNA(mtDNA)损伤的影响[J]. 中国病理生理杂志, 2008,24(3):523-526.
[15] 崔玉丰,张博妍,张 瑞,等. 白藜芦醇对衰老模型小鼠脑组织形态学和氧化应激水平的影响[J]. 卫生研究, 2013, 42(6):996-998.
[16] 叶 薇, 陈赛慧, 郝东杰. 衰老过程中线粒体顺乌头酸酶活性变化对能量合成的影响[J]. 中国病理生理杂志, 2013, 29(7):1275-1282.
[17] 贺改英,徐 颖,吴丽莉,等. 银杏酮酯对衰老大鼠海马炎症相关细胞因子的调节作用[J]. 中国中药杂志, 2012, 37(14):2130-2134.
[18] Bhutia SK, Dash R, Das SK, et al. Mechanism of autophagy to apoptosis switch triggered in prostate cancer cells by antitumor cytokine melanoma differentiation-associated gene 7/interleukin-24 [J]. Cancer Res, 2010, 70(9):3667-3676.
[19] Görlach A, Klappa P, Kietzmann T. The endoplasmic reticulum: folding, calcium homeostasis, signaling, and redox control[J]. Antioxid Redox Signal, 2006, 8(9-10):1391-1418.
[20] Lee AS. GRP78 induction in cancer: therapeutic and prognostic implications[J]. Cancer Res, 2007, 67(8):3496-3499.
[21] 陶天琪,王晓礽,徐菲菲,等. MR-1 通过抑制PERK/Nrf2 途径减轻缺氧/复氧诱导的心肌细胞凋亡[J]. 中国病理生理杂志, 2014, 30(2):193-202.
[22] 姚树桐,秦树存. 内质网应激在动脉粥样硬化发生、发展和防治中的作用[J]. 中国病理生理杂志, 2014, 30( 2):364-368.
[23] 夏 珍,李菊香,丁 浩,等. 内质网应激在Bim 介导缺氧致心肌细胞凋亡中的作用[J]. 中国病理生理杂志, 2013, 29(12):2121-2127.
猜你喜欢
中山大学学报(自然科学版)(中英文)(2021年3期)2021-05-26
现代临床医学(2021年1期)2021-01-26
山东医药(2021年28期)2021-01-11
南风(2020年13期)2020-06-05
中国粮油学报(2019年4期)2019-07-12
中成药(2018年12期)2018-12-29
中成药(2017年3期)2017-05-17
中成药(2017年3期)2017-05-17
药学研究(2015年11期)2015-12-19
——疾病防治的新靶标
中国药理学通报(2015年3期)2015-06-09
