
PAX6基因多态性与精神分裂症的关系探讨
2014-06-14 09:48韩海斌杨建立温子玉
山东医药 2014年12期
韩海斌,杨建立,丁 玉,温子玉
(1天津医科大学,天津300070;2天津市精神卫生中心)
精神分裂症是一种常见的病因尚未完全阐明的精神病,其发生可能与神经发育异常有关[1~3]。目前认为,遗传因素和某些神经发育危险因素的相互作用,使胚胎期大脑发育过程出现某种神经病理变化,是精神分裂症神经发育异常的机制之一。PAX6作为PAX基因家族中的一员,定位于人染色体第11号染色体长臂13位点(11p13)。大脑皮层发育期间PAX6缺陷导致成熟期认知缺陷,发育阶段PAX6缺失导致皮层、皮层间及皮层内连接严重缺损,这可能成为鼠及人类出生后认知缺损的基础[4]。相关研究表明,PAX6基因多态性可能通过调节发育中或成年的大脑而参与偏执型精神分裂症的发生[5]。本研究采用病例对照研究方法,探讨PAX6基因多态性与精神分裂症的关系。
1 资料与方法
1.1 临床资料 患者组:2011~2013年在天津市精神卫生中心治疗的首发精神分裂症患者270例,均为汉族,年龄18~45岁。诊断由2名以上精神科主治医师独立完成,符合DSM-IV关于精神分裂症的诊断标准。排除烟酒及其他精神活性物质滥用者、有其他精神障碍家族史者、有肝肾严重躯体疾病者。患者入组前详细了解本研究目的和程序,并签署知情同意书。对照组:健康志愿者320例,均为汉族,年龄、性别与患者组具有可比性;一般健康问卷各项目总分<30分;经非结构式临床访谈排除精神疾病,且家族中无精神疾病和自杀者,个体间无血缘关系;在详细了解本研究目的和程序后,均签署知情同意书。
1.2 方法
1.2.1 标本采集与处理 以EDTA作为抗凝剂,取肘静脉血10 mL充分抗凝处理,冷藏保存,离心后-80℃冷冻。
1.2.2 基因多态性检测 单核苷酸多态性(SNP)位点:通过HapMap数据库检索PAX6基因34 kb区域(覆盖上游8 kb和下游4 kb),选取其中位于PAX6 的3'非翻译区(3'-UTR)的 rs3026401、rs1506两个标签SNP位点。采用连接酶检测反应研究SNP的多态性。
采用AXYGEN公司的AxyPrep-96全血基因组DNA试剂盒提取人类基因组DNA。检测总DNA浓度、纯度、完整性,扩增 SNP位点所在片段,准备PCR Master Mix(20 μL体系)。反应条件:95℃变性15 min,94 ℃ 30 s、56 ℃ 1 min、72 ℃ 1 min(35个循环),72℃延伸7 min。反应结束后,取2 μL反应产物在3.0%琼脂糖凝胶、0.5×TBE中电泳。剩余样品保存于-20℃。在PCR扩增产物中加入等体积ddH2O稀释,作为连接反应的模板。连接反应体系(10 μL):在 1.5 mL eppendorf离心管中分别加入 100 μL buffer(10 × )、100 μL Probe Mix、5 μL 连接酶、695 μL去离子水。充分混匀后离心,再取9 μL分装在200 μL的 PCR反应管中,最后再加入1 μL PCR反应产物。在Perkin-Elmer Gene Amp PCR Systems9600上设置如下程序:95℃变性2 min,94℃ 30 s、50 ℃ 2 min(35 个循环)。取1 μL LDR 连接产物、1 μL ABI GS-500 ROX荧光标记分子量标准、1 μL去离子甲酰胺上样液混合,95℃加热变性2 min,冰中骤冷;于5%聚丙烯酰胺和5 mol/L尿素中3 000 V电泳2.5 h,应用Genescantm672软件进行数据收集、泳道线校正、迁移片段大小测量和校正内在分子量标准,应用Genemapper软件进行数据分析和基因分型。
1.2.3 临床症状评分比较 治疗前采用阳性与阴性症状量表(PANSS)评定阳性症状、阴性症状及一般精神病理因子情况,对患者组SNP rs3026401不同基因型患者的临床症状评分进行比较。
1.2.4 统计学方法 采用SPSS13.0统计软件。组间基因型和等位基因分布频率比较采用χ2检验,rs3026401各基因型PASS评分比较采用方差分析;使用 SHEsis遗传分析软件[6]进行 Hardy-Weinberg平衡、连锁不平衡(LD)检验及单位点和单体型遗传关联分析。P≤0.05为差异有统计学意义。
2 结果
2.1 样本情况 因为检测过程中样本损失,患者组8例、对照组2例的rs3026401和(或)rs1506位点数据缺失,故在统计中将其剔除。最终患者组262例、对照组318例纳入本研究。
2.2 基因型及等位基因分析结果
2.2.1 Hardy-Weinberg 平衡检验结果 两组的SNP rs3026401基因分布均符合Hardy-Weinberg平衡定律,χ2=0.42、0.37,P 均 >0.05;SNP rs1506 基因分布也均符合 Hardy-Weinberg平衡定律,χ2=2.20、0.08,P 均 >0.05;表明研究样本具有群体代表性。
2.2.2 Genemapper数据分析结果 患者组各位点的分型截图见图1。
2.2.3 两组 PAX6 SNP rs1506、rs3026401 基因型、等位基因分布情况 患者组SNP rs1506基因型、等位基因分布与对照组比较,差异均无统计学意义(χ2=2.877、1.187,P 均 >0.05);患者组 SNP rs3026401 基因型、等位基因分布与对照组比较,差异均有统计学意义(χ2=11.635、11.044,P 均 <0.05)。见表1、2。
2.2.4 SNP rs1506、rs3026401的LD 分析结果 分析结果显示,D'=1、r2=0.812,表明这两个位点之间存在很强的LD。对这两个位点构成的单倍型进行分析,结果显示患者组和对照组之间单倍型rs3026401A/rs1506T、rs3026401G/rs1506T 频率差异具有统计学意义(P均<0.05)。

图1 Genemapper数据分析患者组各位点的分型截图

表1 两组SNP rs1506基因型、等位基因分布[例(%)]
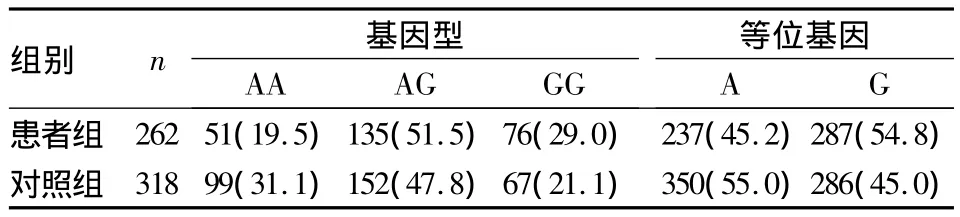
表2 两组SNP rs3026401基因型、等位基因分布[例(%)]
2.2.5 患者组SNP rs3026401不同基因型患者的临床症状评分比较 患者组SNP rs3026401不同基因型的阳性症状评分、一般精神病理因子评分差异均无统计学意义(P均>0.05),而阴性症状评分差异具有统计学意义(P<0.05),即 GG>AA、GG>AG(P均 <0.05)。见表3。
表3 患者组SNP rs3026401不同基因型患者的PANSS 评分比较(±s)

表3 患者组SNP rs3026401不同基因型患者的PANSS 评分比较(±s)
SNP rs3026401基因型PANSS评分(分)阳性症状 阴性症状 一般精神病理因子AA 21.25±3.71 22.88±3.91 40.13±5.46 AG 21.33±4.29 23.38±4.60 41.38±5.34 GG 20.58±3.67 25.03±4.21 41.02±4.24 F 0.907 4.738 1.126 P 0.405 0.010 0.326
3 讨论
精神分裂症多起病于青壮年,常缓慢起病,病程迁延,有慢性化倾向和衰退的可能。国内外相继进行的大规模流行病学调查显示,精神分裂症患病率为1.4‰ ~4.6‰,发病率为0.16‰ ~0.43‰[7]。精神分裂症的病因至今尚未明确,现有治疗方法效果均不理想[8]。
人与鼠的PAX6基因具有高度同源性。动物实验表明,PAX6基因在发育过程中的脑和成体脑中都有表达,且对脑发育和脑结构有重要作用[9,10]。此外,有研究表明PAX6参与出生后海马神经发生的祖细胞的形成和维持[11],而海马的发生又与精神分裂症关系密切[12]。动物学研究还发现,PAX6抑制鼠类星形胶质细胞增殖,促进其成熟[13];PAX6杂合鼠(PAX6-/+)表现出没有松果体及单边多小脑回等[14]。有研究表明,PAX6基因突变能够引起人脑结构异常[15]。精神分裂症的神经发育异常假说认为,由于遗传因素和某些神经发育危险因素的相互作用,在胚胎期大脑发育过程就出现了某种神经病理变化。研究发现,精神分裂症患者海马、额皮层、扣带回和内嗅皮层有细胞结构紊乱,在这些变化中不伴有胶质细胞增生,可能是由脑发育阶段神经元移行异位或分化障碍造成的,破坏了皮层联络的正常模式[16]。近1个世纪以来,大量的关于精神分裂症的家系调查[17]、双生子研究[18]、寄养子研究[19]、遗传流行病学研究等结果强烈提示,精神分裂症与遗传因素密切相关。现有研究显示,精神分裂症并不遵循孟德尔遗传规律,它更可能是一类由多个、有较小影响和外显率、相互作用的潜在基因共同作用的结果。由此可见,寻找并定位与精神分裂症发病有较大关联的基因位点非常重要。很多研究者致力于寻找与精神分裂症相关联的基因多态性的研究,诸如代谢型谷氨酸受体-3基因[20]、精神分裂症断裂基因1(DISC1)[21]等的研究。
前脉冲抑制模型(PPI)是研究精神分裂症的一个重要模型。有关研究显示,PAX6杂合鼠的PPI明显受损,同时伴有出生后的神经发育异常[22]。大脑皮层发育期间PAX6缺陷导致成熟期认知缺陷,发育阶段PAX6缺失导致皮层、皮层间及皮层内连接严重缺损,这成为鼠及人类出生后认知缺损的基础[4]。作为早期表达的PAX6基因多态性,可能通过调节发育中或成年的大脑而参与偏执型精神分裂症的发生[5]。同时也有报道称,PAX6促进精神分裂症可能是通过破坏维甲酸的信号途径[23]。mRNA的3'-UTR作为mRNA的一部分,具有miRNA、siRNA的靶位点,miRNA或siRNA结合靶位点后可引起mRNA的降解或抑制翻译起始,其调控自身mRNA的功能已为人们所熟知。本研究通过检索HapMap数据库在PAX6基因3'-UTR区选取其中两个标签 SNP rs3026401、rs1506 位点[24],既往也有这两个位点在其他神经系统疾病中的相关研究[25]。本研究发现,患者组SNP rs1506基因型、等位基因频率分布频率与对照组相比差异无统计学意义,而SNP rs3026401基因型、等位基因分布频率与精神分裂症均相关,提示PAX6基因rs3026401多态性可能是精神分裂症的危险因素之一。我们进一步比较了SNP rs3026401不同基因型的PANSS评分情况,结果显示阳性症状评分、一般病理因子评分差异无统计学意义,而阴性症状评分存在统计学差异,其中GG型阴性症状评分显著高于AA、AG型,这可能是PAX6基因rs3026401多态性引起精神分裂症易感的重要原因;但目前国内外相关文献对此方面的报道较少,有待进一步研究以验证。
本研究还存在不足之处,应在标准化诊断分类的基础上进一步扩大样本量,通过LD及普通关联分析的方法在不同人群中重复研究,以进一步明确该位点与精神分裂症易感性的关系。同时,还应对PAX6基因rs3026401位点在精神分裂症中的具体功能进行研究,以进一步发展与疾病相关的生物学内表型的分析,这将更有利于深入探讨精神分裂症发生的基础,为更好的预防和治疗精神分裂症提供理论支持。
[1]Prasad KM,Keshavan MS.Structural cerebral variations as useful endophenotypes in schizophrenia:do they help construct"extended endophenotypes"[J].Schizophr Bull,2008,34(4):774-790.
[2]Shenton ME,Dickey CC,Frumin M,et al.A review of MRI findings in schizophrenia[J].Schizophr Res,2001,49(1-2):1-52.
[3]Kircher TT,Thienel R.Functional brain imaging of symptoms and cognition in schizophrenia[J].Prog Brain Res,2005(150):299-308.
[4]Tuoc TC,Radyushkin K,Tonchev AB,et al.Selective cortical layering abnormalities and behavioral deficits in cortex-specific Pax6 knock-out mice[J].J Neurosci,2009,29(26):8335-8349.
[5]Stober G,Syagailo YV,Okladnova O,et al.Functional PAX-6 gene-linked polymorphic region:potential association with paranoid schizophrenia[J].Biol Psychiatry,1999,45(12):1585-91.
[6]Shi YY,He L.SHEsis,a powerful softw are platform for analyses of linkage disequilibrium,haplotype construc tion,and genetic assoc iation at polymorphismloci[J].Cell Res,2005,15(2):97-981.
[7]Jablensky A.Epidemiology of schizophrenia:the global burden of disease and disability[J].Eur Arch Psychiatry Clin Neurosci,2000,250(6):274-285.
[8]李霞,潘丽,金国东,等.帕利哌酮缓释胶囊治疗青少年精神分裂症的疗效及安全性[J].山东医药,2011,15(51):96-97.
[9]Stoykova A,Fritsch R,Walther C,et al.Forebrain patterning defects in Small eye mutant mice[J].Development,1996,122(11):3453-3465.
[10]Engelkamp D,Rashbass P,Seawright A,et al.Role of Pax6 in development of the cerebellar system[J].Development,1999,126(16):3585-3596.
[11]Maekawa M,Takashima N,Arai Y,et al.Pax6 is required for production and maintenance of progenitor cells in postnatal hippocampal neurogenesis[J].Genes Cells,2005,10(10):1001-1014.
[12]Jun H,Mohammed Qasim Hussaini S,Rigby MJ,et al.Functional role of adult hippocampal neurogenesis as a therapeutic strategy for mental disorders[J].Neural Plasticity,2012(2012):1-20.
[13]Sakurai K,Osumi N.The neurogenesis-controlling factor,Pax6,inhibits proliferation and promotes maturation in murine astrocytes[J].J Neurosci,2008,28(18):4604-4612.
[14]Mitchell TN,Free SL,Williamson KA,et al.Polymicrogyria and absence of pineal gland due to PAX6 mutation[J].Ann Neurol,2003,53(5):658-663.
[15]Free SL,Mitchell TN,Williamson KA,et al.Quantitative MR image analysis in subjects with defects in the PAX6 gene[J].Neuroimage,2003,20(4):2281-2290.
[16]Stefan MD,Murray RM.Schizophrenia:developmental disturbance of brain and mind[J].Acta Paediatr Suppl,1997,86(422):112-116.
[17]Bladessarini RJ.Schizophrenia[J].N Engl J Med,1977,297(18):988-995.
[18]Saha S,Chant D,Welham J,et al.A systematic review of the prevalence of schizophrenia[J].PloS Medicine,2005,2(5):413-433.
[19]Macdonald AW,Schulz SC.What we know:findings that every theory of schizophrenia should explain[J].Schizsophrenia Bulletin,2009,35(3):493-508.
[20]潘成英,岳伟华,卢天兰.代谢型谷氨酸受体-3基因多态性和精神分裂症临床症状关联研究[J].中国心理卫生杂志,2009,23(7):528-532.
[21]孙伟,闫俊,王力芳,等.精神分裂症断裂基因1(DISC1)多态性与精神分裂症的关联研究[J].中国心理卫生杂志,2009,23(8):590-594.
[22]Maekawa M,Takashima N,Matsumata M,et al.Arachidonic acid drives postnatal neurogenesis and elicits a beneficial effect on prepulse inhibition,a biological trait of psychiatric illnesses[J].PLoS One,2009,4(3):1-9.
[23]Lamantia AS.Forebrain induction,retinoic acid,and vulnerability to schizophrenia:insights from molecular and genetic analysis in developing mice[J].Biol Psychiatry,1999,46(1):19-30.
[24]Hammond CJ,Andrew T,Mak YT,et al.A susceptibility locus for myopia in the normal population is linked to the PAX6 gene region on chromosome 11:a genomewide scan of dizygotic twins[J].Am J Hum Genet,2004,75(2):294-304.
[25]Jiang B,Yap MK,Leung KH,et al.PAX6 haplotypes are associated with high myopia in Han chinese[J].PLoS One,2011,6(5):1-10.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
智慧健康(2021年17期)2021-07-30
国际检验医学杂志(2021年7期)2021-04-15
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
现代检验医学杂志(2016年5期)2016-08-20
西南农业学报(2016年6期)2016-04-16
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
现代检验医学杂志(2015年6期)2015-02-06
作物研究(2014年6期)2014-03-01
河南医学研究(2014年7期)2014-02-27
