
宫内感染致脑损伤对仔鼠认知发育和海马神经发生的影响*
2014-08-13 12:39江佩芳杨翠微顾伟忠袁天明俞惠民
中国病理生理杂志 2014年11期
江佩芳, 朱 涛, 杨翠微, 高 峰, 顾伟忠, 袁天明, 俞惠民△
(1浙江大学医学院附属儿童医院神经内科,浙江 杭州 310003; 2浙江大学医学院附属邵逸夫医院ICU,浙江 杭州 310016)
妊娠期宫内感染导致的胎儿炎症反应综合征是造成早产和早产儿脑白质损伤(white matter damage, WMD)的最常见原因,是儿童认知功能发育障碍的一个重要危险因素[1]。WMD临床病理特点主要是脑白质出现弥漫的反应性星形细胞胶质化、成髓鞘少突胶质细胞损伤和脑室周围白质软化(periventri-cular leukomalacia, PVL)[2]。它将会影响神经系统发育,造成语言、视觉、空间、记忆和注意力等认知功能损害。因此,宫内感染与儿童认知功能发育密切相关。
炎症反应犹如一把双刃剑,其过程中释放的一些细胞因子可以增加血脑屏障通透性,引起脑水肿,加重脑损伤,抑制神经发生[3]。还有一些细胞因子可以刺激神经发生,促进神经干细胞增殖。研究发现缺氧缺血能诱导海马神经前体细胞增殖,神经干细胞向损伤区域迁移,分化为神经元、星形胶质细胞和少突胶质细胞;并功能性整合到海马神经环路中,促进神经轴突、树突发育,以及突触生成和功能性修饰[4]。虽然缺氧缺血可以诱导海马神经发生,但是宫内感染与神经发生的相关性研究未见报道。我们旨在确证宫内感染致仔鼠脑损伤对认知发育和海马神经发生的影响,为这类患儿的病理学和治疗学研究提供理论依据。
材 料 和 方 法
1 实验动物
清洁级健康成年Sprague-Dawley孕鼠(胎龄15 d),购自浙江省医学科学院实验动物中心,体重400~450 g,按随机数字表法分为造模组(8只)和对照组(7只)。繁殖后的2组新生雄性大鼠按随机数字表法分为宫内感染组(35只)和对照组(35只)。实验期间,室温控制在(24±1) ℃,湿度控制在60%左右,大鼠自由饮食、饮水,昼夜周期12 h/12 h(光照时间为8:00-20:00)。实验过程中人道主义对待实验动物,尽量减少实验动物使用量,设法在处死实验动物时使其少受痛苦。
2 方法
2.1宫内感染模型制备 参照本课题组先前的成功造模方法[5],取胎龄15 d 的Sprague-Dawley孕鼠15只,随机分为2组:(1) 造模组(n=8)予腹腔内注射2%戊巴比妥钠2 mL/kg,麻醉后于左右两侧宫颈内各注射大肠杆菌(E.coli)稀释液0.2 mL,术后置于清洁笼内,常规喂养;(2) 对照组(n=7)于麻醉后左右两侧宫颈内各注射无菌生理盐水0.2 mL。
根据文献[6]的标准,凡宫内感染组鼠的子宫及胎盘内血管充血、水肿,并见大量中性粒细胞浸润者判断为宫内感染。
待孕鼠自然分娩(胎龄约20 d),2组雄性仔鼠按随机数字表法分为宫内感染组(E.coli)和对照组(control)。
2.2Morris水迷宫实验 Morris水迷宫DMS-2系统由浙江大学紫金港动物房中心实验室提供,包括一个圆形不锈钢水池,直径为120 cm,高60 cm。池壁上标有4个不同符号,将水池等分为4个象限,代表4个入水点。目标象限的中央放置一高度50 cm直径10 cm的圆形隐藏平台,平台低于水面1 cm,水温保持在(22±0.5) ℃。预先在水池中注入清水,加入炭素墨水使池水变为不透明的黑色以隐藏平台并便于摄像。迷宫上方安置带有显示系统的摄像机,计算机自动跟踪计时并同步记录大鼠的游泳轨迹,Morris水迷宫数据采集和分析软件记录相关数据及图像结果。整个实验期间水池、光源、鼠笼等实验室各物件的位置保持不变,实验在隔音的房间内进行。
2.2.1迷宫适应训练和游泳能力检验 正式开始实验前1 d,撤去平台,选择池壁一固定点,让大鼠面向池壁入水,自由游泳2 min,并录下游泳轨迹。目的是让大鼠适应环境,并对大鼠游泳能力进行初步检验,对明显异常者予以剔除。
2.2.2定位航行实验 用于检测大鼠的近期学习记忆能力。2组28日龄大鼠(n=10)连续接受5 d训练,每天将大鼠面向池壁分别从4个入水点放入水中,每次实验以120 s为限,记录动物寻找并爬上平台所需时间即逃避潜伏期(escape latency, EL),以爬上平台所需时间作为学习和记忆成绩。若设定时间内未找到平台,计算机停止跟踪,记录时间为120 s,未找到平台的动物由实验者将其引上平台。每次训练间隔30 s,即做好一次训练后让大鼠在平台上休息停留30 s后再继续下一次训练。当天训练完成后,迅速将动物洗净擦干,置于加热器旁,以防动物低体温。计算每天2组大鼠4次逃避潜伏期的平均值。
2.2.3远期记忆保持实验 定位航行实验结束后,休息5 d,即水迷宫的第10天,再对全部大鼠进行一次水迷宫测试,方法同前,测试其远期记忆保持能力。
2.3标本制备 2组仔鼠随机分组,每组每时点5只。从8:00开始给1 d龄大鼠腹腔注射5-溴-2’-脱氧尿嘧啶核苷(bromodeoxyuridine, BrdU; 50 mg/kg, Sigma),每天2次,连续注射3 d,标记海马S期先祖细胞,以反映齿状回细胞的增殖情况。另外于生后1 d龄开始连续腹腔注射相同剂量BrdU 7 d,待28 d龄时行细胞存活分析。大鼠断头处死,取出脑组织先进行大体解剖学观察,再将脑组织置于4%多聚甲醛中固定24~48 h后石蜡包埋,固定后冠状切面连续切片每隔3张取1张切片组成1套,每套切片约5张,每片厚4 μm,然后对上述切片进行TUNEL和免疫组化分析。
2.4TUNEL分析 对2组标本进行TUNEL染色,实验操作参照试剂说明书进行。石蜡切片经二甲苯脱蜡,乙醇水化,置3% H2O2中孵育15 min,蛋白酶K消化液作用20 min,加入TUNEL反应混合物,于37 ℃烤箱中孵育60 min,滴加转化剂POD,于37 ℃烤箱中孵育30 min,DAB显色20 min,乙醇脱水,二甲苯透明,中性树脂封片。显微镜下观察切片中细胞形态,TUNEL染色阳性的凋亡细胞呈棕色。倒置显微镜下随机取5个视野观察阳性细胞并计数,取平均值,计算出细胞凋亡指数(apoptosis index,AI)=(凋亡细胞数÷细胞总数)×100%。
2.5Caspase-3免疫组化分析 按照SABC免疫组化试剂盒进行操作。切片蒸馏水新鲜配制3%H2O2室温10 min,PBS洗3次。滴加正常山羊血清封闭液,室温20 min甩去多余液体不洗。滴加1∶100稀释的Ⅰ抗,37 ℃1 h,4 ℃过夜。复温l h,PBS洗3次,滴加生物素化Ⅱ抗,37 ℃ 20 min,PBS洗3次。滴加SABC,37 ℃ 30 min,PBS洗4次,DAB显色。乙醇梯度脱水,二甲苯透明,中性树酯封片。阴性对照用PBS代替Ⅰ抗。倒置显微镜下随机取5个视野观察阳性细胞并计数,取平均值。
2.6BrdU免疫组化 切片常规脱蜡、透明至水化。切片置于0.01 mol/L枸橼酸钠溶液(pH 6.0)高温高压抗原修复1 min,0.025% PBST(含Triton X-100)漂洗3次。3% H2O2室温下消化10 min以灭活内源性过氧化物酶,0.025% PBST漂洗3次。羊血清37 ℃封闭20 min。以小鼠抗BrdU抗体(1∶50, Santa Cruz)37 ℃孵育1 h;0.025% PBST漂洗3次。滴加Envision 检测试剂,37 ℃孵育1 h;0.025% PBST漂洗3次。室温下DAB显色,镜控2 min。苏木素轻度复染,脱水、透明、封片。胞浆内呈棕黄色染色为BrdU阳性细胞,选取海马DG区,200倍视野下计数每0.01 mm2面积中BrdU阳性细胞数量,每张切片计数5个视野,取平均值。
3 统计学处理
实验数据以均数±标准误(mean±SEM)表示,采用SPSS 17.0统计学软件进行分析。细胞增殖分析、TUNEL分析和caspase-3免疫组化分析采用双因素方差分析(two-way ANOVA),以Tukey法进行组间的两两比较;定位航行实验和远期记忆保持实验采用重复测量方差分析(one-way repeated measures ANOVA),以Tukey法进行组间的两两比较;细胞存活分析采用独立样本t检验。以P<0.05为差异有统计学意义。
结 果
1 宫内感染致仔鼠脑损伤的大体解剖学观察
空白对照组双侧大脑对称,饱满,脑组织表面血管清晰,脑区结构完整。宫内感染组双侧大脑对称,脑组织轻度萎缩,表面血管增粗、紊乱,脑区结构完整,未见坏死、液化、空洞现象,见图1。

Figure 1. Effect of intrauterine infection on anatomy observation of brain.
2 宫内感染对仔鼠海马神经细胞凋亡的影响
TUNEL染色分析显示宫内感染组TUNEL阳性细胞数较对照组显著增加(P<0.05),另一凋亡标志物caspase-3染色显示caspase-3阳性细胞数较对照组显著增加(P<0.05),见图2。
3 宫内感染对仔鼠认知功能的影响
3.1游泳能力的检测 正式开始实验前1 d,撤去平台,选择池壁一固定点,让大鼠面向池壁入水,自由游泳2 min,并录下游泳轨迹。各组大鼠第l d通过测试,全部大鼠游泳能力均正常。
3.2定位航行实验 宫内感染组游泳轨迹较对照组杂乱,重复测量方差分析显示组别因素(group)、时间因素(day)以及时间和组别(day*group)的交互作用均具有统计学意义(P<0.01)。同一时间组间比较结果显示,宫内感染组第1天到第4天的逃避潜伏期均显著延长(P<0.05),第5天2组的逃避潜伏期无统计学差异(P>0.05),说明宫内感染可以影响仔鼠的近期学习记忆能力,见图3、表1。
3.3远期记忆保持实验 远期记忆保持实验从第10 d开始,宫内感染组游泳轨迹较对照组杂乱,重复测量方差分析显示组别因素(group)、时间因素(day)以及时间和组别(day*group)的交互作用均具有统计学意义(P<0.01)。宫内感染组逃避潜伏期较对照组显著延长(P<0.05),说明宫内感染可以影响仔鼠的远期学习记忆能力,见图4、表2。

Figure 3. Effect of intrauterine infection on offspring’s learning and memory ability. Figure shows the trace of place-navigation test from control group and E. coli group at different time points.

表1 2组大鼠定位航行试验的逃避潜伏期比较

Figure 4. Effect of intrauterine infection on offspring’s long-term memory ability. Figure shows the trace of long-term memory test from control group and E. coli group at different time points.
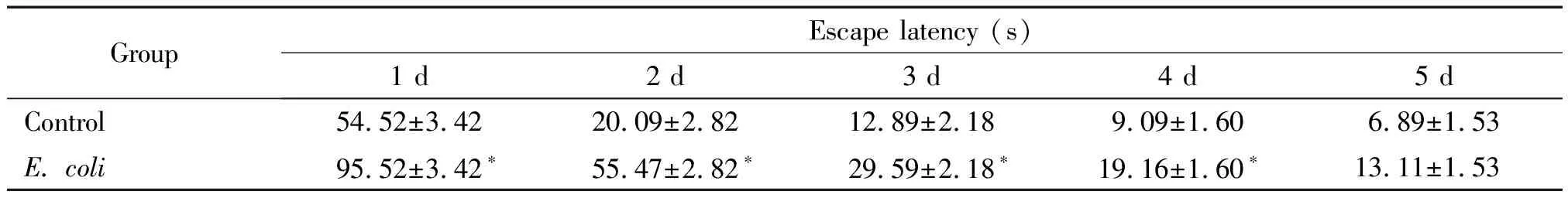
表2 2组大鼠远期记忆保持实验的逃避潜伏期比较
4 宫内感染对仔鼠海马神经发生的影响
细胞增殖分析表明宫内感染组仔鼠于3、7和14日龄时海马齿状回BrdU阳性细胞数较对照组显著增加(P<0.05),于7日龄时达高峰。细胞存活分析显示28日龄时宫内感染组与对照组仔鼠海马齿状回BrdU阳性细胞数无统计学差异(P>0.05),见图5。
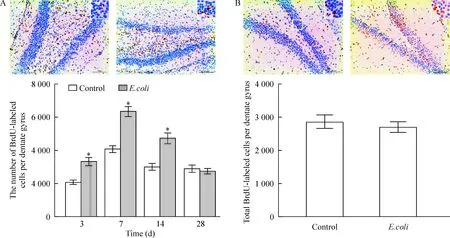
Figure 5. Effect of intrauterine infection on proliferation and survival of newborn cells in the dentate gyrus. The cells with BrdU (brown stained) that clearly localized in the nucleus were counted as BrdU-labeled cells. A: representative microphotographs of BrdU-labeled cells and quantification of BrdU-labeled cells during postnatal 28 days in the control and E. coli rats; B:representative microphotographs of BrdU-labeled cells and quantification of total BrdU-labeled cells at postnatal day 28 in the control and E. coli rats. Scale bars=200 μm. *P<0.05 vs control.
讨 论
学习记忆是大脑的高级功能,学习记忆及认知功能减退是脑损伤导致智力低下的一个重要标志。海马结构完整性和突触可塑性对空间学习记忆能力的正常发育非常重要[7]。研究认为新生期缺氧缺血性脑损伤主要累及皮层、海马,出现神经元肿胀变性、大量核固缩、核碎片、空泡形成和排列紊乱,进而造成近期、远期学习记忆功能缺陷,遗留神经系统后遗症[8]。虽然缺氧缺血性脑损伤与认知功能缺陷的报道较多,但关于宫内感染造成脑损伤与认知发育的研究罕见报道。
本研究发现宫内感染可导致仔鼠脑组织萎缩,但脑区结构保持完整。结合前期研究发现宫内感染造成脑白质结构稀疏、呈筛网状改变,少数出现凝固性坏死和囊状病变。由此我们推断脑白质可能是宫内感染脑损伤中最易受累的部位。星形胶质细胞是中枢神经系统的基本骨架,合成和分泌众多的神经营养因子、趋化因子和细胞因子,参与中枢神经系统信息转导与免疫反应[9]。轻度星形胶质细胞活化被证实有神经保护作用,而过度活化可导致星形胶质细胞凋亡,垂死的星形胶质细胞呈现神经毒性作用,进一步杀死邻近细胞[10]。因此,我们推测星形胶质细胞在脑白质损伤的病理生理机制中扮演了重要角色,宫内感染后过度活化的星形胶质细胞可影响海马神经元的营养支持及细胞功能,造成神经细胞体积缩小、数量减少并最终导致大脑萎缩。
宫内感染脑白质损伤的病理生理机制复杂,与认知发育障碍的相关机制尚未完全阐明,有学者认为细胞凋亡可能是系统性炎症反应中的重要机制之一[11]。本研究发现宫内感染后仔鼠海马神经元大量凋亡,结合前期研究发现宫内感染后仔鼠脑内细胞因子,一氧化氮合成及诱导型一氧化氮合酶表达增加,我们推断宫内感染促发一系列炎症级联反应,诱发炎症风暴,促进神经细胞凋亡。神经细胞凋亡导致海马神经元数量减少、功能受累,海马神经轴突、树突发育不良、突触生成受阻和功能性修饰受累[12]。所以,宫内感染后仔鼠的近期及远期记忆保持能力均显著下降,可能与神经细胞凋亡密切相关。
本研究还发现随着游泳训练时间延长,2组逃避潜伏期的差距逐渐缩短,提示宫内感染脑损伤后神经细胞可能存在一定的自身修复能力。多年来研究者们一直认为神经元不能再生,但随着干细胞研究的深入,发现成年哺乳动物脑内存在具有多向分化潜能的神经干细胞和持续的神经发生现象,为补充因损伤、衰老而丧失的神经元或胶质细胞提供来源[13-14]。虽然炎症损伤与神经发生的关系目前还不明确,但Bakirci等[15]报道在脓毒症模型中发现了神经发生现象。本研究也发现宫内感染导致海马神经元凋亡的同时也能促进神经发生。我们推测炎症物质在炎症亚急性期后作为神经营养因子,作用于神经发生微环境,调节神经干细胞增殖、迁移和分化,发挥神经保护作用,这种神经自身修复的潜能可能是脑白质损伤后认知功能障碍得到一定程度改善的重要物质基础。但是我们发现早期增殖的细胞大部分不能持续存活,推测可能没有适当的微环境刺激,神经发生得不到适当调节,这些增殖的神经干细胞互相竞争,一部分因营养耗竭而死亡,一部分不能正常分化为成熟的星形胶质细胞和神经元,这可能是宫内感染后仔鼠出现认知发育障碍后遗症的重要原因。
虽然宫内炎症反应造成脑损伤后可以促进神经发生,但是炎症反应何时以何种强度刺激有利于神经发生,以及如何通过调控炎症反应来影响损伤组织修复和神经功能恢复等问题还不明确。目前研究发现cAMP反应元件结合蛋白(cAMP response element binding protein, CREB)是一种重要的核转录因子,在神经元再生、突触形成及学习记忆等方面具有重要的调节作用,是细胞内多种信号通路的一种关键成分。动物实验发现CREB基因敲除小鼠多死于围生期,与中枢神经系统神经元大量缺失有关。研究还发现PI3K/Akt信号通路可参与调节神经干细胞的增殖、神经前体细胞迁移、分化以及突触形成,保持神经干细胞的维持及其分化的平衡。但是PI3K/Akt信号通路与CREB是否共同参与宫内感染后的神经发生,这三者之间的关系目前尚未明确。因此,宫内感染后神经发生的相关分子机制研究是本课题组今后工作的研究方向。对这一过程的探讨,无疑将为研究宫内感染后脑损伤患儿认知发育障碍的治疗提供新的手段。
[参 考 文 献]
[1] Bashiri A, Burstein E, Mazor M. Cerebral palsy and fetal inflammatory response syndrome: a review[J]. J Perinat Med, 2006, 34(1):5-12.
[2] Buser JR, Maire J, Riddle A, et al. Arrested preoligodendrocyte maturation contributes to myelination failure in premature infants[J]. Ann Neurol, 2012, 71(1):93-109.
[3] Zonis S, Ljubimov VA, Mahgerefteh M, et al. p21Ciprestrains hippocampal neurogenesis and protects neuronal progenitors from apoptosis during acute systemic inflammation[J]. Hippocampus, 2013, 23(12):1383-1394.
[4] Bruel-Jungerman E, Davis S, Rampon C, et al. Long-term potentiation enhances neurogenesis in the adult dentate gyrus[J]. J Neurosci, 2006, 26(22):5888-5893.
[5] Jiang P, Sun Y, Zhu T, et al. Endogenous neurogenesis in the hippocampus of developing rat after intrauterine infection[J]. Brain Res, 2012, 1459:1-14.
[6] Yuan TM, Yu HM, Gu WZ, et al. White matter damage and chemokine induction in developing rat brain after intrauterine infection[J]. J Perinat Med, 2005, 33(5):415-422.
[7] Ouchi Y, Banno Y, Shimizu Y, et al. Reduced adult hippocampal neurogenesis and working memory deficits in theDgcr8-deficient mouse model of 22q11.2 deletion-associa-ted schizophrenia can be rescued by IGF2[J]. J Neuro-sci, 2013, 33(22):9408-9419.
[8] Volpe JJ. Neonatal encephalopathy: an inadequate term for hypoxic-ischemic encephalopathy[J]. Ann Neurol, 2012, 72(2):156-166.
[9] Li X, Cudaback E, Breyer RM, et al. Eicosanoid receptor subtype-mediated opposing regulation of TLR-stimulated expression of astrocyte glial-derived neurotrophic factor[J]. FASEB J, 2012, 26(7):3075-3083.
[10] 张 运,王莉莉,赵 茜,等. GRK5对大鼠星形胶质细胞活化的作用及其机制研究[J].中国病理生理杂志, 2014, 30(4):577-583.
[11] Adib-Conquy M, Cavaillon JM, Jacqmin S, et al. CD24-triggered caspase-dependent apoptosis via mitochondrial membrane depolarization and reactive oxygen species production of human neutrophils is impaired in sepsis[J]. J Immunol, 2014, 192(5):2449-2459.
[12] 行妍妍,汪军兵,谢 赛,等. 姜黄素对放线菌素D/TNF-α诱导的PC12细胞和大鼠海马神经元损伤的影响[J].中国病理生理杂志, 2012, 28(10):1746-1750.
[13] Birch AM, McGarry NB, Kelly AM. Short-term environmental enrichment, in the absence of exercise, improves memory, and increases NGF concentration, early neuronal survival, and synaptogenesis in the dentate gyrus in a time-dependent manner[J]. Hippocampus, 2013, 23(6):437-450.
[14] Gould E, Gross CG. Neurogenesis in adult mammals: some progress and problems[J]. J Neurosci, 2002, 22(3):619-623.
[15] Bakirci S, Kafa IM, Uysal M, et al. Increased adult neurogenesis in the subventricular zone in a rat model of sepsis[J]. Neurosci Lett, 2011, 497(1):27-31.
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
作文周刊·小学二年级版(2022年20期)2022-05-05
神经损伤与功能重建(2020年10期)2020-12-23
神经损伤与功能重建(2020年11期)2020-12-01
创新作文(小学版)(2019年10期)2019-09-25
幸福(2019年21期)2019-08-20
飞碟探索(2015年11期)2015-09-10
中国生化药物杂志(2015年4期)2015-07-07
中国康复(2015年4期)2015-04-10
郑州大学学报(医学版)(2015年2期)2015-02-27
