
出芝温度对灵芝基质物质含量和酶活的影响*
2014-07-08 06:45:03余梦瑶许晓燕韩嘉钰郑林用李文涛
中国食用菌 2014年6期
魏 巍,余梦瑶,许晓燕,韩嘉钰,3,江 南,郑林用, 李文涛,3,罗 霞**
(1.四川省中医药科学院中药细胞与分子生物学实验室,四川 成都 610041; 2.四川省农业科学院,四川 成都 610066; 3.四川大学生命科学学院,四川 成都 610064)
出芝温度对灵芝基质物质含量和酶活的影响*
魏 巍1,余梦瑶1,许晓燕1,韩嘉钰1,3,江 南1,郑林用2, 李文涛1,3,罗 霞1**
(1.四川省中医药科学院中药细胞与分子生物学实验室,四川 成都 610041; 2.四川省农业科学院,四川 成都 610066; 3.四川大学生命科学学院,四川 成都 610064)
研究了25℃、30℃和35℃三个出芝温度条件下,利用人工气候箱袋式栽培灵芝,其基质不同时期-部位的物质含量(总糖、还原糖、水溶性蛋白和游离氨基酸)与木质素酶系酶(漆酶和锰过氧化物酶)、纤维素酶系酶(羧甲基纤维素酶和滤纸酶)和酸性蛋白酶活性变化规律。结果显示,在25℃时,灵芝能够正常出芝,温度高于30℃以后子实体分化受到抑制;且在25℃条件下,不同时期-部位基质中总糖和可溶性蛋白含量低于其它2个温度处理组,而还原糖和游离氨基酸含量高于其它2个温度处理组;同时,不同时期-部位基质中木质素酶系和纤维素酶系酶活力低于其它2个温度处理组,但酸性蛋白酶活力高于另外其它2个温度处理组。提示袋料灵芝栽培时出芝温度控制在25℃左右利于其子实体生长发育。
灵芝;袋料;温度;出芝;酶活
灵芝(Ganodermalucidum)作为1种在民间及临床应用广泛且具有极高药用价值的名贵药用真菌,被《中华人民共和国药典》(2010年版)与《美国草药药典与治疗概要》收录,其现代研究、开发与应用已受到国内外诸多学者的重视[1]。
温度作为灵芝生长发育过程中相当重要的环境因素,目前研究仅限于其对菌丝生长速度、子实体产量等农艺指标的影响[2-4],尚未见出芝温度对灵芝营养代谢影响的深入研究报道。
在现有食(药)用菌营养代谢变化规律的研究中,往往将基质内菌丝体或子实体视为1个整体,仅按照不同时间进行取样分析[5],而忽视了菌丝体和子实体不同部位因生长时间的不同、发育阶段的不同而导致的分化,使得研究结果存在一定的局限性。
本论文采用时空模式的研究思路,研究了出芝温度对灵芝不同时期-部位基质的物质含量和酶活力的影响,为深入探究灵芝袋料栽培过程中最适出芝温度条件及其机理奠定了基础。
1 材料与方法
1.1 供试菌株
灵芝ZL13,由四川省中医药科学院细胞与分子生物学实验室提供。
1.2 实验材料及试剂
木屑、棉籽壳、玉米芯、麦麸、石灰和石膏购自农资市场,ABTS和木聚糖购自Sigma公司,其余试剂均为国产分析纯。
1.3 供试培养基
1.3.1 母种培养基配制
土豆200 g、葡萄糖20 g、琼脂20 g,蒸馏水 1 000 mL,pH自然。
1.3.2 原种培养基配制
玉米粒99%、石膏1%,pH自然,含水量60%左右。
1.3.3 栽培培养基配制
木屑15%、棉籽壳60%、玉米芯15%、麦麸8%、石灰1%、石膏1%,pH5~6,含水量60%左右。将木屑、麦麸、玉米芯、石灰和石膏混合均匀,掺入预湿24 h的玉米芯中拌匀。选用23 cm×42 cm×0.03 cm的聚乙烯袋,每袋装湿料 2 000 g,121℃高压灭菌2.5 h,冷却至室温后每袋接入玉米粒原种10粒。
1.4 培养条件及栽培方法
菌丝生长阶段的培养条件为25℃,避光,湿度50%~60%;出芝阶段培养条件为光照300 lx,湿度90%,温度处理分25℃、30℃和35℃进行。所有催蕾及出芝均在人工气候箱内进行。
1.5 采样方法及粗酶液制备
把灵芝生殖生长期分为5个阶段:菌丝满袋(时期1)、原基期(时期2:灵芝刚长出白色球状子实体)、菌蕾期(时期3:白色球状子实体开始分化,有突起产生)、开片期(时期4:灵芝为肾形,有淡黄色边缘)、子实体成熟期(时期5:弹射孢子)。将培养料均分为4部分,分别标记为Ⅰ、Ⅱ、Ⅲ和Ⅳ,具体采样方法如表1所示。

表1 取样表
准确称取同时期同一待测样品2份,每份5 g,将其中1份称好的样品放入50 mL三角瓶中,加蒸馏水25 mL,在40 ℃恒温水浴锅内浸提2 h,3层纱布过滤,10 000 r·min-1离心10 min,上清液即为粗酶液,用于测定酶活;另外1份在80℃恒温干燥箱内烘至恒重,用于确定基质干重。设置3个重复。
1.6 基质中物质含量的测定
采用3,5-二硝基水杨酸(DNS)比色法测定总糖[6]和还原糖的含量;采用考马斯亮蓝法测定可溶性蛋白含量;采用茚三酮显色法测定游离氨基酸含量。
1.7 基质中胞外酶活性的测定
漆酶活力采用ABTS法测定[7];锰过氧化物酶活力采用Mn(Ⅱ)氧化法测定[7];纤维素酶和滤纸酶活力采用3,5-二硝基水杨酸(DNS)比色法测定[8];酸性蛋白酶活性采用福林-酚试剂法测定[9]。
漆酶活力单位定义为每分钟催化1 μmolABTS氧化所需要的酶量;锰过氧化物酶活力单位定义为每分钟使吸光度值增加0.1的酶量;纤维素酶和滤纸酶活力单位定义为每分钟催化生成1 mg葡萄糖所需要的酶量;酸性蛋白酶活力单位定义为每克干料1 min内水解酪蛋白产生1 μg酪氨酸所需的酶量。
2 结果与分析
2.1 不同出芝温度对灵芝子实体生长发育的影响
在本研究中,灵芝生殖生长期均在人工气候箱内进行,以便能够精准地控制灵芝出芝温度。灵芝菌丝满袋后,将菌袋移入3个温度分别设定为25℃、30℃和35℃的人工气候箱内(无昼夜温差)。研究结果表明,在25℃培养条件下,灵芝能够正常出芝;当温度为30℃时,灵芝能出原基,但是由于培养温度稍高,灵芝原基很快干缩,不能正常分化开片;在35℃时,灵芝则不能出原基。
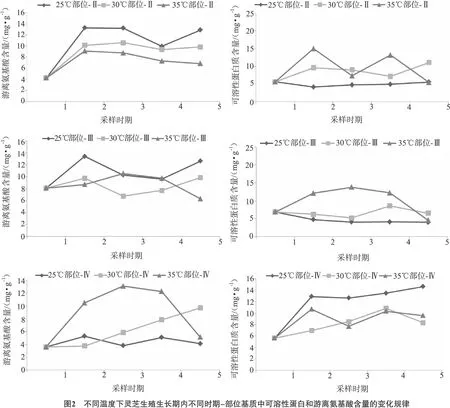
2.2 不同时期-部位基质中总糖和还原糖含量的变化规律
不同出芝温度下,灵芝生殖生长期内不同时期和部位基质中总糖和还原糖含量变化规律如图1所示。在不同培养温度下,不同部位基质中总糖含量差异不大。当出芝温度为25℃和30℃时,部位-Ⅰ、部位-Ⅱ和部位-Ⅳ基质中总糖含量变化规律相同,即先升高后下降,然而部位-Ⅲ中总糖含量变化趋势却与其他部位相反。在35℃培养条件下,不同部位基质中总糖含量一直呈升高趋势。
图1结果显示,温度对基质中还原糖含量的影响较大,在25℃培养条件下,各部位基质中还原糖含量始终高于30℃和35℃处理组。在灵芝的生殖生长期内,30℃和35℃对基质中还原糖含量的影响基本相同,即还原糖含量都维持在较低水平,无较大波动。
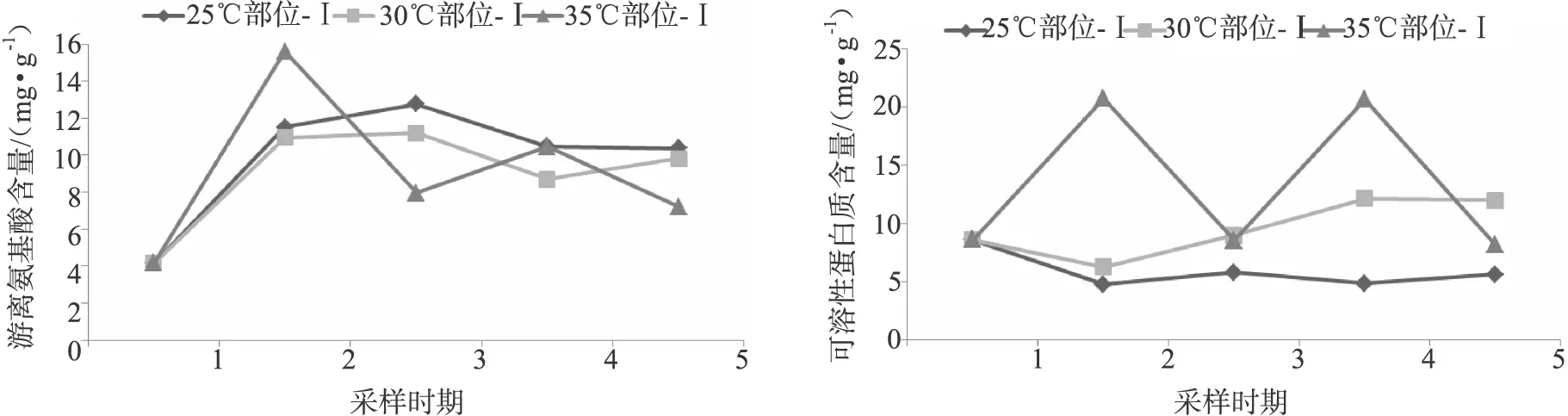
2.3 不同时期-部位基质中可溶性蛋白质和游离氨基酸含量的变化规律
基质中可溶性蛋白质和游离氨基酸含量在不同出芝温度下各时期及各部位的变化规律如图2所示。
从图2可以看出,可溶性蛋白质含量总体呈现下降趋势,并且当培养温度为25℃时,不同部位基质中可溶性蛋白质含量始终低于另外2个处理组。温度对基质各部位中游离氨基酸含量变化趋势影响不明显,在不同温度处理下,游离氨基酸含量从满袋期至原基时期呈升高趋势,其含量均在原基期出现1个峰值然后略有下降。由图2可知,25℃时,不同时期和部位基质中游离氨基酸含量总体高于其他2个温度处理。
2.4 不同时期-部位基质中木质素酶系酶活的变化规律
2.4.1 不同温度对漆酶活性变化规律的影响
灵芝生殖生长阶段温度对不同时期和部位基质中漆酶活性变化规律如图3所示。

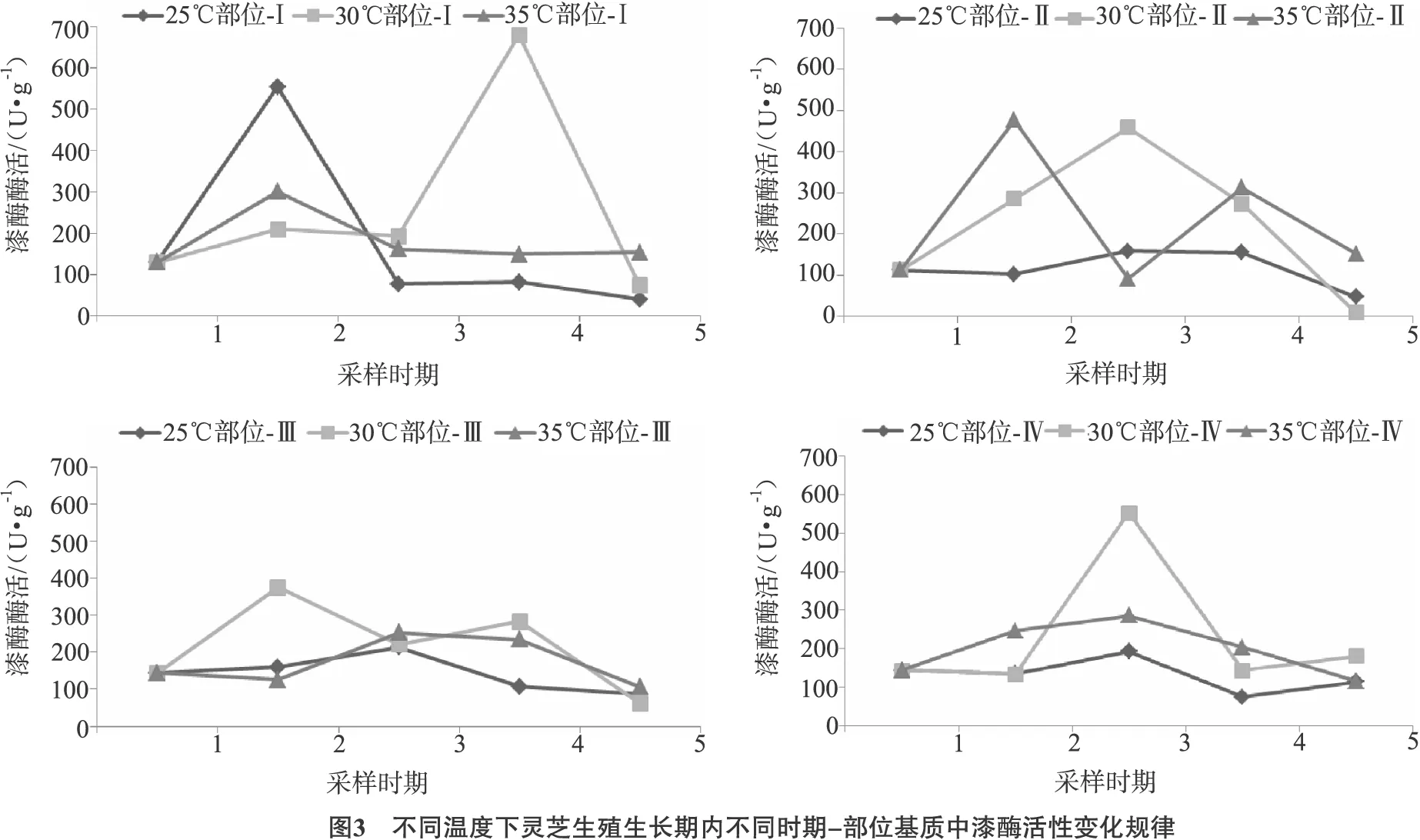
总体来看,生殖生长阶段漆酶活性先经历1个上升阶段最终呈下降趋势,温度对漆酶活性峰值出现的时期有一定影响。25℃时,基质部位-Ⅰ的漆酶酶活原基期出现1个高峰,随后急剧下降,并稳定在较低水平;30℃时,漆酶活性总体处于较高水平,部位-Ⅰ处漆酶酶活在开片期(时期4)达到最高;35℃时,漆酶活性的峰值则出现在菌蕾期。
2.4.2 不同温度对锰过氧化物酶活性变化规律的影响
不同温度对锰过氧化物酶活性变化规律的影响情况见图4。

从图4可以看出,锰过氧化物酶活性变化趋势与漆酶活性变化趋势类似,温度对其影响也基本相同,但部位-Ⅰ、部位-Ⅱ在30℃时的峰值提前到原基时期(时期1),而当温度为25℃和35℃时,锰过氧化物酶活力在菌蕾期(时期3)和开片期(时期4)出现峰值。当培养温度为25℃时,各部位基质中锰过氧化物酶活力略低于另外2个实验组。
2.5 不同时期-部位基质中纤维素酶系酶活变化规律
2.5.1 不同温度对羧甲基纤维素酶活性变化规律的影响
不同温度对羧甲基纤维素酶活性变化规律的影响见图5。

从图5可以看出,随着灵芝的生长发育,灵芝生殖生长阶段基质内羧甲基纤维素酶活性总体呈上升趋势。温度对不同时期和部位基质中羧甲基纤维素酶活力的影响较大,25℃时基质-Ⅰ内的羧甲基纤维素酶活性较低,变化趋势平稳且始终低于30℃和35℃处理组。
2.5.2 不同温度对滤纸酶活性变化规律的影响
温度对灵芝生殖生长阶段不同时期和部位基质中滤纸酶活性变化的影响与对羧甲基纤维素酶活性影响相似,结果见图6。

从图6可以看出,25℃时滤纸酶活性在原基期(时期2)最低,然后有所升高,但总体活性水平仍低于其余2个温度实验组。在30℃和35℃条件下,不同时期和部位基质中灵芝滤纸酶活力波动较大,且活力较高。
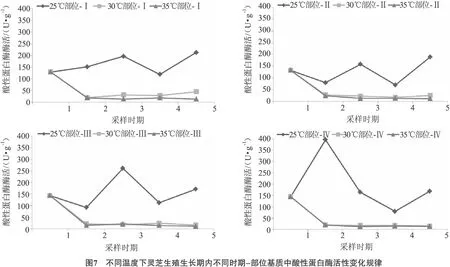
2.6 不同时期-部位基质中酸性蛋白酶活性的变化规律
灵芝生殖生长阶段内,温度对不同时期和部位基质中酸性蛋白酶活性变化规律的影响如图7所示。
从图7可以看出,25℃时,酸性蛋白酶活力总体呈现先升高后下降再升高的趋势,其中部位-Ⅰ、部位-Ⅱ、部位-Ⅲ在菌蕾期(时期3)活性有峰值,而部位-Ⅳ酶活的峰值出现在原基期,各部位在成熟期(时期5)酸性蛋白酶活性有所升高。30℃、35℃时,酸性蛋白酶活性在原基期(时期2)降至最低后基本维持不变。结果显示,出芝温度为25℃时,灵芝酸性蛋白酶活力较高,而其它2个实验组中酸性蛋白酶活力则较低,并且始终低于25℃处理组。
3 讨论
食(药)用菌在菌丝生长阶段,通过分解基质内的木质素和纤维素等,合成、储存子实体发育所需的能量物质[10],进入生殖生长时期后通过分解储存的能量物质供应子实体的发育。本研究发现,出芝温度在25℃时,灵芝可正常出芝;30℃时,仅分化原基;35℃时完全不出芝。推测在25℃时由于灵芝能正常出芝,故需要将基质中储存的能量物质分解以供应其子实体生长,因而其基质的不同时期-部位的还原糖含量均高于30℃和35℃处理组,而总糖含量则低于30℃和35℃处理组。
在本研究中,不同温度条件下,灵芝生殖生长阶段不同时期-部位的基质中木质素酶系的漆酶和锰过氧化物酶整体均呈现小幅下降的趋势,这可能是由于大量木质素在菌丝生长阶段已经被降解,生殖生长阶段灵芝菌丝很少再分泌该类酶,因此温度对其影响不大。由于植物组织中木质素与纤维素以共价键形式紧密结合,并将纤维素分子包埋在其中,木质素降解后再开始降解纤维素,因此纤维素酶系的羧甲基纤维素酶和滤纸酶在生殖生长阶段活性会开始增加[8,11]。本研究也发现,温度对灵芝生殖生长阶段不同时期-部位基质中纤维素酶系的酶活力影响显著,而且随着温度升高其酶活力增加,较高的温度更利于纤维素酶的分泌。
真菌菌丝体中富含蛋白酶,蛋白酶在菌丝体生长过程中参与蛋白质的水解,为菌丝体提供氮源类物质[12]。灵芝转入生殖生长期之后,基质中的蛋白酶活力逐渐升高,用于满足灵芝子实体发育对氮源的需求,因此在灵芝生殖生长阶段,其基质中可溶性蛋白含量下降,而游离氨基酸含量逐渐上升。到了子实体发育后期,由于灵芝对CN需求的变化,灵芝胞外蛋白酶活力迅速下降并维持在较低水平。温度对蛋白酶活力影响较大,当出芝温度25℃时,灵芝基质的不同部位-时期的蛋白酶活力均高于30℃和35℃处理组,提示蛋白酶活力与子实体正常发育密切相关。
综上所述,温度对灵芝子实体形成具有显著的影响,在高于30℃的恒定温度条件下灵芝很难分化子实体,其作用机制还有待进一步深入研究。在实际生产中,灵芝大棚温度需注意保持在25℃左右,避免长期高于30℃,否则会严重影响灵芝的正常生长。
[1]林志彬. 灵芝的现代研究(第三版)[M]. 北京:北京大学医学出版社,2007:1-359.
[2]Adaskaveg JE, Gilbertson RL. Cultural studies of four North American species in theGanodermalucidumcomplex with comparisons toG.lucidumandG.tsugae[J]. Mycological Research, 1989, 92(2): 182-191.
[3]韦会平,刘正宇,谭杨梅,等. 温度条件对金佛山灵芝生长的影响[J]. 中国农学通报,2005,21(10):85-87.
[4]刘雁英,李林辉,谢富刚,等. 食用菌子实体分化发育研究进展[J]. 菌物研究,2010,8(1):57-62.
[5]韦会平,李学刚,蒲盛才. 灵芝生长过程中灵芝多糖含量变化规律研究[J]. 中医药学报,2006,34(5):10-12.
[6]孙伟伟,曹维强,王静. DNS法测定玉米秸秆中总糖[J]. 食品研究与开发,2006,27(6):120-124.
[7]刘建鹏,兰进. 栽培灵芝胞外酶活性及多糖变化规律的初步研究[J]. 中国中药杂志,2009,34(4):483-485.
[8]倪新江,潘迎捷,冯志勇,等. 香菇生长过程中几种胞外酶活性的变化规律[J]. 食用菌学报,1995,2(4):22-27.
[9]王福荣,庞玉珍. 福林—酚试剂法测定蛋白酶活力的条件试验[J]. 调味副食品科技,1981(12):23-26.
[10]Obodai M, Cleland-Okine J, Vowotor KA. Comparative study on the growth and yield ofPleurotusostreatusmushroom on different lignocellulosic by-products[J]. Journal of Industrial Microbiology and Biotechnology, 2003, 30(3): 146-149.
[11]薛海燕. 降解玉米秸秆产纤维素酶和木质素酶菌种的筛选[J]. 酿酒科技,2008(10):44-47.
[12]李艳茹,于海茹,许广波,等. 猪苓菌丝体胞外蛋白酶和漆酶的酶学性质[J]. 江苏农业科学,2011,39(6):423-424.
Effects of Temperatures on the Substance Content and Extracellular Enzyme Activity of Culture Substrate DuringGanodermalucidumReproductive Stage
WEI Wei1, YU Meng-yao1, XU Xiao-yan1, HAN Jia-yu1,3, JIANG Nan1, ZHENG Lin-yong2, LI Wen-tao1,3, LUO Xia1
(1.Sichuan Academy of Chinese Medicine Science, SichuanChengdu610041; 2.Sichuan Academy of Agricultural Sciences, SichuanChengdu610066; 3.College of Life Sciences, Sichuan University, SichuanChengdu610064)
Under 25℃, 30℃ and 35℃ conditions, the changes of substance content (total sugar, reducing sugar, water soluble protein and free amino acid), lignin enzymes (laccase and manganese peroxidase), cellulose enzymes (carboxymethyl cellulase and filter paper enzyme), acid protease activity of bag culture substrate were studied duringGanodermalucidumreproductive stage in different periods of different parts. The results showed thatG.lucidumfruiting body could form at 25℃, and the formation was inhibited above 30℃. DuringG.lucidumreproductive stage, in different periods of different parts, the substrate total sugar and water soluble protein was lower at 25℃ than it at 30℃ and 35℃, and the reducing sugar and free amino acid contents was higher at 25℃ than it at the other two temperatures. The lignin enzymes and cellulase enzymes activity in the culture substrate at 25℃ was lower than it at the other two temperatures, and acid protease activity at 25℃ was higher than it at the other two temperatures. It provided that it was helpful for bag culturedG.lucidumfruiting body formation at about 25℃.
Ganodermalucidum; Bag culture; Temperature; Reproductive stage; Enzymes activities
*项目来源:国家农业产业技术体系药用菌栽培岗位建设项目,项目编号:农科教发[2007]12号;菌类药材研究与开发四川省科技创新团队,项目编号:2011JTD0021;四川省十二五育种攻关项目“菌类药材优质种质资源的收集及新材料的选育”,项目编号:2011NZ0098-12-04;四川省公益性科研院所基本科研项目“袋料灵芝营养代谢-药效成分时空关联模式的研究”,项目编号:A-2014N-6。
魏巍(1982-),男,助理研究员,主要从事食(药)用菌的研究与开发。
**通信作者:罗霞,E-mail:lx1443_cn@sina.com
2014-09-30
S646.9
A
1003-8310(2014)06-0049-07
猜你喜欢
小学生学习指导(低年级)(2022年3期)2022-03-28 08:34:30
小溪流(画刊)(2019年8期)2019-10-09 04:03:15
作文通讯·初中版(2019年2期)2019-04-15 03:26:14
当代陕西(2019年5期)2019-03-21 05:12:30
文苑(2018年22期)2018-11-19 02:54:30
现代园艺(2017年23期)2018-01-18 06:57:57
商周刊(2017年22期)2017-11-09 05:08:34
新农业(2016年18期)2016-08-16 03:28:31
领导科学论坛(2016年4期)2016-02-28 09:18:39
西南军医(2016年6期)2016-01-23 02:21:21
