
不同温度下脉冲式磷酸盐输入对裸甲藻生长的影响
2014-06-26 09:54牛海凤冯剑丰周慧敏
海洋科学 2014年5期
牛海凤,冯剑丰,周慧敏,朱 琳
(南开大学 环境科学与工程学院,环境污染过程与基准教育部重点实验室,天津 300071)
随着城市化进程的加快,临海工业的迅速发展,大量的工业废水和生活污水未经处理直接排入海中,造成近海水质不断恶化,富营养化程度日趋严重,导致赤潮灾害频发,经济损失严重[1-2]。作为一个复杂的海洋生态现象,引起赤潮发生的因素有很多,涉及海洋水文、气象、物理、化学等诸多生态环境因子,但尤其以因营养盐质量浓度增加导致的海水富营养化最为突出[3-6]。目前我国近海大部分海域已呈现出磷酸盐限制的趋势[7-9],而且相对于氮营养盐的多种输入方式[10-11],磷营养盐多来自陆源输入。营养盐脉冲是指营养物质在短时间内急剧升高的现象[12],营养盐脉冲输入作为陆源营养盐向海洋输入的一种重要方式,可以调控营养物质的短期可利用性[13-18],改变海洋浮游生物的种群数量以及群落结构等[19-21]。研究结果也表明,营养盐的不同输入方式(连续输入或者脉冲式输入)能够对浮游生物群落结构产生影响[22-24]。Yamamoto[25]在对广岛湾ohta河口的研究中,模拟脉冲输入和连续输入对 3种海洋赤潮微藻的影响,发现中肋骨条藻对营养盐的脉冲输入变化敏感,链状裸甲藻在营养盐持续添加的模式下生长较好。Sofie等[26]的研究表明,连续的营养盐输入模式对海洋生态系统功能产生有利影响,而高质量浓度营养盐脉冲输入能够极大的影响生物的自然演替,降低生态系统的多样性,而且极可能促进有害赤潮藻的大量爆发。刘亚林等[27]研究了脉冲式营养盐输入对中肋骨条藻生长的影响,结果表明营养盐的脉冲输入对于典型的赤潮藻中肋骨条藻生长有明显的影响,并且营养盐脉冲输入的频率和中肋骨条藻生长波动的频率相同。
除营养盐限制以外,温度也是影响赤潮形成的关键因素[28-30]。营养盐和温度这两个环境因素往往会同时影响着海洋藻类的生长,而营养盐脉冲在不同温度下对海洋藻类生长的影响的研究还较少。作者选取我国近岸海域的典型赤潮藻裸甲藻(Gymnodinium sp.)作为实验藻种,在实验室条件下,模拟陆源营养盐的脉冲输入方式,研究了 3种不同频率的磷酸盐脉冲输入在不同温度下(10℃和 25℃)对海洋浮游植物种群动力学的影响。
1 材料与方法
1.1 藻种及培养
实验所用裸甲藻购自中科院海洋研究所,在温度恒定的光照培养箱(KRG-250BP,上海柏欣仪器设备厂)内培养,培养温度24℃±1℃,光照强度4 000 lx,光暗周期为L︰D=12︰12,培养藻种选用f/2改良培养基[31],人工海水(artificial seawater,AW)的配制参考Artificial Seawater Media[32]。
1.2 实验方法及质量浓度设置
本实验在 10℃和 25℃下分别采用 3种磷酸盐(NaH2PO4)脉冲方式: 即每1天、每4天和每8天投加一次磷酸盐,一个周期(即8天)内所加的磷酸盐总量是相同的。每天投加一次的方式可以认为是近似连续的营养盐输入模式,另外两种可以认为是不同频率的脉冲输入模式,这样就可以更清晰的表明脉冲输入相对于连续输入模式的区别。为了更全面的反映磷酸盐在不同质量浓度水平下的脉冲输入对藻种生长的影响差异,本研究中磷酸盐质量浓度分别设低、高质量浓度组。实验开始前裸甲藻经过无磷饥饿培养,可认为实验开始前培养基内的磷酸盐质量浓度为零。实验过程中,低质量浓度组初始氮质量浓度设为f/2培养基中硝酸盐质量浓度(75 mg/L),高质量浓度组中氮质量浓度扩大4倍(300 mg/L),从而保持氮磷比远大于16︰1,并且其他营养元素供给充足,所以本实验中磷元素为影响裸甲藻生长的主要限制因子。不同温度和质量浓度下均采用此 3种脉冲形式添加营养盐,各组质量浓度设置见表1。

表1 3种脉冲输入模式下磷酸盐的供给量Tab.1 The amount of nutrients supply under three phosphate supply modes
1.3 实验藻种接种及培养
实验前,将处于指数生长期的藻液离心,将沉淀物重新悬浮于无磷培养基中饥饿培养3 d。3 d后,分别取一定体积的藻种以 3 000 r/min的速度离心5 min,弃掉上清液,用无菌人工海水洗涤后离心,重复3次,稀释后接种至实验培养基。初始接种密度为2×104个/mL。
实验在体积为250 mL锥形瓶中添加100 mL培养基中进行,各实验组设置3组平行,每天人工摇动3~4次[33],每日取样并测定藻细胞密度,并且按照不同脉冲方式添加磷酸盐。
1.4 藻细胞计数及磷酸盐测定
采用血球计数板计数的方法,每天取样在光学显微镜下检测细胞密度。接种的当日为第1天,接种密度为2×104个/mL,每天在同一时间计数。取一滴裸甲藻置于血球计数板上,用鲁哥试剂固定后计数,整个取样过程都在无菌操作台上进行。由于取样量极少,相对于培养液总体积可以近似认为符合分批培养试验“试验过程中无底物和菌体的供应和移走”的要求。为了验证实验结果的可靠性,每2天测定一次磷酸盐剩余量,磷酸盐测定采用钼锑抗分光光度法,在一个周期内,总量一定的情况下,磷酸盐吸收量可以用加入磷酸盐总量减去磷酸盐剩余量来计算。
1.5 比生长率

式中,X2为某一时间间隔终结时的藻类现存量;X1为某一时间间隔开始时的藻类现存量;T2−T1为某一时间间隔。
2 结果与分析
2.1 10℃条件下,磷酸盐脉冲对裸甲藻生长的影响
图1a和图2a是10℃低质量浓度磷酸盐脉冲组下裸甲藻的生长曲线和生长率曲线,可以看出在该实验组中裸甲藻均于第13天进入对数生长期。在对数期内,每天输入一次的脉冲方式下藻的长势最好。每天和每 8天输入一次的脉冲方式下,裸甲藻分别在第8天和第10天达到第一个小峰值,分别为10.3×104个/mL和 12.7×104个/mL。在每 4天输入一次的脉冲方式下,裸甲藻藻密度一直处于稳步上升态势,直到实验进行到第21天才出现了较为明显的峰值。每天输入一次的脉冲方式下藻的长势最接近自然状态下藻的的生长趋势,生长曲线比较稳定。每4天和每 8天的脉冲方式下的裸甲藻对数期延长,在实验周期结束时,裸甲藻依然保持较高的增长趋势。从表2(10℃低质量浓度脉冲组磷酸盐吸收量)中可得出,前两个周期内,每天输入的脉冲方式下,磷酸盐吸收量最多,所以其长势也最好(图1a)。
图1b和图2b是10℃高质量浓度磷酸盐脉冲组下裸甲藻的生长曲线和生长率曲线,可以看出,高质量浓度组裸甲藻的藻密度高于低质量浓度各脉冲组。每8天输入一次的脉冲方式、每4天输入一次的脉冲方式和每天输入一次的脉冲方式下的裸甲藻分别在实验进行第11天、第13天和第21天达到第一个峰值,其峰值分别为 13.8×104个/mL,9.8×104个/mL,45×104个/mL。每4天输入一次的脉冲方式下藻的生长曲线有较大波动。每 8天输入一次的脉冲方式下的藻密度在实验周期内一直具有较大优势,从表2(10℃高质量浓度脉冲组磷酸盐吸收量)中也可得出,每8天输入一次的脉冲方式下,磷酸盐吸收量最多。

图1 10℃条件下,裸甲藻在各磷酸盐脉冲组下的生长曲线Fig.1 Growth curves of Gymnodinium sp.under three pulsed phosphate supply at 10℃
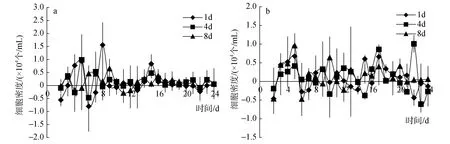
图2 10℃条件下,裸甲藻在各磷酸盐脉冲组下的比生长率曲线Fig.2 Growth rate curves of Gymnodinium sp.under pulsed phosphate supply groups at 10℃
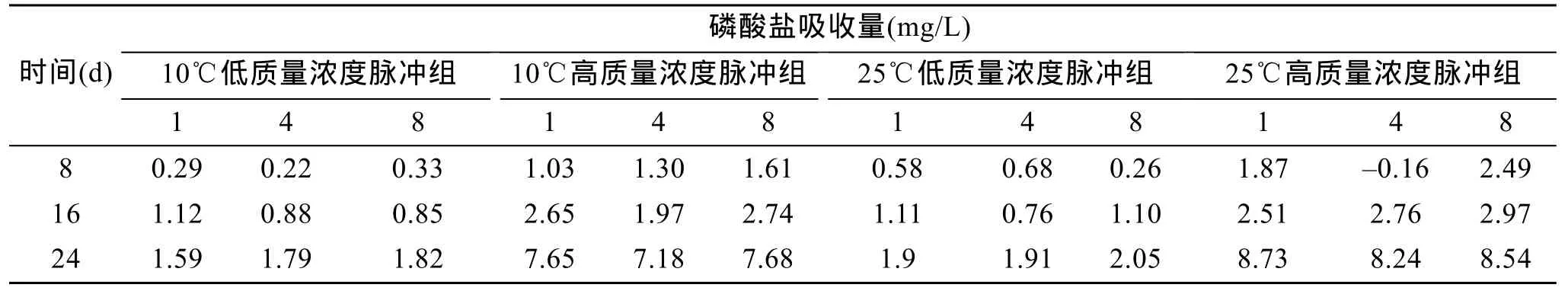
表2 裸甲藻在每个周期内吸收磷酸盐总量Tab.2 The phosphate absorption amount of Gymnodinium sp.in ones cycle
图 3a和 3b分别表示在 10℃条件下,低、高质量浓度磷酸盐脉冲组对裸甲藻每日藻密度平均值的影响。10℃低温条件下,低质量浓度磷酸盐脉冲组中(图 3a),每天输入一次的脉冲方式下裸甲藻的长势在一段时间段内占据优势,但整体来看,差异并不显著(P>0.05);高质量浓度磷酸盐脉冲组中(图 3b),每 8天输入一次的脉冲方式下藻的长势占据明显优势,差异显著(P<0.05)。
2.2 25℃条件下,磷酸盐脉冲对裸甲藻生长的影响
在25℃实验条件下,根据4a和图5a可以看出,25℃低质量浓度组中,裸甲藻在每天输入一次、每4天输入一次和每 8天输入一次的脉冲方式下,先后达到第一个小峰值,分别为15×104个/mL(第9天)、11.17×104个/mL(第 10 天)和 17.83×104个/mL(第 12天)。在整个试验周期中,每天输入一次的脉冲方式下的裸甲藻密度具有较大优势,每4天和每8天输入一次的脉冲方式下的生长曲线波动性较大。随着实验的进行,每天输入的脉冲方式下的裸甲藻进入衰亡期,藻密度不断下降,而每4天和每8天脉冲输入方式下的藻密度不断波动上涨,藻的生长周期延长。从表2(25℃低质量浓度脉冲组磷酸盐吸收量)中得出的实验结果也支持上述观点。
图4b和图5b是25℃高质量浓度磷酸盐脉冲组下裸甲藻的生长曲线和生长率曲线,由图中可得出,裸甲藻在各脉冲输入方式下达到第一个小峰值的的时间和密度大小分别为: 每 8天输入一次(21.83×104个/mL,第9天),每4天输入一次(18.5×104个/mL,第10天),每天输入一次(33.5×104个/mL,第11天)。在整个试验周期中。每天输入的脉冲方式下裸甲藻的藻密度具有明显的优势。从表 2(25℃高质量浓度脉冲组磷酸盐吸收量)中得出,每天输入一次的脉冲方式下,藻的磷酸盐吸收量最大,与藻密度曲线所得出的结论相符。
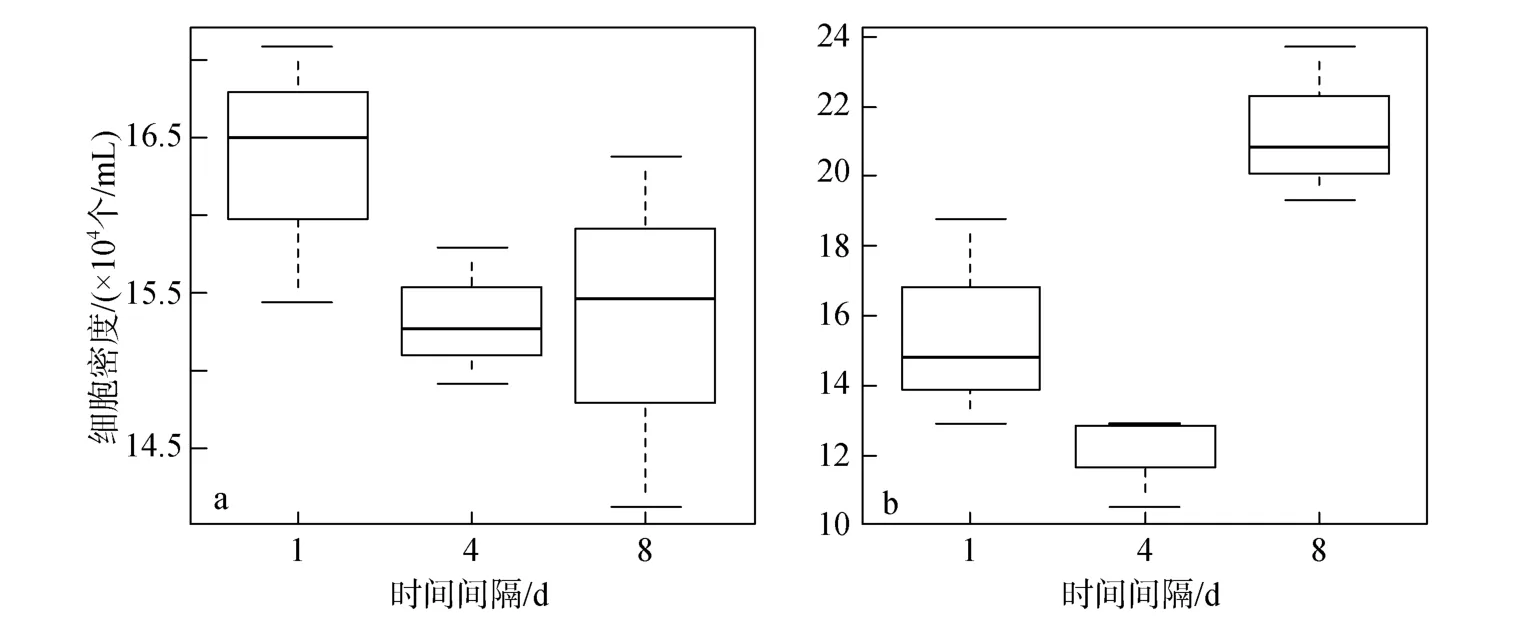
图3 10℃条件下磷酸盐脉冲组对裸甲藻每日藻密度平均值的影响Fig.3 The effect of pulsed phosphate supply on the daily average of Gymnodinium sp.density at 10℃

图4 25℃条件下,裸甲藻在各磷酸盐脉冲组下的生长曲线Fig.4 Growth curves of Gymnodinium sp.under three pulsed phosphate supply at 25℃
图 6a和图 6b分别表示在 25℃条件下,低、高质量浓度磷酸盐脉冲组对裸甲藻每日藻密度平均值的影响。与 10℃低温条件下的结果不同,在 25℃常温条件下(图 6),无论是低质量浓度脉冲组还是高质量浓度脉冲组,基本表现为每天输入一次的脉冲方式下的裸甲藻藻密度都占有较明显优势(低质量浓度组 P<0.01;高质量浓度组 P<0.01)。在 25℃常温条件下,脉冲频率对裸甲藻的的影响基本表现为: 随着脉冲间隔的增加,裸甲藻的平均藻密度呈先降低后升高的趋势,在中等频率的脉冲下,藻密度最低。

图5 25℃条件下,裸甲藻在各磷酸盐脉冲组下的比生长率曲线Fig.5 Growth rate curves of Gymnodinium sp.under pulsed phosphate supply groups at 25℃

图6 25℃条件下磷酸盐脉冲组对裸甲藻每日藻密度平均值的影响Fig.6 The effect of pulsed phosphate supply on the daily average of Gymnodinium sp.density at 25℃
3 讨论与结论
在 10℃实验组中,磷酸盐输入频率的变化会对裸甲藻的生长状况产生影响,并且因磷酸盐质量浓度的不同而不同。低质量浓度磷酸盐脉冲条件下: 每天输入一次的脉冲方式下的裸甲藻先达到一个小峰值,但其峰值小于另外两种脉冲方式;进入对数期后,每天输入的脉冲方式下的裸甲藻在一段时间内具有较大优势,并且该方式下裸甲藻的长势接近自然状态下藻的生长周期,有比较明显的调整期、对数期、稳定期和衰亡期;随着实验时间延长,每4天和每 8天脉冲输入模式下的藻细胞数量才逐渐超过每天输入一次的脉冲方式。有关研究也表明,低温胁迫下,初始阶段藻的适应能力较差,光合作用弱,随着实验时间延长,才逐渐启动恢复机制[34]。每4天和每8天的脉冲输入方式下的裸甲藻对数期延长,在实验周期结束时,裸甲藻依然保持较高的增长趋势,生长周期延长,相关的研究成果也表明脉冲间隔的增加在一定条件下会延长藻的生长周期[35]。在10℃实验组的高质量浓度磷酸盐脉冲条件下: 每 8天输入一次的脉冲方式下藻的长势在整个试验周期中具有明显优势,并且在试验周期内一直保持上升态势,这与低质量浓度组所得的结果有所差异。这可能是因为在低温胁迫以及高质量浓度磷酸盐水平下,藻的其他生理指标发生变化,而没有在藻密度方面体现出来,这一点有待于作者做进一步的研究。该脉冲组中,每天输入一次的脉冲方式下藻的生长曲线较为平稳。
通过比较发现,在 10℃实验条件下,无论是低质量浓度脉冲组还是高质量浓度脉冲组,每天输入的脉冲方式下的裸甲藻生长曲线都较为稳定,这可能是因为脉冲输入模式相比连续输入模式对藻类生长的干扰程度强,而每天输入的方式接近连续输入模式,使得另外两种脉冲输入方式对裸甲藻生长的干扰更大,刘亚林等[27]的研究也表明,每天输入的脉冲模式下,藻的生长曲线最为平稳。
在 25℃实验组中,无论是低质量浓度脉冲组还是高质量浓度脉冲组,每天输入一次的脉冲方式下的裸甲藻藻密度都具有比较大的优势,这可能是因为每天输入模式给藻的生长创造了一个较为稳定的环境,使其较快适应环境,有利于藻的生长。每4天和每 8天输入的脉冲方式下的藻类生长曲线波动较大,每天输入的脉冲方式下的生长曲线相对平稳,这也与 10℃实验组所得的结论一致。根据磷酸盐脉冲组对裸甲藻每日藻密度平均值的影响结果可得出,较高温度条件下,长周期脉冲输入方式可能会导致裸甲藻的平均密度下降,而在较低温度及较高的磷酸盐质量浓度水平下,长周期脉冲输入方式可能会导致藻细胞数量的显著增加,从而可能引发甲藻赤潮的形成。
在所有实验组中,脉冲频率对裸甲藻的的影响基本表现为: 随着脉冲间隔的增加,裸甲藻的平均藻密度呈先降低后升高的趋势,在中等频率的脉冲下,藻密度最低,出现这种结果可能有以下两方面的原因: 一方面是因为设定的脉冲频率较少,还不能够完全反应出藻种在不同脉冲频率下的生长状况;另一方面,可能是因为藻的其他生理指标发生了变化,但是没有在藻密度方面体现出来。这一点也需要作者做更深入的研究,以进一步探索脉冲式营养盐输入对海洋浮游植物的影响。
本文的研究结果表明,近似连续的输入方式(每天输入一次的脉冲方式)下藻的生长状况比脉冲式的输入方式(每4天和每8天输入一次的脉冲方式)稳定,后两者生长曲线波动较为明显,这说明脉冲输入模式对藻类生长的干扰程度强于连续输入模式;在较高温度条件下,长周期脉冲输入方式可能会导致赤潮藻的平均密度下降,而在低温及较高的磷酸盐质量浓度水平下,长周期脉冲输入方式可能会导致藻细胞数量的显著增加,从而可能引发甲藻赤潮的形成。
[1]赵冬至,赵玲,张丰收.我国海域赤潮灾害的类型、分布与变化趋势[J].海洋环境科学,2003,3: 7-11.
[2]王初升,唐森铭,宋普庆.我国赤潮灾害的经济损失评估[J].海洋环境科学,2011,3: 428-431.
[3]黄小平,黄良民,谭烨辉,等.近海赤潮发生与环境条件之间的关系[J].海洋环境科学,2002,4: 63-69.
[4]徐宁,吕颂辉,段舜山,等.营养物质输入对赤潮发生的影响[J].海洋环境科学,2004,5: 20-24.
[5]姚炜民,郑爱榕,邱进坤.浙江洞头列岛海域水体富营养化及其与赤潮的关系[J].海洋环境科学,2007,5:466-469.
[6]何本茂,韦蔓新.北海湾赤潮形成原因及机理[J].海洋环境科学,2009,1: 62-66.
[7]孙丕喜,王宗灵,战闰,夏滨,王湘芹.胶州湾海水中无机氮的分布与富营养化研究[J].海洋科学进展,2005,23(4): 466-471.
[8]霍文毅,俞志明,邹景忠,等.胶州湾中肋骨条藻赤潮与环境因子的关系[J].海洋与湖沼,2001,32(3):311-318.
[9]王保栋.黄海和东海营养盐分布及其对浮游植物的限制[J].应用生态学报,2003,14(7): 1122-1126.
[10]徐开钦,林诚二,牧秀明,等.长江干流主要营养盐含量的变化特征——1998~1999年中日合作调查结果分析[J].地理学报,2004,59(1): 118-124.
[11]王佳宁,晏维金,贾晓栋.长江流域点源氮磷营养盐的排放、模型及预测[J].环境科学学报,2006,26(4):658-666.
[12]Ostfeld R S,Keesing F.Pulsed resources and community dynamics of consumers in terrestrial ecosystems[J].Trends in Ecology and Evolution,2000,15: 232-237.
[13]Taslakian M J,Hardy J T.Sewage nutrient enrichment and phytoplankton ecology along the central coast of Lebanon[J].Marine Biology,1976,38: 315-325.
[14]GarciaSoto C,Demadariaga I,Villate F,et al.Day-to-day variability in the plankton community of a coastal shallow embayment in response to changes in river runoff and water turbulence[J].Estuarine,Coastal and Shelf Science 1990,31(3): 217 -229.
[15]Padisak J.The influence of different disturbance frequencies on the species richness,diversity and equitability of phytoplankton in shallow lake[J].Hydrobiologia,1993,249: 135-156.
[16]Justic D,Rabalais N N,Turner R E,et al.Changes in nutrient structure of river-dominated coastal waters:stoichiometric nutrient balance and its consequences[J].Estuarine,Coastal and Shelf Science,1995,40(3):339-356.
[17]Piehler M F,Twomey L J,Hall N S,et al.Impacts of inorganic nutrient enrichment on phytoplankton community structure and function in Pamlico Sound,NC,USA[J].Estuarine,Coastal and Shelf Science,2004,61(2): 197-209.
[18]Buyukates Y,Roelke D.Influence of pulsed inflows and nutrient loading on zooplankton and phytoplankton community structure and biomass in microcosm experiments using estuarine assemblages[J].Hydrobiologia,2005,548: 233-249.
[19]顾新根.“南水北调”对浮游植物的影响[J].海洋渔业,1993,1: 8-12.
[20]吴玉霖,付月娜,张永山,等.长江口海域浮游植物分布及其与径流的关系[J].海洋与湖沼,2004,35(3):246-251.
[21]Ornolfsdottir E B,Lumsden S E,Pinckney J L.Nutrient pulsing as a regulat or of phytoplankton abundance and community composition in Galveston Bay,Texas[J].J Exp Mar Biol Ecol,2004,303(2): 197-220.
[22]Gaedeke A,Sommer U.The influence of the frequency of periodic disturbances on the maintenance of phytoplankton diversity[J].Oecologia,1986,71: 25-28.
[23]Capblancq J.Nutrient dynamics and pelagic food web interactions in oligotrophic and eutrophic environments:an overview[J].Hydrobiologia,1990,207: 1-14.
[24]Buyukates Y,Roelke D.Influence of pulsed inflows and nutrient loading on zooplankton and phytoplankton community structure and biomass in microcosm experiments using estuarine assemblages[J].Hydrobiologia,2005,548: 233-249.
[25]Yamamoto T,Hatta G.Pulse nutrient supply as a factor inducing phytoplankton diversity[J].Ecological Modelling,2004,171(3): 247–270.
[26]Sofie S,George T,Daniel B D,et al.Effects of pulsed nutrient inputs on phytoplankton assemblage structure and blooms in an enclosed coastal area[J].Estuarine,Coastal and Shelf Science,2007,73: 807-815.
[27]刘亚林,韩笑天,白洁,等.脉冲式营养盐输入对中肋骨条藻生长的影响[J].海洋科学,2008,2: 51-55.
[28]陈艳拢,赵冬至,杨建洪,等.赤潮藻类温度生态幅的定量表达模型研究[J].海洋学报,2009,31(5):156-161.
[29]缪宇平,袁琪,周宏农,等.环境因子对微小亚历山大藻 Amtk-9生长与产毒的综合影响[J].海洋渔业,2009,31(3): 279-281.
[30]邵盛男,缪宇平,周宏农,等.环境因子对链状亚历山大藻生长的影响[J].海洋渔业,2011,33(1): 66-73.
[31]Guillard R R L,Ryther J H.Studies of marine planktonic diatoms.I Cyclotella nana Hustedt,and Detonula confervacea(Cleve)Gran[J].Microbiol,1962,8(2): 229-239.
[32]Guillard R R L.Culture of phytoplankton for feeding marine invertebrates[C]//Culture of Marine Invertebrate Animals.Plenum Press: New York USA,1975: 26-60.
[33]Bitaubé P E,Caro P I,Pérez R L.Kinetic model for growth of Phaeodactylum tricornutum in intensive culture photobioreactor[J].Biochemical Engineering Journal,2008,40(3): 520-525.
[34]梁英,陈书秀,田传远,等.温度对三角褐指藻叶绿素荧光特性及生长的影响[J].中国海洋大学学报,2008,38(3): 377-383.
[35]樊娟.氮、磷浓度及氮输入频度对两种海洋微藻生长的影响研究[D].天津: 南开大学,2010.
猜你喜欢
海洋学报(2021年2期)2021-03-15
理论与创新(2020年16期)2020-11-03
学与玩(2019年8期)2019-10-29
资源节约与环保(2019年10期)2019-01-21
妇女生活(2017年5期)2017-05-16
幼儿教育·教育科学版(2016年5期)2016-09-29
浙江大学学报(工学版)(2015年1期)2015-03-01
中国酿造(2014年9期)2014-03-11
棋艺(2001年9期)2001-07-17
棋艺(2001年11期)2001-05-21
