
2012年楚科奇海及其邻近海域浮游植物现存量和初级生产力粒级结构研究
2014-06-01 12:30乐凤凤郝锵金海燕李涛庄燕培翟红昌刘诚刚陈建芳
海洋学报 2014年10期
乐凤凤,郝锵,3,金海燕,3,李涛,庄燕培,翟红昌,刘诚刚,3,陈建芳,3
(1.国家海洋局第二海洋研究所浙江杭州 310012;2.国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州 310012;3.卫星海洋环境动力学国家重点实验室,浙江杭州 310012;4.中国海洋大学,山东青岛 266003)
2012年楚科奇海及其邻近海域浮游植物现存量和初级生产力粒级结构研究
乐凤凤1,2,郝锵1,2,3,金海燕1,2,3,李涛4,庄燕培1,2,翟红昌1,2,刘诚刚1,2,3,陈建芳1,2,3
(1.国家海洋局第二海洋研究所浙江杭州 310012;2.国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州 310012;3.卫星海洋环境动力学国家重点实验室,浙江杭州 310012;4.中国海洋大学,山东青岛 266003)
通过2012年夏季第五次北极科学考察期间在楚科奇海及其邻近海域现场调查所获得的数据分析研究了海域的粒度分级叶绿素a浓度和初级生产力。结果表明,叶绿素a浓度和初级生产力的高值均出现在楚科奇海陆架区,并且远高于深海区。去程时调查海域水层平均叶绿素a浓度的变化范围为0.32~15.66 mg/m3,平均(2.77±3.96)mg/m3,高值区出现在南部邻近白令海峡海域、北部阿拉斯加巴罗近岸和冰缘区;初级生产力的范围为50.11~943.28 mg/(m2·d),高值出现在冰缘水华区。返程时水层平均叶绿素a浓度的变化范围为0.07~1.52 mg/m3,平均(0.41±0.40)mg/m3,高值仍出现在陆架区,但比去程时低了一个数量级;初级生产力的分布范围为12.31~41.35 mg/(m2·d),高值出现在陆架区。浮游植物粒度分级测定结果表明,在生物量较低的深海区,叶绿素a浓度和初级生产力的粒级结构以微微型浮游生物(Pico级份)占优势(其贡献率分别为46.1%和56.9%),小型(Net级份)和微型(Nano级份)对总叶绿素a浓度的贡献差异极小,分别为26.6%和27.3%,对总初级生产力的贡献分别为23.8%和19.3%;而在生物量较高的水深小于200 m的陆架区,Net级份叶绿素a浓度所占百分比最高,Pico级份次之,Nano级份最低,分别为59.8%、27.9%和12.3%,初级生产力的粒级结构中叶绿素a浓度所占百分比由高到低同样是Net、Pico和Nano,所占百分比分别为60.6%,32.2%和7.2%。
楚科奇海;北极;浮游植物现存量;初级生产力;粒级结构
1 引言
北冰洋是全球纬度最高的大洋,也是对全球变化最敏感的地区之一。有研究表明,近几十年来北冰洋发生了前所未有的变化,在全球增暖的大背景下,北极地区变暖趋势非常明显[1],其中最显著的表现就是北极海冰的快速缩减[2—4],而海冰面积的减少、冰层厚度的降低则会影响与海冰关系密切的海水生态系统,并打破海冰生态系统和海水生态系统初级生产力之间的平衡[5]。因此,北冰洋生态系统对海冰快速变化的响应已经成为海洋科学家关注的热点问题,也是我国历次北极考察的科学目标之一。
楚科奇海位于北冰洋西部,是北冰洋中最大的边缘海(大部分海域水深浅于50 m),也是北太平洋与北冰洋相互连接的通道,其南部通过白令海与太平洋相连,北面以76°N的北冰洋陆坡为界,西部通过得朗海峡与东西伯利亚海连通,在东北部巴罗角附近与波弗特海相接。该海域生态系统中浮游植物生物量和生产力水平受北冰洋环流、海冰动力过程和北太平洋入流水的性质影响。浮游植物生物量和初级生产力是我国历次北极考察的重要内容[6—7],但由于北极地区气候、位置等方面的独特性造成了采样难度大的状况,有关楚科奇海浮游植物生物量和生产力的实测数据积累较少。本研究基于第五次北极考察期间在楚科奇海域实测获得的叶绿素、初级生产力及其粒级结构数据,结合物理、化学资料,对其分布特征进行研究,并分析讨论不同水团对浮游植物现存量和初级生产力的影响,为楚科奇海域物理-化学-生物耦合过程研究提供基础资料和科学依据。
2 材料与方法
2.1 调查海区和采样站位
2012年7月18日—9月8日搭乘“雪龙”号极地科学考察船在楚科奇海及其邻近海域进行了叶绿素a浓度和初级生产力现场调查,站位布设范围在66.7°~82°N,162°~173°W海域内。7月18—20日(去程)完成4个断面(R断面设5站,CC断面设7站,C区设6站)共18个站位的观测,9月4—8日(返程)完成2个断面(SR断面设13站,M断面设7站)共20个站位的观测,其中SR断面是R断面的复测断面,站位SR01、03、05、07、09为R01、02、03、04、05的复测站位。CC断面和C区观测断面自近岸向外延伸,R断面和SR断面为沿169°W经线呈南北走向的观测断面,M断面为自门捷列夫深海平原向陆架一侧的南北向断面(图1)。
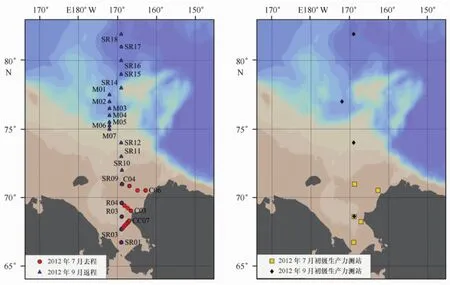
图1 2012年楚科奇海采样观测站位分布Fig.1 Sampling stations in the Chukchi Sea during the summer of 2012
2.2 样品的采集和观测方法
水样的采集使用ROSETTE采水器,叶绿素a样品的采集按标准层或根据海区真光层深度、跃层分布特点确定采样深度,按萃取荧光法[8]进行测定。海水透明度使用Secchi盘测定,根据透明度求得各不同光透射率层次深度[9],按表层和海面入射光强衰减至50%、30%、10%、3%和1%的深度采集现场初级生产力测定用水样。初级生产力的测定采用改进的同位素(14C)示踪法[10—11]进行。
所有水样收集前,经200μm孔宽的筛绢预过滤,以除去大多数的浮游动物。叶绿素a和初级生产力粒度分级测定用水样经孔宽20μm的筛绢、孔径2.0 μm的核微孔滤膜和Whatman GF/F玻璃纤维滤膜过滤,以分别获取小型(Net,亦称Micro级份,大于20 μm),微型(Nano级份,大于2μm,小于20μm)和微微型(Pico级份,小于2μm)的光合浮游生物。
3 结果
由于去程(7月18—20日)和返程(9月4—8日)调查时间间隔较大,调查海域环境条件变化巨大,为了更好地阐述浮游植物生物量和生产力的分布特征及其与环境因素之间的关系,将所获观测结果按去程和返程分开进行描述。
3.1 浮游植物现存量
3.1.1 叶绿素a浓度及其分布
去程时,所有站位水深较浅,平均水深只有47 m,各站位水层平均叶绿素a浓度的平面分布见图2。从图2可见,在R断面北部冰缘区、邻近白令海峡的楚科奇海南部和阿拉斯加巴罗近岸出现叶绿素a浓度高值区,显著高于中部的叶绿素a浓度(小于1.00 mg/m3)。R断面最北端R05站叶绿素a浓度高达15.66 mg/m3,发生浮游植物水华。图3展示的是按做站顺序绘制的叶绿素a浓度垂直剖面图。由图可见,叶绿素a浓度区域化特征明显。在邻近白令海峡的楚科奇海南部海域(R01、R02和CC1站),次表层10 m以下至底层的浓度明显高于表层,叶绿素a浓度最大值出现在底层和近底层。在调查海域中部,叶绿素a浓度最大层则分布在次表层10~20 m附近。在调查海域北部阿拉斯加巴罗近岸,表层及次表层叶绿素a浓度极低,底层和近底层出现叶绿素a浓度高值。而在最北部冰层密集区的R05站,观测到浮游植物水华发生,表层叶绿素a浓度高达24.00 mg/m3,在20 m以浅水体中分布均匀,20 m以深浓度迅速降低。从表1所示各层次叶绿素a浓度平均值看,叶绿素a浓度随深度增加而升高,10 m层以下至底层的浓度明显高于表层,底层(50 m)出现最大值(2.65± 4.09)mg/m3。
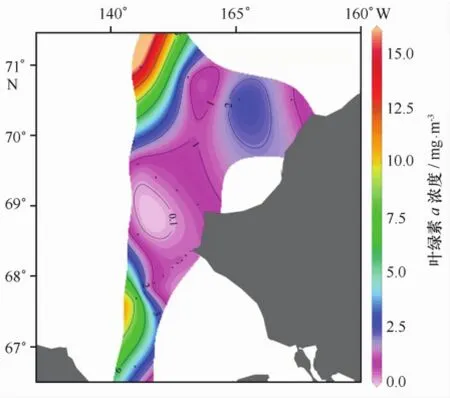
图2 2012年7月楚科奇海水层平均叶绿素a浓度平面分布Fig.2 Spatial distribution of integral average Chl a concentration in July 2012
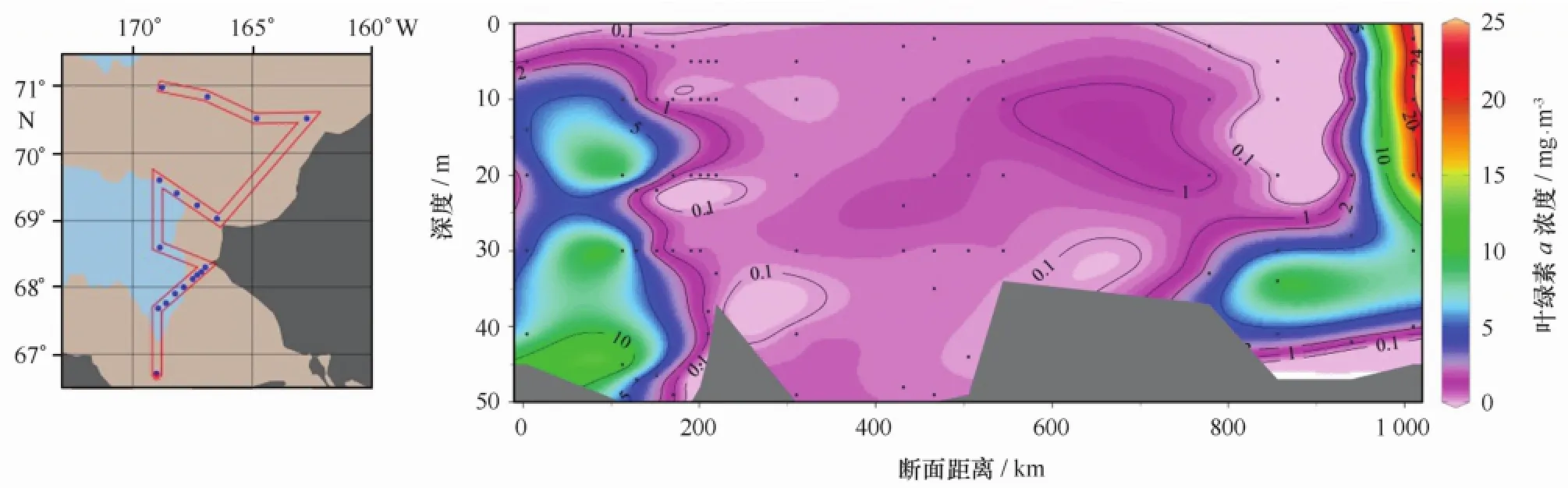
图3 2012年7月楚科奇海叶绿素a浓度垂直分布Fig.3 Vertical distribution of Chl a concentration in July 2012
返程时,SR和M断面的叶绿素a浓度垂直分布见图4。由图可见,随着调查船自北部深海平原向南航行,叶绿素a浓度逐渐升高,陆架区浓度明显高于陆架以外的深海海域,水层平均叶绿素a浓度最大值出现在SR03站,为1.52 mg/m3。深海区叶绿素a浓度的次表层最大值现象非常明显,40 m和50 m层叶绿素a浓度达到最大,分别为(0.73±0.46)mg/m3和(0.73±0.52)mg/m3(表1)。陆架区叶绿素a浓度的垂直分布稍显复杂,SR03和SR05站的叶绿素a浓度最大值均出现在底层,而邻近的SR07站和SR09站则相反,表层叶绿素a浓度高于表层以下水体中的叶绿素a浓度。
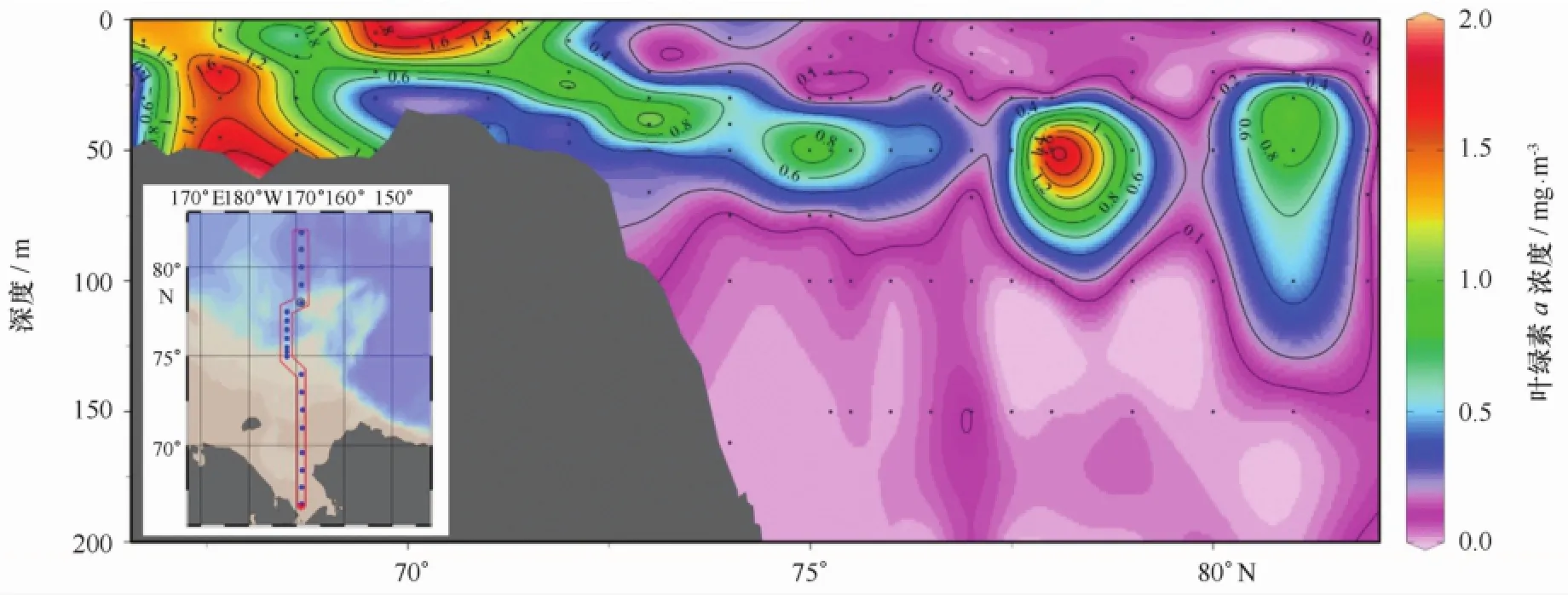
图4 2012年9月楚科奇海及邻近海域叶绿素a浓度垂直分布Fig.4 Vertical distribution of Chl a concentration in September 2012
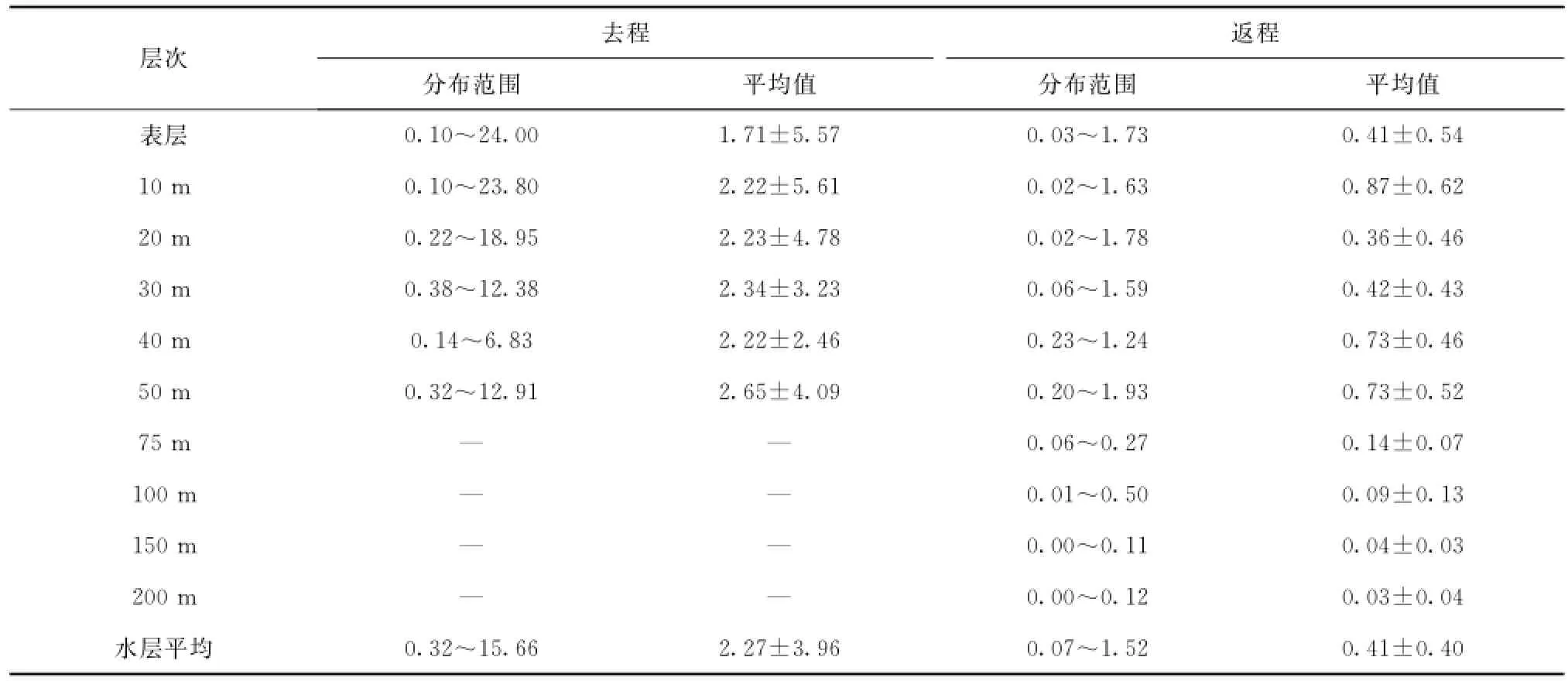
表1 2012年夏季楚科奇海及其邻近海域叶绿素α浓度(mg/m3)垂直分布Tab.1 The Chlαconcentration(mg/m3)of different water layer in Chukchi Sea and the adjacent area during the summer of 2012
3.1.2 粒度分级叶绿素a浓度
浮游植物粒度分级调查结果表明,在生物量较低的深海区,叶绿素a浓度的粒级结构以Pico级份占优势(其贡献率为46.1%),Net级份和Nano级份对总叶绿素a浓度的贡献差异极小,分别为26.6%和 27.3%。而在生物量较高的水深小于200 m的陆架区,Net级份浮游植物所占比重最高,Pico级份次之,Nano级份最低,分别为59.8%、27.9%和12.3%。
不同海域水层中叶绿素a粒级结构的垂直分布见图5。在陆架区50 m以浅水体中叶绿素a浓度均较高,各级份对总叶绿素a浓度的贡献情况变化不大,由大到小依次为Net级份、Pico级份、Nano级份,在叶绿素a浓度较高的10 m和50 m层,Net级份所占比重分别达到62.1%和66.8%。在深海区,Pico级份完全取代Net级份的优势地位,成为总叶绿素a浓度的主要贡献者。Pico级份叶绿素a在50 m层叶绿素a浓度最大层达到最大值0.26 mg/m3,但由于Net和Nano级份生物量同时增大,其所占比重相对其他层有所降低(39.9%)。
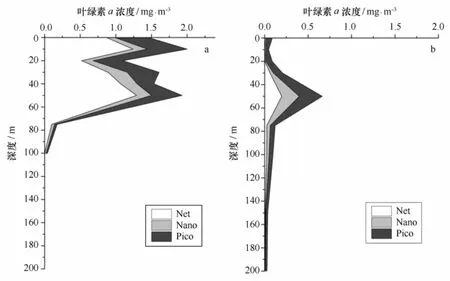
图5 2012年夏季不同海域粒度分级叶绿素a浓度平均值垂直分布Fig.5 Vertical distributions of size-fractionated Chl a concentration in different regions during the summer of 2012
3.2 初级生产力
3.2.1 初级生产力分布
浮游植物现存量高的陆架区初级生产力也较高。调查结果显示,观测海区初级生产力分布范围为12.31~943.28 mg/(m2·h),平均为(178.06± 299.48)mg/(m2·h)。在叶绿素a浓度最高的R05站初级生产力出现调查海域的最高值,陆架区5个站位的初级生产力(红色显示)均较高,返程时4个站位的初级生产力(蓝色显示)则较低,SR18站低至12.31 mg/(m2·h)(见图6)。
在初级生产力的垂直分布上(见图7a、7b),除返程SR05站以表层初级生产力最大外,其他各站的最大值基本出现在光衰减至3%~50%的次表层水。陆架区真光层深度普遍较深,约为40~50 m,而实测平均水深为56 m,可知光照可直达近底层或底层,深海区SR18和M02站真光层深度则深达90 m和100 m。R05站例外,该站真光层深度仅15 m,是由于Net级浮游植物的迅速旺发造成了水体透明度的降低。除了真光层底部,该站各个光衰减层次的初级生产力均高出其他站位平均水平十倍不止。
3.2.2 初级生产力粒级结构
从粒度分级初级生产力测定结果(见表2)来看,在陆架区以Net级份对总初级生产力的贡献为最大(60.6%),Pico(32.2%)和Nano级份(7.2%)次之,深海区则以Pico级份所占比重为大,比重由大到小依次为Pico(56.9%)、Net(23.8%)和Nano级份(19.3%)。初级生产力粒级结构分布特征与叶绿素a粒级结构基本一致。
在垂直分布上,Net与Pico级份在各光衰减层上对总初级生产力的贡献基本上呈互为消长的关系,随着深度的增加,Net级份的贡献先减少后增大,Pico级份则在光衰减至30%和10%层次贡献最大,Nano级份所占比重随深度变化不大。
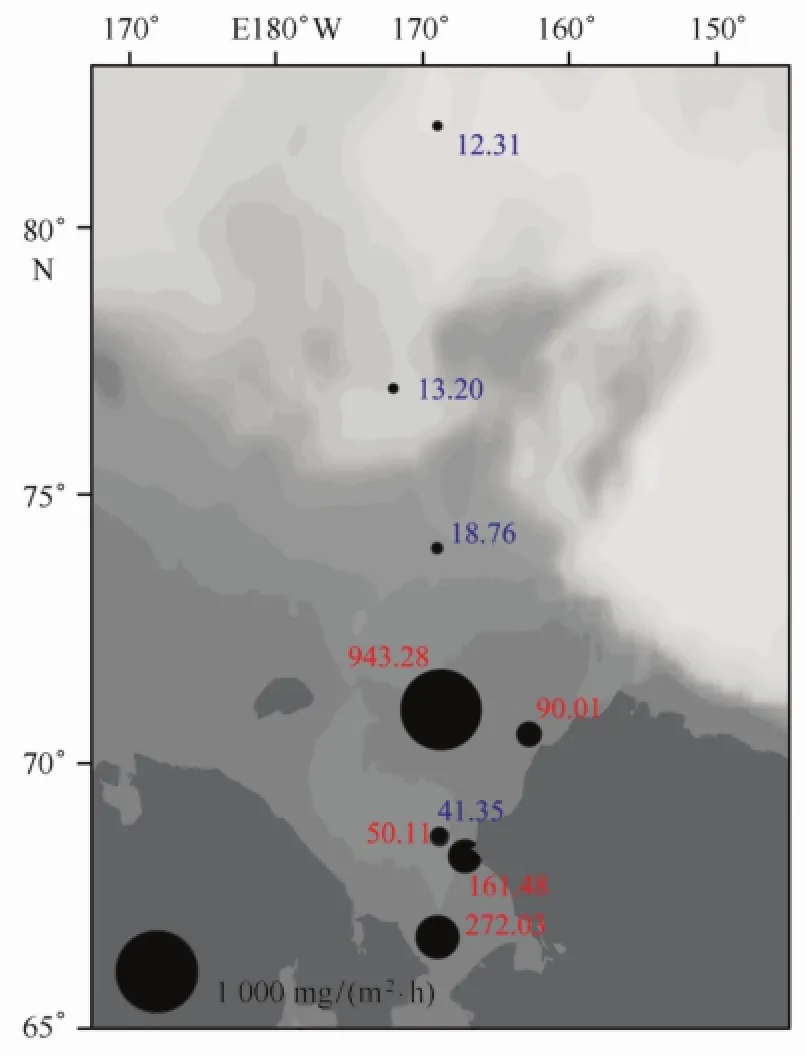
图6 2012年夏季楚科奇海及其邻近海域初级生产力[(mg/(m2·h)]的平面分布Fig.6 Spatial distribution of primary production[(mg/(m2·h)]in the Chukchi Sea and the adjacent area during the summer of 2012
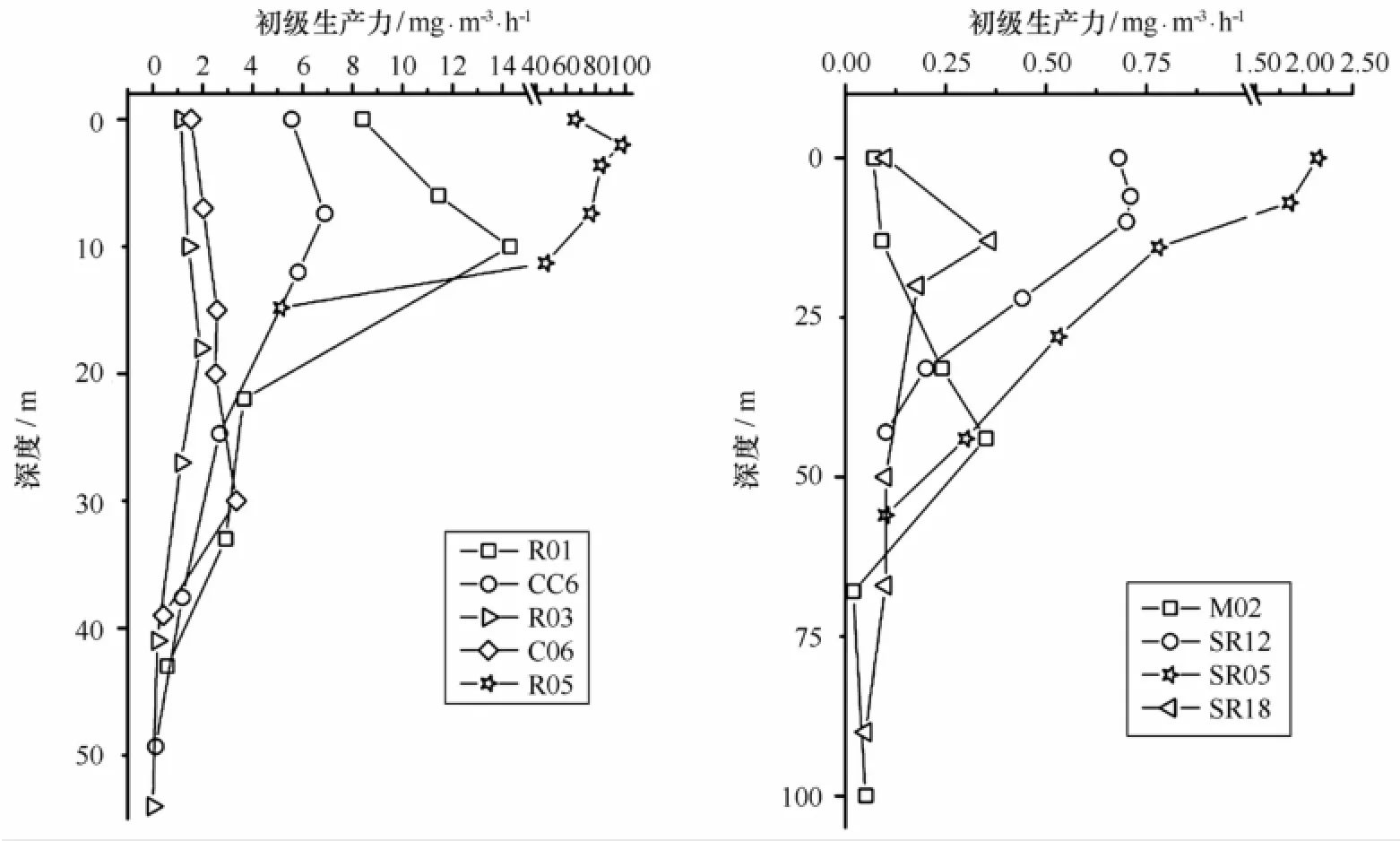
图7 2012年夏季楚科奇海及其邻近海域初级生产力[(mg/(m3·h)]的垂直分布Fig.7 Vertial distributions of primary production[(mg/(m3·h)]in the Chukchi Sea and the adjacent area during the summer of 2012
观测站位的光合作用同化数(以Chl a转化为C的值计)平均为(2.19±1.65)h-1,各级份中以Pico级份同化数为最高,但与Net、Nano级份差别不大(见表2)。在不同区域各级份同化数之间有较大差别,深海区Net级份高于Nano级份高于Pico级份,陆架区则相反。陆架区的平均同化数高于深海区。
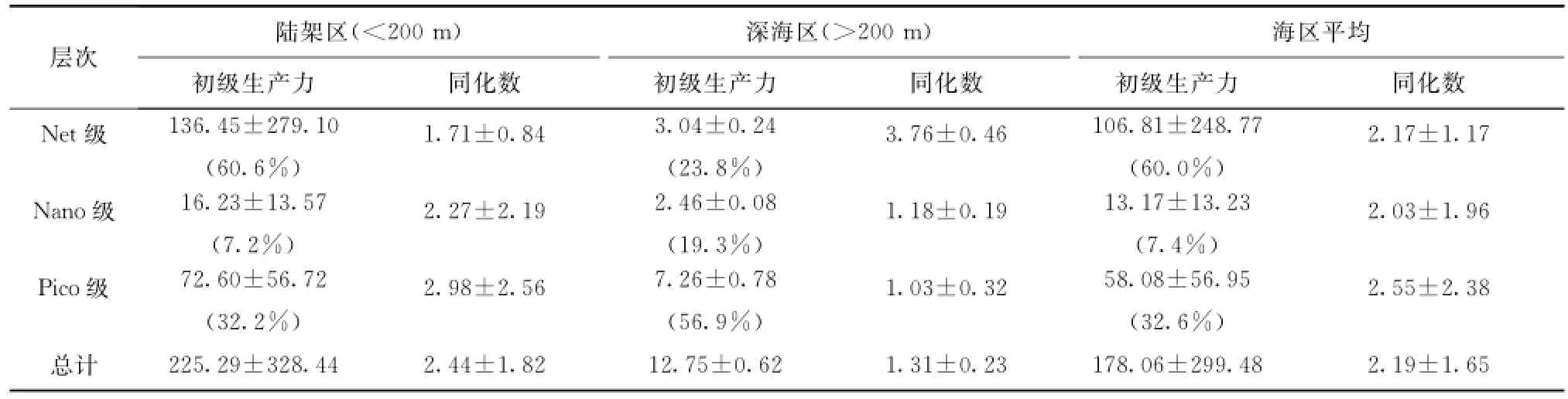
表2 楚科奇海及其邻近海域初级生产力[(mg/(m2·h)]粒级结构和同化数(h-1)Tab.2 Size structure of primary production[(mg/(m2·h)]and assimilation number(h-1)in the Chukchi Sea and adjacent area
4 讨论
4.1 调查海域环境特征
北极海域的浮游植物叶绿素和初级生产力存在极高的空间变异性[12],其中叶绿素的浓度变化范围为0.0~40.0 mg/m3,初级生产力为0.004~44.2 mg/(m3·h),二者的最高值均出现在楚科奇海。可以说楚科奇海是北极海域初级生产量的重要贡献者。本次调查结果同样显示出楚科奇海陆架区高生物量和高初级生产力的特征。
调查海域从白令海峡北口的楚科奇海向北延伸至其北部平原区、门捷列夫海脊区,直至深海,其西部与东西伯利亚海相连,东部至波弗特海,北部到达81°54'N。南北跨越15°(66.7°~81.9°N),东西跨越10°(162.8°~172.0°W),水深30~3 400 m。由于白令海峡两端的水位差,驱动北太平洋水由此进入楚科奇海,而海域中部凸起的Herald浅滩对入流水的流动和海冰融化过程有着显著影响[13]。一般认为,太平洋入流水分3个水团进入楚科奇海,它们是西侧高盐低温、高营养盐的阿纳德尔水,东侧低盐高温、低营养盐的阿拉斯加沿岸流和海峡中间的白令海陆架水[14]。各个水团的覆盖范围存在较大的季节和年际变化[15]。2012年夏季楚科奇海陆架区中部即阿拉斯加Pt.Hope近岸海域表层水温在6℃以上,明显高于北部和南部。这反映了陆架区中部受阿拉斯加沿岸流的影响,表现出明显的高温、低盐(小于31)、低营养盐(NO3-浓度小于0.5μmol/L、PO34-浓度小于0.6μmol/L)的特征。在南部靠近白令海峡海域,营养盐得到相对低温(小于5℃)、高盐(大于32)的白令海陆架水的补充,而北部海域受融冰过程影响,尤其是R05站,表层盐度极低(小于25),营养盐方面则由于冰雪融化带来较为丰富的养分(表层NO3-浓度和PO34-浓度约为1μmol/L)。
有研究表明[13],在楚科奇海存在2个不同的水团,一个位于海冰覆盖区,是早期进入楚科奇海的水团,以盐度高于33,温度低于-1℃,硅酸盐含量高于20μmol/L为特征,属于来自白令海深海盆的阿纳德尔水。另一个位于开阔水,是后期进入楚科奇海的水团,以盐度低于32.8,温度高于-0.3℃,硅酸盐含量低于10μmol/L为特征,属于来自白令海陆架的白令海陆架水。2个水团的分界线与海冰边缘线大体一致,在融冰过程中阿纳德尔水团随海冰边缘线一起向北退缩。本次调查中R断面的理化和生物参数的垂直分布特征显示出这2个水团对于调查海域浮游植物叶绿素a浓度和生产力空间分布的影响。强温跃层出现在R05站以南的15~20 m水层,这与以往研究结果一致[13]。而在R02站,盐度等值线向上抬升,硝酸盐和磷酸盐浓度均在此处出现高值,说明该站可能存在底层海水涌升现象,富含营养物质(NO3-浓度大于10μmol/L、PO34-浓度大于1.5μmol/L)的底层水上升至温跃层下方(见图8)。
4.2 不同海域浮游植物的现存量和生产力及其环境调控
楚科奇海浮游植物生长受众多因素的影响,呈现复杂的综合调控机制。在具有不同环境特征的海域,特定因素成为主要制约因子,因此不能以一种简单调控模式加以解释。调查海域跨越深海平原区和水深较浅的陆架及浅滩区,对于浮游植物的主要影响因素也必然需要结合环境特征进行分析。
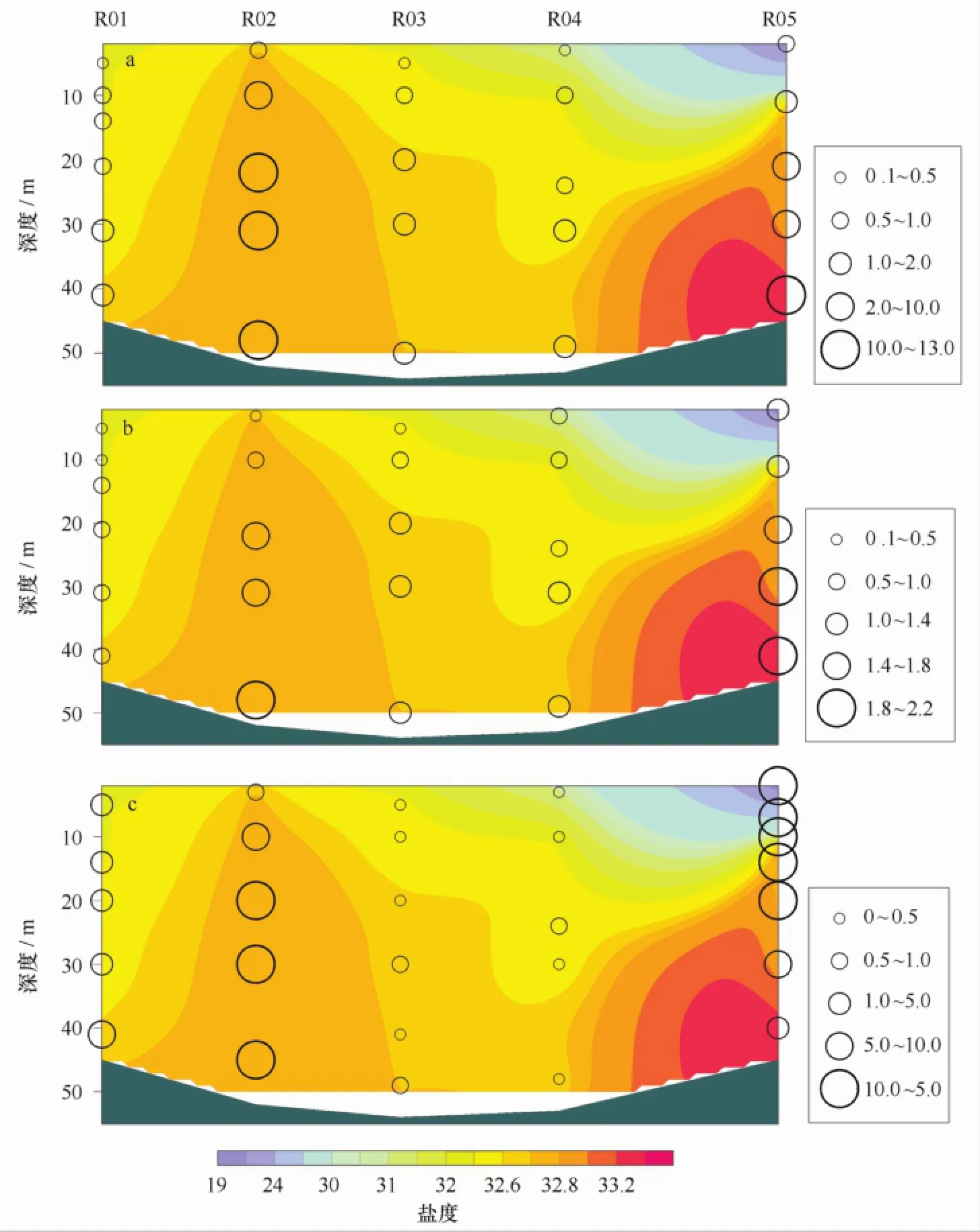
图8 2012年夏季楚科奇海R断面硝酸盐(a)、磷酸盐(b)浓度(μmol/L)和叶绿素a浓度(c)(mg/m3)在等盐线图上的垂直分布Fig.8 Vertical distributions of nitrate(a),phosphate(b)(μmol/L)and Chl a(c)(mg/m3)concentrations over the isohaline map of Section R
在楚科奇海域,浮游植物生长所需的营养物质主要来源于经白令海峡流入北冰洋的太平洋水[16],其所携带的溶解无机氮是控制西北冰洋海洋初级生产力的关键因素[17]。陆架区营养盐分布格局受不同水团分布范围的影响非常明显,并进一步影响到浮游植物生物量和初级生产力及其粒级结构特征。陆架区中部受阿拉斯加沿岸流影响,营养盐浓度较低,浮游植物生长受营养盐限制,大部分站位叶绿素a浓度低于0.50 mg/m3,R03站初级生产力以碳计仅50.11 mg/(m2·h),其中Pico级份对生物量和生产力的贡献最大,这是因为在贫营养海域,小粒径浮游植物个体微小,比表面积大,能量转换效率高,相比大粒径浮游植物具有更高的营养盐吸收效率和更快的生长率[18]。Cottrell和Kirchman[19]的研究结果表明,微型浮游植物的旺发带来丰富的溶解有机物,加上相对较高的水温,使得随着太平洋暖水以及沿岸径流一起进入楚科奇海的聚球藻迅速生长。本次调查中CC6站较高的初级生产力水平也主要由Pico级份贡献(占该站总初级生产力的67%),该级份光合作用同化数高达8.68 h-1,说明该站微微型浮游植物群落处于适宜的生长状态,生产力高。楚科奇海南部海域营养盐得到白令海陆架水的补充,表层叶绿素a浓度稍高于中部(大于1.0 mg/m3),R01站初级生产力(以碳计)为272.03 mg/(m2·h)。R02站的底层海水涌升现象,使得浮游植物特别是Net级份浮游植物旺发,可供养该区域浮游动物的生长繁殖[20—21]。值得注意的是该区融冰较早,上层海水在经历融冰期较高初级生产力和融冰水的稀释作用后营养盐浓度降低[17],加之强温跃层的影响,使得上层水体中营养盐得不到补充,浮游植物生物量仍然处于较低水平,但由于陆架区水深较浅而水体透明度较高的原因,光照可直达底层或近底层,温跃层以下水体中的浮游植物特别是Net级份在光照充足、营养盐丰富的条件下迅速旺发,造成R01、R02站下层水体中叶绿素a浓度远高于上层水体的现象。从相关性分析结果(表3)可以看到,陆架区叶绿素a浓度与各项营养盐浓度之间呈显著的正相关关系,而与温度、盐度成负相关;初级生产力和温度的相关则不明显。这一结果与Worden-Andersen[22]的结论相符,他认为在各类环境因子中温度对初级生产力的限制似乎有不确定的一面,营养盐和光照往往是刺激浮游植物水华旺发的主要因子。在陆架区北部冰缘区,我们在R05站捕捉到一次水华现象,该站表层叶绿素a浓度高达24.00 mg/m3,初级生产力达到943.28 mg/(m2·h),这是因为融冰过程带来充足养分,且冰藻的释放对水华有叠加和“播种”作用[23]。有研究指出冰藻对北冰洋中部初级生产力的贡献可占到一半以上[24],水华与海冰消融后退之间存在强烈的生物物理学联系[25—26],已发现水华的发生测站顺序基本上随冰缘逐渐向北退缩而移动[24,27]。
浮游植物在深海区与陆架区具有截然不同的环境调控机制,从表3可以看到,深海区叶绿素a浓度和初级生产力和各项理化环境参数之间的相关性并不显著。在深海区,海冰融化后,开阔海水对太阳辐射有更强的吸收能力[13],导致上层海水温度较高(50 m以浅水体温度高于-1℃),而融冰同时造成表层水体的盐度下降(50 m以浅水体盐度低于32),形成强大的温盐跃层,水体垂直稳定度极高,上下层水体之间的垂直交换受阻[28],叶绿素a浓度的垂直分布表现出明显的次表层最大值现象,最大值出现在温盐跃层底部及营养盐跃层以上,即50 m层附近,该跃层处营养盐和光合作用可利用光达到最佳权衡,有利于浮游植物生长(SR14站50 m层叶绿素a浓度高达1.93 mg/m3,Net和Nano级份所占比重大幅增加),这也可解释为该处异养浮游细菌丰度增加的重要原因之一[28]。第四次北极考察结果表明[29],高纬度冰水界面表层水营养盐状况表现为显著的N限制。本次考察中跃层以上水体显然同样受营养盐限制,叶绿素a浓度低于0.2 mg/m3,跃层以下营养盐不再受限,但光合作用可利用光不足,叶绿素a浓度迅速降低。各粒级中Pico级份对总叶绿素a浓度和总初级生产力的贡献最大,这与第三次北极考察所得结果一致[6],认为微微型光合浮游生物是北冰洋深海区浮游生物群落中的重要组成部分,其在北冰洋生态系统的物质循环和能量流动中起着重要作用。

表3 叶绿素α浓度和初级生产力与环境因子的Pearson相关性Tab.3 Pearson correlation analysis between Chlαconcentration,primary production and environmental factors
4.3 R断面往返观测结果比较
R断面从南部连接白令海峡处的北向延伸断面,也是历次北极科考的传统调查断面。表4列举了7月份去程时R断面和9月份返程时SR断面重复观测站位在两次观测期间各个参数的变化。
由表1可见,去程叶绿素a浓度远高于返程,R断面上叶绿素a分布呈现两端极高中间极低的特征(见图2),SR断面各站位之间叶绿素a浓度差距不大(见图4),但二者在叶绿素a浓度的垂向分布特征上非常一致,均表现出南部高值出现在近底层或底层,北部高值出现在表层和次表层的特征。这与第二次和第三次北极考察的结果[6—7]并不吻合,初步分析与水文条件及营养盐分布的年际变化有关。如4.1节所述,南部R02站出现底层水涌升现象,其复测站位SR03的各项理化参数同样显示出这一现象的存在(图未展示),且强度更大,低温(小于0℃)、高盐(大于33)、高营养盐(NO3-浓度大于10μmol/L、PO34-浓度大于2μmol/L)海水可直达20 m层,进而影响到次表层和表层,整个水柱中叶绿素a浓度高于1 mg/m3,高值区出现在底层,并延伸到SR05站的底层。而北部SR09站正处于海脊隆起上方,深层海水在运动过程中因受地形阻挡而产生由北向南的爬升,SR09站30 m层比20 m层水温低3.63℃,盐度高0.98,NO3-浓度高6倍,PO34-浓度高1倍。这一上升流的存在已得到以往考察结果的证实[6]。SR07和SR09站的上层水体可能受南北两个方向底层水涌升的推挤作用而将浮游植物聚集到较为温暖的表层和次表层,形成该二站20 m以浅水体中的叶绿素a高值区。尽管两次观测结果显示叶绿素分布格局相似,但不同粒径浮游植物在其中所发挥的作用截然不同。去程时由表至底Net级份占据了绝对优势(约80%),Pico次之,Nano级份所占比重最小;返程时各级份对叶绿素的贡献由大到小依次为:Pico级、Nano级和Net级,Net级份仅在底层贡献一半以上。从表4可以看出,各层次Nano和Pico级份叶绿素在两次观测期间变化并不大,只有Net级份急剧降低,可知该级份浮游植物对环境变化的灵敏度较高,须在最佳生态环境中才能快速生长,极易随环境改变而变化,说明楚科奇海的初级生产具有一定的脆弱性。
初级生产力的重复观测结果显示,SR05站生产力水平稍低于R03站,从垂向分布上可以看出初级生产力在R03站真光层中上部水体中分布比较均匀,而SR05站从表层往下,真光层内生产力迅速降低,从侧面反映出去程时调查海域内水体垂直稳定度好于返程时。另一方面从光合有效辐射(Photosynthetically A-vailable Radiation,简称PAR)数据(来源于NASA(http://oceancolor.gsfc.nasa.gov/)发布的MODIS L-3数据产品,时间精度为月平均,空间分辨率为4 km×4 km)可知,7月PAR以光量子计为(35.00±4.93)mol/(m2·d),是9月PAR[(14.95±1.80)mol/(m2·d)]的2倍多,返程时光强较弱,浮游植物生长受光限制,初级生产力较低。在光照较好、水体较稳定的7月,浮游植物可在一定深度水体内维持较高的生产力。初级生产力指数的垂直分布可佐证这一观点,R03站的初级生产力指数远高于SR05站,说明去程时浮游植物处于较好的生长状态,生理活性水平较高,固碳能力强。
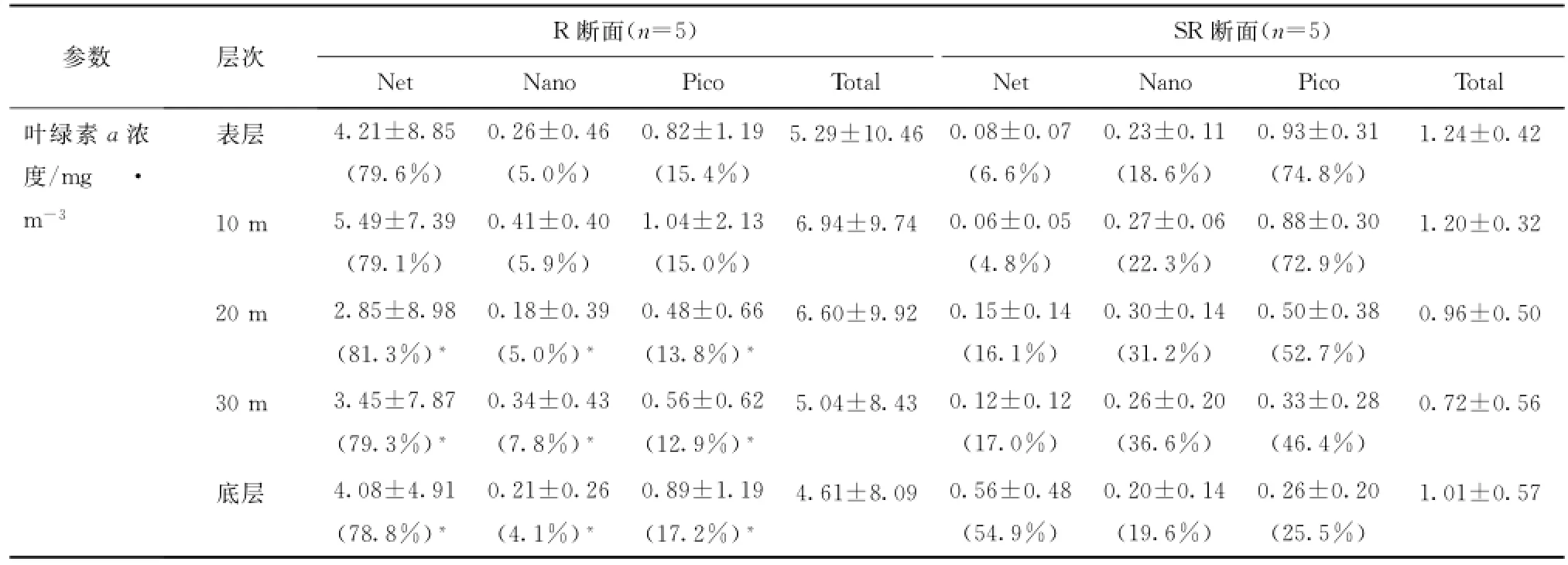
表4 楚科奇海陆架区R断面和SR断面各参数垂直分布Tab.4 The Chlαconcentration,primary production and assimilation number of different water layer in Section R and SR
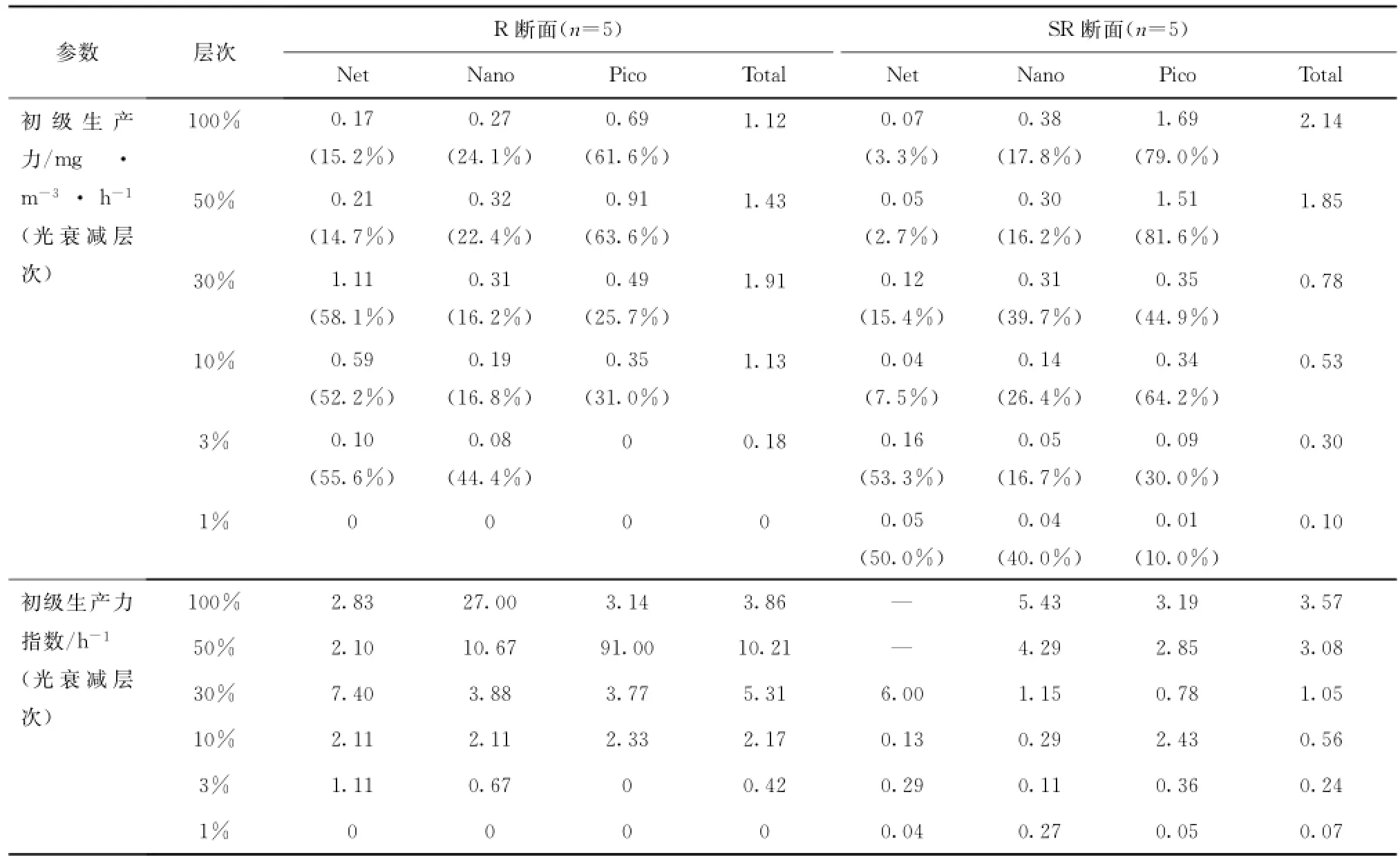
续表4
5 结论
在2012年夏季对楚科奇海及其邻近海域的浮游植物现存量和初级生产力进行了调查,结果如下:
调查海域浮游植物现存量和初级生产力的区域性差异显著。深海区因受强大温盐跃层的影响,浮游植物现存量和初级生产力均较低,具有明显的次表层最大值现象。陆架区因有太平洋入流水补充营养盐,因此叶绿素a浓度和初级生产力均较高,但由于不同水团的影响,同样表现出明显的区域性特征:南部有白令海陆架水补充营养物质,浮游植物现存量和生产力较高;中部受阿拉斯加沿岸流影响,浮游植物生长受营养盐限制,叶绿素a浓度低于0.5 mg/m3;北部冰缘区则因为融冰过程补充养分以及冰藻被大量释放入海,导致浮游植物迅速旺发发生水华,叶绿素a浓度(15.66 mg/m3)和现场初级生产力[943.28 mg/(m2·h)]达到本次调查最高值。
粒度分级Chl a和初级生产力结果显示,Net级份浮游植物对陆架区浮游植物现存量和生产力贡献最大,而粒径小于2μm的Pico级份浮游植物在深海区占优。
R断面往返观测结果显示,去程时浮游植物现存量和初级生产力远高于返程时。值得注意的是,本次调查发现楚科奇海南部海域和北部陆架坡折区均存在底层水涌升现象,这对浮游植物现存量和生产力及其在水层中的分布起着重要的影响作用,建议以后工作中加大对这一特殊区域的研究力度,以更好地了解海域中物理、化学、生物之间的耦合关系。
[1] Parkinson C L,Cavalieri D J,Gloersen P,et al.Arctic sea ice extents,areas,and trends,1978-1996[J].Journal of Geophysical Research:O-ceans(1978-2012),1999,104(C9):20837-20856.
[2] Comiso J C.A rapidly declining perennial sea ice cover in the Arctic[J].Geophysical Research Letters,2002,29(20):17-1-17-4.
[3] Holland M M,Bitz C M,Tremblay B.Future abrupt reductions in the summer Arctic sea ice[J].Geophysical Research Letters,2006,33(23):L23503.
[4] Wang M Y,Overland J E.A sea ice free summer Arctic within 30 years?[J].Geophysical Research Letters,2009,36(7):L7502.
[5] Arrigo K R,Van Dijken G,Pabi S.Impact of a shrinking Arctic ice cover on marine primary production[J].Geophysical Research Letters,2008,35(19):L19603.
[6] 刘子琳,陈建芳,刘艳岚,等.2008年夏季西北冰洋观测区叶绿素a和初级生产力粒级结构[J].海洋学报,2011,33(2):124-133.
[7] 刘子琳,陈建芳,张涛,等.楚科奇海及其海台区粒度分级叶绿素a与初级生产力[J].生态学报,2007,27(12):4953-4962.
[8] Holm-Hansen O,Lorenzen CJ,Holmes R W,et al.Fluorometric determination of chlorophyll[J].ICESJournal of Marine Science,1965,30(1):3-15.
[9] Poole H H,Atkins W R G.Photo-electric measurements of submarineillumination throughout the year[J].Journal of the Marine Biological Association of the United Kingdom(New Series),1929,16(1):297-324.
[10] Evans C A,O'Reilly J E,O'Reilly J E,et al.A handbook for the measurement of chlorophyll a and primary production[M]//Biological Investigations of Marine Antarctic Systems and Stocks(BIOMASS).Texas:Texas A and M University,1987:8,114.
[11] Ning X R,Vaulot D,Lin ZS,et al.Standing stock and production of phytoplankton in the estuary of the Changjiang(Yangste River)and the adjacent East China Sea[J].Marine Ecology-Progress Series,1988,49(10):141-150.
[12] Rao D S,Platt T.Primary production of Arctic waters[J].Polar Biology,1984,3(4):191-201.
[13] 赵进平,史久新,金明明,等.楚科奇海融冰过程中的海水结构研究[J].地球科学进展,2010,25(2):154-162.
[14] Grebmeier J M,Mcroy C P,Feder H M.Pelagic-benthic coupling on the shelf of the northern Bering and Chukchi seas.1.Food supply source and benthic bio-mass[J].Marine Ecology-Progress Series,1988,48(1):57-67.
[15] Aagaard K,Weingartner T J,Danielson S L,et al.Some controls on flow and salinity in Bering Strait[J].Geophysical Research Letters,2006,33(19):L19602.
[16] Grebmeier J M,Cooper L W,Feder H M,et al.Ecosystem dynamics of the Pacific-influenced northern Bering and Chukchi Seasin the Amerasian Arctic[J].Progress in Oceanography,2006,71(2):331-361.
[17] 李宏亮,陈建芳,高生泉,等.西北冰洋中太平洋入流水营养盐的变化特征[J].海洋学报,2011,33(2):85-95.
[18] Parsons T R,Takahashi M.Environmental control of phytoplankton cell size[J].Limnology and Oceanography,1973,18(4):511-515.
[19] Cottrell M T,Kirchman D L.Photoheterotrophic microbes in the Arctic Ocean in summer and winter[J].Applied and Environmental Microbiology,2009,75(15):4958-4966.
[20] 林景宏,戴燕玉,张金标,等.夏季楚科奇海浮游动物的生态特征[J].极地研究,2001,13(2):107-116.
[21] 张光涛,孙松.2003年夏季西北冰洋浮游动物群落结构和地理分布研究[J].海洋学报,2011,33(2):146-156.
[22] Norden-Andersen O G.The Annual Cycle of Phytoplankton Primary Production and Hydrography in the Disko Bugt Area,West Greenland[M].Copenhagen:Commission for Scientific Research in Greenland,1981:68.
[23] Michel C,Legendre L,Therriault J,et al.Springtime coupling between ice algal and phytoplankton assemblages in southeastern Hudson Bay,Canadian Arctic[J].Polar Biology,1993,13(7):441-449.
[24] 杨清良,林更铭,林茂,等.楚科奇海和白令海浮游植物的种类组成与分布[J].极地研究,2002,14(2):410-521.
[25] Wang J,Cota G F,Comiso J C.Phytoplankton in the Beaufort and Chukchi Seas:distribution,dynamics,and environmental forcing[J].Deep-Sea Research Part li-Topical Studies in Oceanography,2005,52(24):3355-3368.
[26] Perrette M,Yool A,Quartly G D,et al.Near-ubiquity of ice-edge blooms in the Arctic[J].Biogeosciences,2011,8(2):515-524.
[27] Heimdal B R.Arctic ocean phytoplankton[M]//The Arctic Seas.US:Springer,1989:193-222.
[28] 张芳,何剑锋,郭超颖,等.夏季北冰洋楚科奇海微微型,微型浮游植物和细菌的丰度分布特征及其与水团的关系[J].极地研究,2012,24(3):238-246.
[29] 庄燕培,金海燕,陈建芳,等.北冰洋中心区表层海水营养盐及浮游植物群落对快速融冰的响应[J].极地研究,2012,24(2):151-158.
(1.Second Institute of Oceanography,State Oceanic Administration,Hangzhou 310012,China;2.Key Lab of Marine Ecosystems and Biogeochemistry State Oceanic Administration,Hangzhou 310012,China;3.State key Laboratory of Satellite Ocean Environment Dynamics,Hangzhou 310012,China;4.Ocean University of China,Qingdao 266003,China)
Size structure of standing stock and primary production of phytoplankton in the Chukchi Sea and the adjacent sea area during the summer of 2012
Le Fengfeng1,2,Hao Qiang1,2,3,Jin Haiyan1,2,3,Li Tao4,Zhuang Yanpei1,2,Zhai Hongchang1,2,Liu Chenggang1,2,3,Chen Jianfang1,2,3
Size fractionated standing stock and primary production of phytoplankton in the Chukchi Sea and the adjacent sea area were studied during the 5th Chinese National Arctic Research Expedition in the summer of 2012.The results clearly showed that there was marked feature of spatial zonation in the sea areas investigated.In the shelf region,the standing stock and primary production were much higher than those of deep-sea area.The water column integrated average values of Chl a concentration varied from 0.32 to 15.66 mg/m3[on average(2.77±3.96)mg/m3]during the outward voyage.The high values appeared in the southern Chukchi Sea,coast areas near Point Barrow of Alaska and northern ice edge area.The lowest and highest value of primary production(PP)were 50.11 mg/(m2·d)and 943.28 mg/(m2·d),and the high value was encountered in the ice edge where phytoplankton bloom happened.During the return voyage,the water column integrated average value of Chl a concentration was an order of magnitude lower than the outward voyage.The values of PP were 12.31~41.35 mg/(m2·d),with the high value appearing in the shelf region.The results of size-fractionated Chl a and PP showed that photosynthetic picoplankton was the greatest contributor to phytoplankton stocks and production in deep-sea area(46.1% and 56.9%respectively),and the differences between contributions of microplankton and nanoplankton to Chl a and PP were very small.But in the shelf region,microplankton took the dominant position of Chl a(59.8%)and PP(60.6%),picoplankton second and nanoplankton the minimum.
Chukchi Sea;Arctic;phytoplankton standing stock;primary production;size structure
P727;Q178.53
A
0253-4193(2014)10-0103-13
2014-02-16;
2014-04-26。
国家自然科学基金项目(41206181,41306162);南北极环境综合考察与评估专项(CHINARE2012-03-05,CHINARE2014-04-03)。
乐凤凤(1981—),女,浙江省舟山市人,助理研究员,主要从事海洋生态学研究。E-mail:le_ff@126.com
乐凤凤,郝锵,金海燕,等.2012年楚科奇海及其邻近海域浮游植物现存量和初级生产力粒级结构研究[J].海洋学报,2014,36(10):103—115,
10.3969/j.issn.0253-4193.2014.10.011
Le Fengfeng,Hao Qiang,Jin Haiyan,et al.Size structure of standing stock and primary production of phytoplankton in the Chukchi Sea and the adjacent sea area during the summer of 2012[J].Acta Oceanologica Sinica(in Chinese),2014,36(10):103—115,doi:10.3969/j.issn.0253-4193.2014.10.011
猜你喜欢
海洋通报(2022年2期)2022-06-30
石油地球物理勘探(2022年3期)2022-06-11
海洋石油(2021年3期)2021-11-05
河北环境工程学院学报(2021年1期)2021-03-19
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
焊接(2015年6期)2015-07-18
应用海洋学学报(2014年1期)2014-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
组合机床与自动化加工技术(2014年12期)2014-03-01
