
北极王湾2010年夏季水体营养盐分布及影响因素
2014-06-01 12:30季仲强高生泉金海燕何剑锋白有成王斌杨志陈建芳
海洋学报 2014年10期
季仲强,高生泉,金海燕,2*,何剑锋,白有成,王斌,杨志,陈建芳,2
(1.国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州 310012;2.卫星海洋环境动力学国家重点实验室,浙江杭州 310012;3.中国极地研究中心,上海 200136)
北极王湾2010年夏季水体营养盐分布及影响因素
季仲强1,高生泉1,金海燕1,2*,何剑锋3,白有成1,王斌1,杨志1,陈建芳1,2
(1.国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州 310012;2.卫星海洋环境动力学国家重点实验室,浙江杭州 310012;3.中国极地研究中心,上海 200136)
王湾海域位于北极斯瓦尔巴群岛西北侧,其水体特征主要受到北极冰川与大西洋的共同影响。为了研究冰川融水与大西洋水对王湾营养盐分布以及生态系统的影响,在2010年夏季黄河站考察期间,对王湾海水与地表径流进行采样分析,并测定了营养盐和叶绿素a。结果表明:2010年夏季王湾是大西洋水影响较弱的年份,以低温低盐高营养盐的本地变异水为主导。2010年7月王湾水体可以划分为4个水团,表层水(SW)、中层变异水(TIW)、本地变异水(TLW)和大西洋变异水(TAW),其中,表层水与中层变异水营养盐浓度较低,随着深度增加,本地变异水与大西洋变异水营养盐浓度都较高。大西洋变异水主要影响200 m以深水柱,其营养盐浓度低于本地变异水影响的底部值,本地变异水中营养盐浓度的增加主要来自于颗粒物中营养盐的再生。表层水主要来源于地表径流与冰川融水,这些淡水输入与浮游植物的初级生产作用共同决定了表层水的营养盐浓度水平及结构。本地变异水中在水深100 m上下出现的铵盐高值可能与浮游动物和/或微生物的代谢过程有关。
北极;王湾;营养盐特征;水团
1 引言
王湾(Kongsfjorden)(79°N,12°E周围区域)是挪威斯瓦尔巴群岛(Svalbard)西北部一个典型的峡湾海洋系统,湾内有大量冰川发育。湾中水体主要受大西洋水(Atlantic Water,AW)和北极水(Arctic Water,Ar W)两大水团的影响,它们在沿陆架北上过程中与峡湾(fjord)内部水体发生交换(见图1)[1—2]。王湾浮游生态系统对大西洋水与北极水在湾内的强弱变化最为敏感,而底栖生态系统主要受水文长期变化以及海冰、冰川径流与降水变化的影响[3]。在王湾内部,冰川与海水直接相连,冰川的输入会引起底栖生物,浮游动物与浮游植物的减少[3]。王湾内部海冰与冰川的消融以及大西洋水的入侵都与气候变化有关联,因而,大西洋与北极的气候变化信号将对峡湾系统的理化因子和生态环境产生可观测的影响[4]。
通过水体交换过程,富营养盐的大西洋水(AW)和北极水(Ar W)年际间的变化将引起湾内水体特征的改变[5—7],同时,海冰与冰川的消融也将引起湾内营养盐结构的变化,从而对湾内生产力的发展产生重要影响。目前国内有一些对王湾水体浮游植物与细菌群落等研究进行的报道[8—10]。而营养盐作为串联水体环境变化和生态系统演变的关键因子,目前王湾海域这方面的研究还较少[11—14]。本文拟通过2010年夏季王湾中海水及邻近陆地冰川消融产生径流水的采集,分析其中的营养盐与叶绿素等关键指标,并结合温度与盐度,讨论了北极海洋地区子系统营养盐分布与水团变化的关系,及其对气候变化引起的海流改变的响应。
2 研究区域概况及实验设计
2.1 区域环境
由于高温高盐的大西洋水AW带来大量的热量,使得斯瓦尔巴群岛陆架西侧全年基本无冰,形成较为典型的海洋性湿润气候。因而,沿着群岛西海岸,7月温度常常在1~10℃,夏季最高月平均气温约5℃,冬季最低月平均气温约-15℃。王湾沿东南向西北展布,长约20 km,而宽度在4 km到10 km范围之间(图1)。由于湾口有明显的冰川沟槽向陆架延伸,有利于峡湾内外水体交换,对王湾的水文、物理与生物等变化有决定性影响[1],使得峡湾内水团在不同季节下存在不同控制性水团[5]。首先,源于湾外的两个水团分别是WSC中的大西洋水(AW)和沿岸流中的北极水(Ar W),由锋面区域分隔;当AW进入王湾时,会与Ar W混合,从而,改变其水团性质,称为大西洋变异水(Transformed Atlantic Water,TAW);表层水(Surface Water,SW)在冰川消融时形成并在晚春与夏季占主导,层厚向着湾口会逐渐减小,同时,增强的太阳辐射使得水温升高,并有较大的变化范围;上层的SW与下层的AW或TAW混合形成中层水(Intermediate Water,IW);在秋季和冬季,由于表层的变冷和对流,两个水团将在湾内形成,即本地水(Local Water,LW)和冬季冷却水(Winter Cold Water,WCW),它们在春季与夏季逐渐与表层水混合,从而,在温盐特征上也向中层水IW靠近[1,5]。此外,夏季王湾周边有5处冰川消融直接进入湾内,是淡水的主要来源。入海淡水年平均总通量,包括冰川、雪融水、降水、径流和地下水,约占峡湾水体质量的5%[5]。所以,王湾水体中水团的变化也会带动营养盐特征的改变,从而影响湾内生态环境。
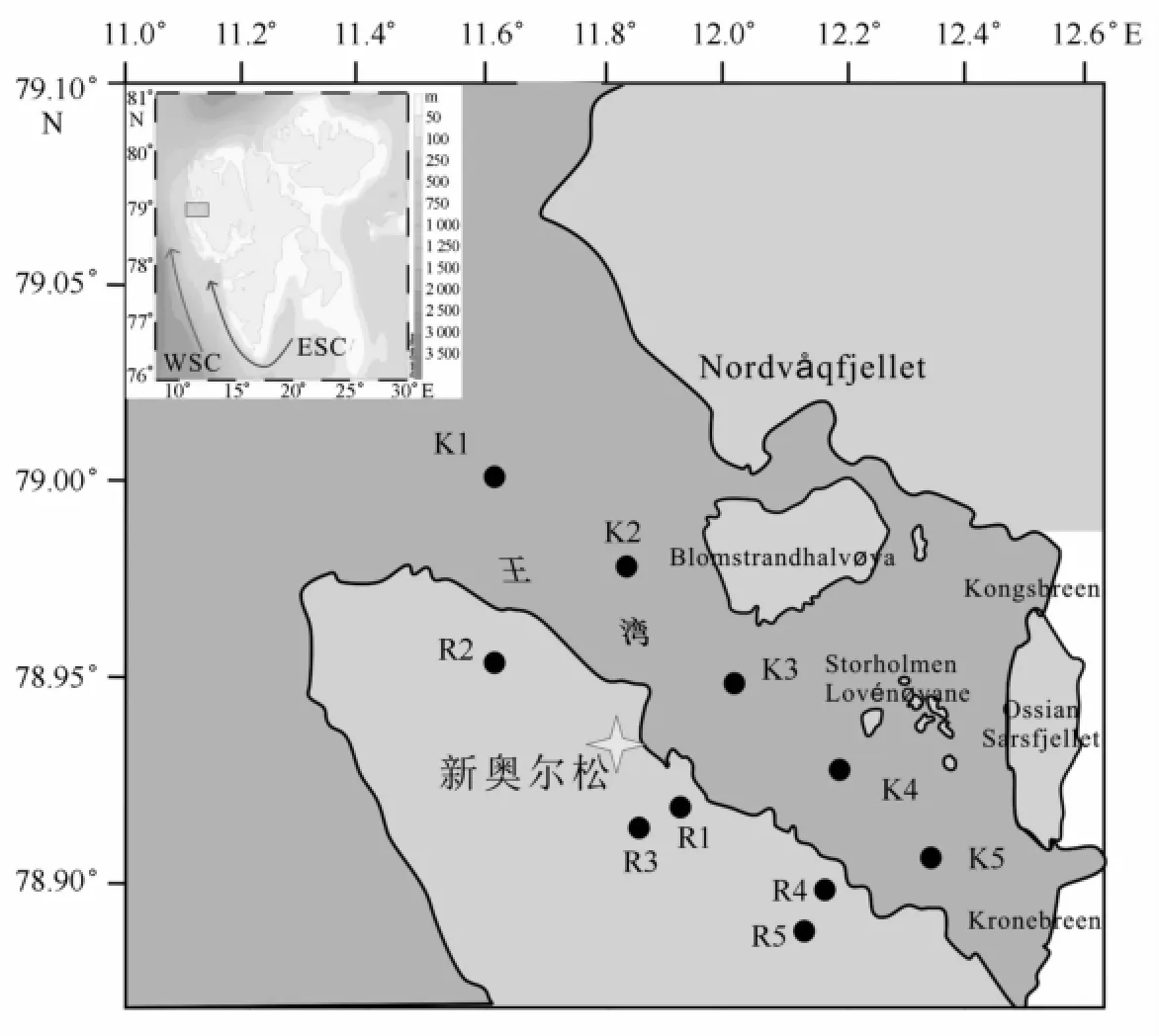
图1 2010年夏季王湾样品采集站位Fig.1 Sampling sites around Kongsfjorden in summer 2010
2.2 样品采集与分析
王湾地区所在的新奥尔松(Ny-Ålesund)是北极气候环境研究的一个重要考察平台,来自世界各地的科学家在这里开展着气候变化对北极地区生态环境影响的研究。中国于2004年在该地区建立了黄河站,并开始了长期的调查研究。2010年夏季,我们对王湾地区进行了海水与陆源径流水的采样,具体站位见图1与表1。海水采样层位分别为0 m、2 m、5 m、10 m、20 m、30 m、50 m、100 m、150 m和200 m,共45个海水样品。温度和盐度数据由CTD(SD204,SAIV A/S)获得。陆源径流水是直接采集表层水,共5个样品。
样品采集后在海洋实验室进行过滤和样品的收集工作。叶绿素a样品(GF/F膜,Whatman公司)冷冻保存于-80℃冰箱中,营养盐样品加氯仿并于-18℃下冷冻保存;所有样品带回国内实验室进行分析。叶绿素a使用唐纳荧光分析仪(Turner10-AU,美国)进行测定,检出限0.01μg/L。铵盐用723型分光光度计(靛酚蓝法)测定;其他营养盐使用营养盐自动分析仪(Skalar San++,荷兰)进行测定,其中,硝酸盐、亚硝酸盐、铵盐、磷酸盐和硅酸盐的检出限分别为0.1、0.01、0.01、0.03和0.1μmol/L。
3 结果
3.1 水文特征
图2是王湾海水调查断面的温度、盐度和密度分布图。盐度值在27.7到34.7范围,由于受到淡水输入影响,盐度值在表层较低,并随着深度的增加而增加。温度范围在-1.2℃到8.5℃之间。表层受到夏日极昼辐射的影响,温度较高;而在约30 m到200 m水深处温度较低(小于1℃);最低值出现在约50~120 m深度(小于-1℃)。位密度值的变化范围在21.6~28.7 kg/m3,表层密度变化较大,到下层则较为均匀。从位密度的分布可以看到,夏季水柱呈垂向分层结构,海水层化作用明显。
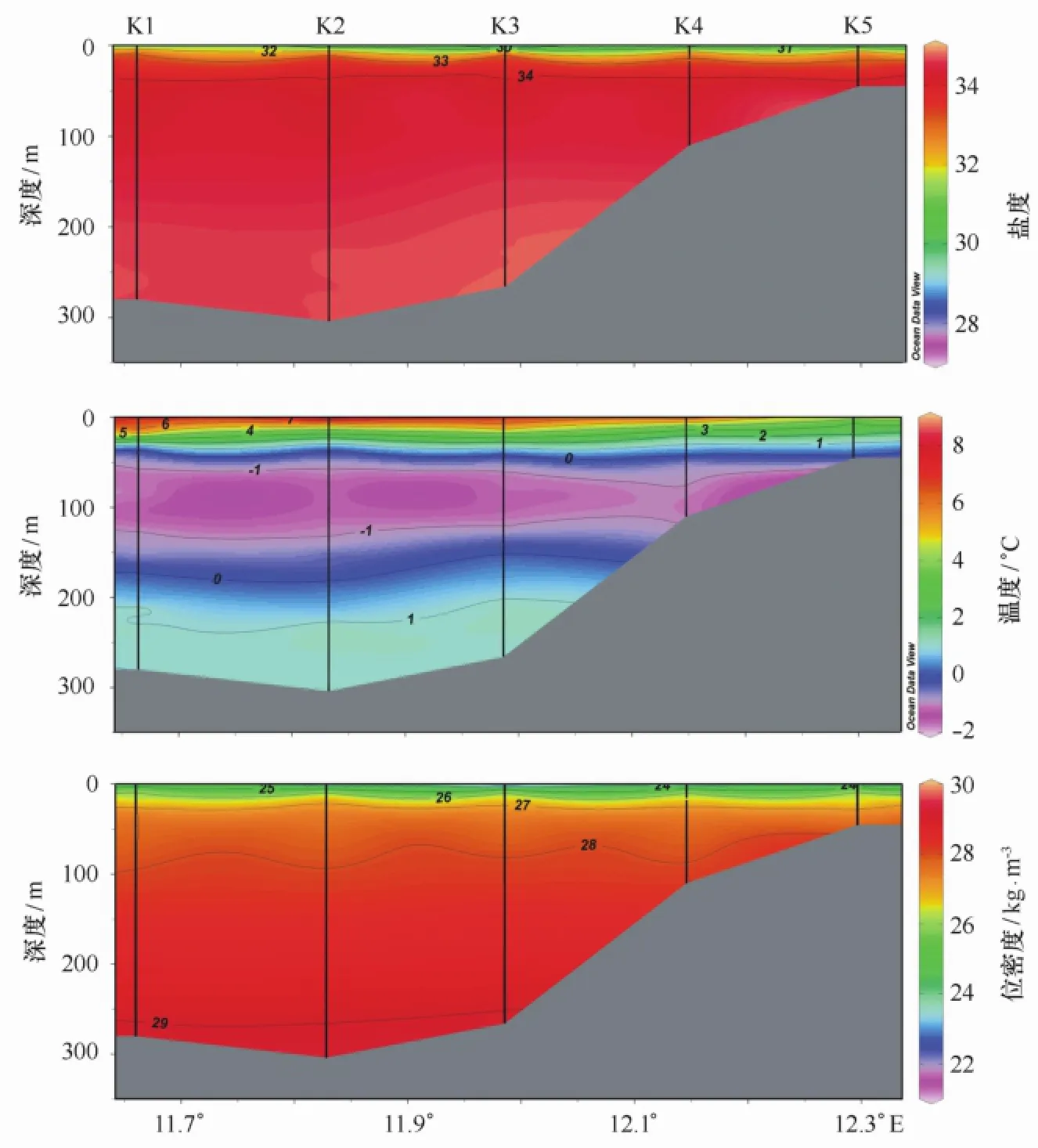
图2 2010年夏季王湾盐度、温度与位密度断面分布Fig.2 Profile distributions of salinity,temperature and potential density in Kongsfjorden,summer 2010
3.2 营养盐与叶绿素α分布
2010年7月王湾海水营养盐浓度范围分别为:硝酸盐(NO3-N,0.7~13.7μmol/L),亚硝酸盐(NO2-N,从低于检出限到0.56μmol/L),铵盐(NH4-N,0.25~2.67μmol/L),磷酸盐(PO4-P,0.13~1.53 μmol/L)和硅酸盐(SiO3-Si,1.1~6.7μmol/L)(图3)。其中,NO3-N表层浓度为0.7~2.0μmol/L,水平方向上呈湾内高湾外低的分布,垂向上随着深度的增加而增加,到150~200 m水深,浓度稳定在12.0~13.7 μmol/L之间;NO2-N浓度值基本在0.20μmol/L以下,在K2站20~50 m与K1和K2站100~150 m范围有较高浓度(0.30~0.56μmol/L);NH4-N浓度在50~150 m水深处较高,范围在1.50~2.67μmol/L,而水柱其他位置的浓度都较低;PO4-P浓度值随着深度不断增加,到100 m以深时呈现由陆架向湾内递减的趋势;表层SiO3-Si浓度值(2.5μmol/L)稍高于30~50 m水深的1.5μmol/L,并随着深度的增加而增加,但其浓度增加不明显。图4还显示了王湾水柱中营养盐结构的变化。氮磷比值与氮硅比值均呈现由表层向深层增加的趋势,分别由1.9增加到20.4和由0.2增加到2.6。
夏季,王湾水体叶绿素a的浓度较低(0.01~1.24μg/L),平均值仅0.45μg/L,基本在1.00μg/L以下,表明夏季浮游植物的现存量较低,且从图3可以看出,叶绿素a相对高值主要分布在50 m以浅范围,50 m以深叶绿素a浓度很低,均小于0.25μg/L。
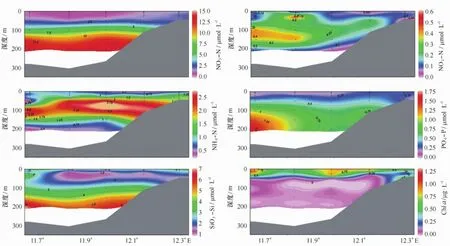
图3 2010年夏季王湾营养盐与叶绿素a断面分布Fig.3 Profile distributions of nutrients and Chl a in Kongsfjorden,summer 2010
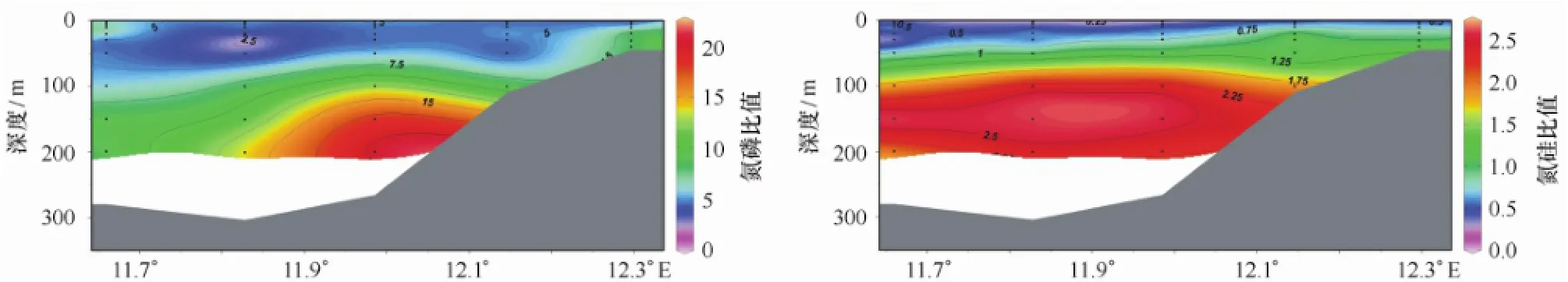
图4 2010年夏季王湾氮磷比值与氮硅比值断面分布Fig.4 Profile distributions of N/P ratio and N/Si ratio in Kongsfjorden,summer 2010
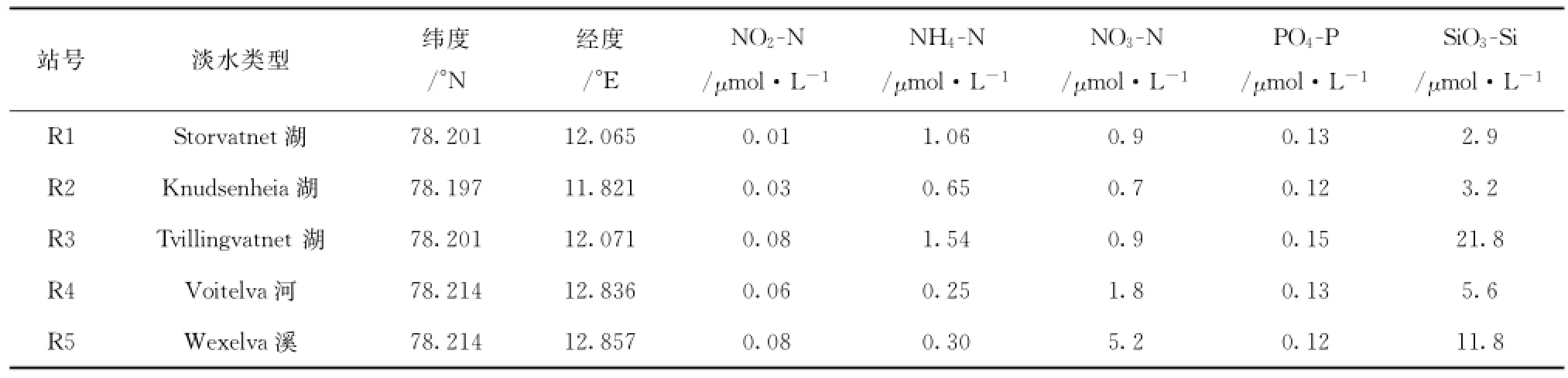
表1 陆地上采集样品站位及其营养盐含量Tab.1 Terrestrial sampling sites and their nutrient contents
表1是地表径流不同湖泊与河流水体中营养盐的浓度值。从表中可以看出,新奥尔松地表径流水中,湖水营养盐浓度总体较低,仅NH4-N与SiO3-Si浓度稍高,前者最高达1.54μmol/L,后者除了湖R3外,基本在3.0μmol/L左右;河水与溪流水中硝酸盐与硅酸盐浓度明显升高,最高可达5.2μmol/L和11.8μmol/L。
4 讨论
4.1 王湾海域水团及其营养盐特性
斯瓦尔巴群岛西部峡湾存在多个水团,其对应有不同温盐特征,同时营养盐和生物特征也各不相同。水文条件的变化对王湾区域营养盐及生物分布都有较大影响。2002年4、5月,Hodal等在王湾中部站位做连续观测发现,北极水ArW影响较大时,峡湾内旺发藻种以硅藻为主,而5月中旬,大西洋水AW控制时,棕囊藻或者与硅藻均占主要优势[12]。同样的,Willis等发现平流进入王湾水团与浮游动物群落结构的变化之间也有密切的关系[15]。AW进入峡湾的时间和范围不同还将影响王湾区域春季浮游植物旺发时间和面积[13]。
在Svendsen等的分类基础上[1],结合2010年王湾水体的温盐特征,将2010年7月王湾水体分为4个水团进行讨论,分别为表层水(SW)、本地变异水(TLW)、中层变异水(TIW)和大西洋变异水(TAW)(图5),同时,表2是这些水团对应的温度、盐度和营养盐特征。
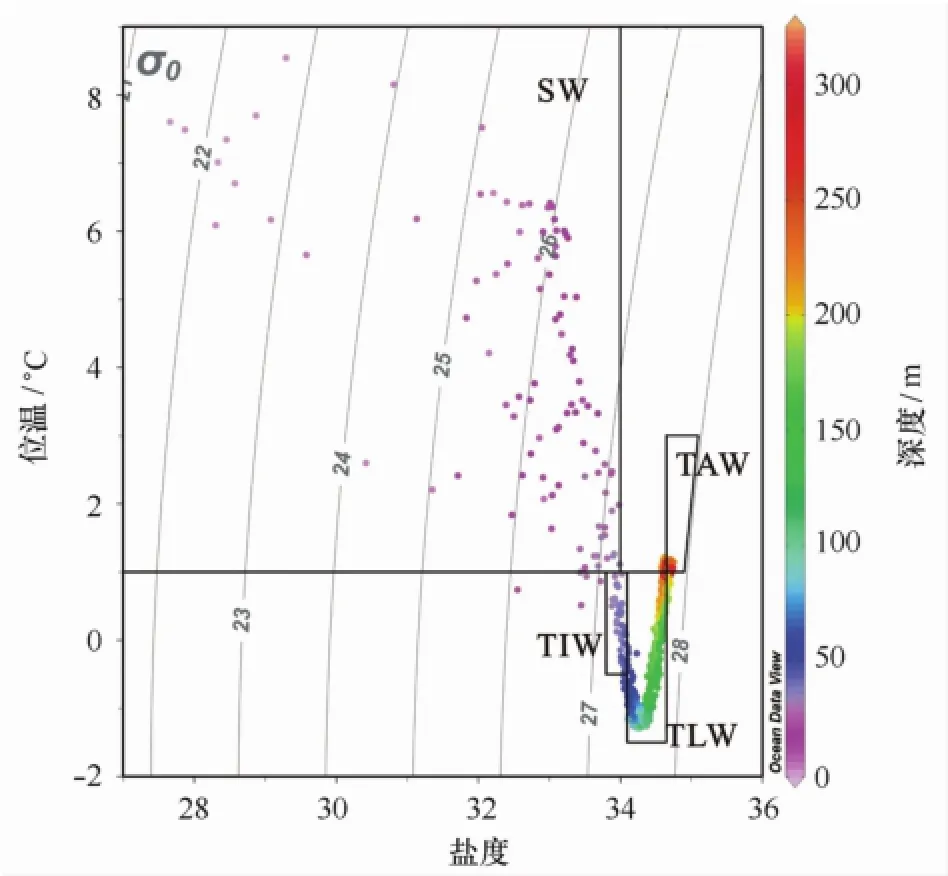
图5 王湾调查站位T-S图及其水团划分Fig.5 T-S diagram and the division of the water mass
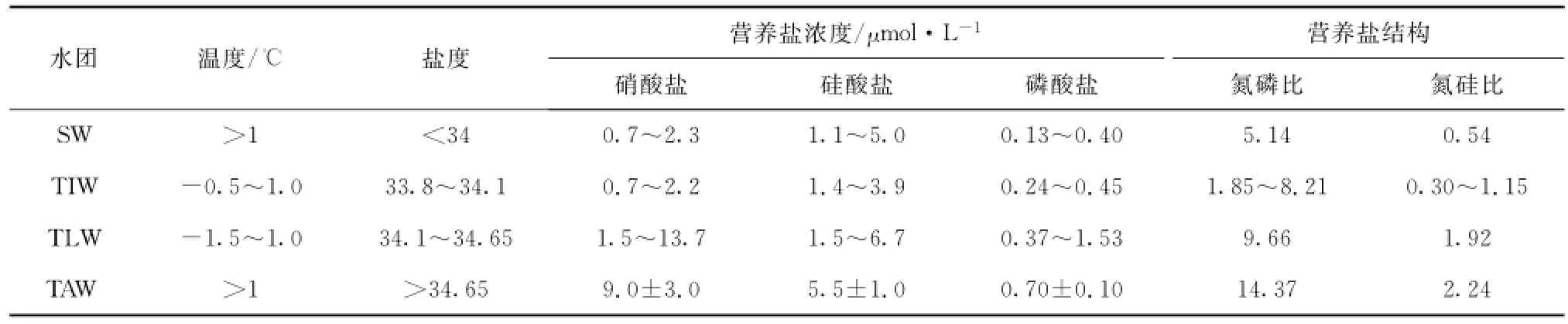
表2 王湾水团温度、盐度及其营养盐浓度和结构Tab.2 Temperature,salinity and nutrient concentrations and composition of the water masses in Kongsfjorden
夏季,冬季冷却水(WCW)与周围温暖的表层水混合转化,从而在表层以下不断的形成本地水(LW)[16];对应地,根据温盐特征,王湾50~200 m水深的低温水体是残留的WCW和LW的混合水体,将其命名为本地变异水(TLW,温度变化范围-1.5~1.0℃,盐度变化范围34.1~34.65),其营养盐浓度范围分别为NO3-N 1.5~13.7μmol/L(平均7.8μmol/L,n=14);PO4-P 0.37~1.53μmol/L(平均0.78 μmol/L,n=14);SiO3-Si 1.5~6.7μmol/L(平均3.7 μmol/L,n=14),对应深度叶绿素a的范围则在0.01~0.18μg/L之间(平均0.04μg/L,n=14)。此外,WCW与LW在春季与夏季开始与表层水SW混合,慢慢向IW改变[5],从而在TLW上30~50 m水深处形成了中层水(温度变化范围-0.5~1.0℃,盐度变化范围33.8~34.1),因其与Svendsen等提出的SW与TAW混合产生的IW在温盐上都有所不同[1],我们定义其为中层变异水(TIW),营养盐浓度平均值分别为NO3-N 1.6μmol/L;PO4-P 0.35μmol/L;SiO3-Si 2.0μmol/L。
在表层,随着夏季到来,冰川融水,雪融水和少量海冰融水直接或者间接通过径流进入湾内,使得表层水盐度降低,且由于极昼条件下持续的太阳辐射,导致表层水温度升高。有的情况下,冰川融水对王湾表层水盐度的影响可以到达距冰川前缘45 km的位置,且深度可以达30 m[17]。Wang等认为SW水能够影响到湾外侧水柱40 m以浅的海水[9]。调查区域内,30 m以浅盐度均小于34,且温度大于1℃,与前述SW温盐一致,划为表层水(SW,温度大于1℃,盐度小于34),其营养盐浓度范围NO3-N为0.7~2.3 μmol/L(平均1.2μmol/L,n=30);PO4-P为0.13~0.40μmol/L(平均0.24μmol/L,n=30);SiO3-Si为1.1~5.0μmol/L(平均2.5μmol/L,n=30),对应深度叶绿素a的范围则在0.07~1.24μg/L之间(平均0.64μg/L,n=30)。但此次在王湾的SW较之前王湾观测到的SW[1],温度(平均4.54℃)偏高,而盐度(平均31.9)低很多,说明2010年夏季王湾海域陆源和冰川输入将对王湾海域表层海水营养盐产生强烈影响。
在本地变异水以下,200 m以深的水团盐度与温度较高,符合大西洋水与北冰洋水的混合水团的温盐范围(温度大于1℃,盐度大于34.65),所以,该层即大西洋变异水(TAW)。研究发现,1991年夏季(7月),王湾口外典型大西洋水具有高温高盐,高营养盐的特征,NO3-N,PO4-P和SiO3-Si最大值分别为13.0 μmol/L,1.13μmol/L和6.0μmol/L;由陆架向湾内,营养盐明显降低(NO3-N,3.0~9.0μmol/L;SiO3-Si,2.0~3.0μmol/L;NH4-N,0.80~2.00μmol/L;PO4-P,0.15~0.35μmol/L)[11],说明,相较于大西洋水,大西洋变异水的营养盐浓度受到北极水影响而有所降低。
由于控制水团的不同,以上水团特征与Svendsen等的分类有所不同[1]。一般,夏末AW侵入范围达到最大[5];在某些年份,AW的影响范围可以到达整个王湾[18]。然而,大西洋温盐水和北极淡冷水相对强度的变化影响着它们在峡湾中的比例。2000年是王湾夏季大西洋来源水(AW)影响较弱的年份,在7月下旬,75~300 m水深处均由LW控制(温度小于1℃)[4]。类似的,王湾南部的Isfjord峡湾中,LW以及WCW甚至在夏末(2004年9月)仍存在[16],通过模型计算,Nilsen等认为这与冬季热通量及风向有关,它们决定了冬季湾内水体的密度,从而控制了峡湾系统与陆架区域的交换过程[16]。可见,2010年也是王湾受大西洋水影响较弱的年份,该年夏季的王湾水体主要受本地水与冬季残留水控制,水温较低,而且夏季水体层化明显(见图1),混合较弱,热交换相对较慢,有利于低水温层的保持,但该水层温度与范围还可能受其他因素影响,需进一步研究。
王湾水团的形成与其营养盐特征有着密不可分的联系,不同来源水的输入以及各水团的元素地球化学循环过程控制着王湾水体的营养盐分布特征。
本地水TLW以及中层变异水TIW主要来自于前一年夏末湾内水体,由于冬季温度降低,混合作用加强,其水体特征趋向均匀。2009年夏末,该水团中各营养盐含量随着深度增加而逐渐增高,其浓度分别为NO3-N 3.2~7.9μmol/L(平均5.9μmol/L,n=14);PO4-P 0.23~0.55μmol/L(平均0.44μmol/L,n=14);SiO3-Si 2.2~4.1μmol/L(平均3.2μmol/L,n=14)。2010年,TLW与TIW中,营养盐浓度总体比2009年夏末时高,而且大西洋高营养盐水与湾内交换也较弱,所以,本地水增加的营养盐值与前一年秋季至2010年夏季期间水体中颗粒物的营养盐再生有关。营养盐结构随着深度的增加也在逐渐的发生变化。本地水的氮磷比值与氮硅比值在深部分别增高接近16与2以上,可见,下层水体逐渐由潜在的氮限制转变为氮磷比较适宜与潜在的硅限制,此规律与2009年夏末该水体中,氮磷与氮硅比值的规律相似。该较高氮硅比值与北极欧亚侧大洋营养盐特征有所不同。亚欧北极区域受到陆源影响较强,硅营养盐在中下层水体中再生,浓度较高(可达10~60μmol/L)[19],而本区域常年受大西洋水进入影响,大西洋水氮/硅比值通常在2以上[11],从而,造成了王湾区域氮硅比值的高值。
底层200 m以深的大西洋变异水其营养盐来源主要是大西洋水与北极水的混合。由采样结果可以发现,接近200 m深处营养盐浓度平均值分别为NO3-N 13.5μmol/L;PO4-P 1.05μmol/L;SiO3-Si 6.1μmol/L,虽然该值与大西洋水的营养盐特征值接近[11],但其可能与大西洋水有一定差别。Hegseth与Tverberg在2006、2007与2008年4月与5月采集TAW水的硝酸盐与硅酸盐浓度范围分别为8.5~12.5μmol/L和4.5~6.5μmol/L[13]。Corte等在2008年6—7月也对王湾营养盐进行了分析,对应TAW的硝酸盐与磷酸盐浓度分别为6.87~8.28 μmol/L和0.66~0.75μmol/L[14]。可见,大西洋变异水的营养盐浓度相比于王湾外陆架大西洋水营养盐要低许多,说明大西洋水在进入湾内后与沿岸北极水混合,使得各营养盐浓度总体下降。根据文献值得到的王湾深部底层水(大于200 m)TAW的各营养盐水平分别约为NO3-N(9.0±3.0)μmol/L;PO4-P(0.70±0.10)μmol/L;SiO3-Si(5.5±1.0)μmol/L。以上值基本低于本地变异水的底层高值,这也间接说明TLW底部高值是与水体颗粒物营养盐的再生有关,并不能通过大西洋变异水的补给获得。
4.2 陆源输入对王湾表层水的影响
低营养盐的王湾夏季表层水可能来源有冰川融水,海冰融水,雪融水与地表径流等。王湾的海冰一般是在冬季形成,然后在春季快速消融,海冰中包含的卤水,颗粒物以及冰藻等对春季浮游植物的旺发有重要作用[12]。王湾还由于受到大西洋水入侵的影响,有时,海冰较早就开始消融,如2006年2月份,甚至2007年与2008年冬季湾内无冰[13]。同时,湾内靠近入海冰川处,会向海水延伸形成岸冰(含部分海冰),但王湾周围陆地75%被冰川覆盖,有5条冰川直接入海(这些入海冰川是在地面上的,而不是漂浮着的[1]),其他还有大量陆内冰川,因而,岸冰融水对表层水输入的贡献较为有限。Hagen等[20]对进入王湾系统淡水的年输入量进行了估算,发现淡水输入主要来自冰川崩解形成的融水,以及冰川与雪融水形成的径流,它们分别占到总径流量的18%和56%。因此,特别是在夏季,冰川崩解入海形成的融水以及冰雪融水形成的陆地径流是王湾表层水的主要来源。
在2010年夏季,对新奥尔松一侧陆地湖泊与河流进行了水样采集,并对其营养盐进行了分析(见表1)。根据测定结果可以发现,新奥尔松地表径流水中,湖水营养盐浓度总体较低,但其对王湾水体贡献很小,而河水与溪流水中硝酸盐与硅酸盐浓度相对较高,且该融水通量占淡水总输入量的一半以上,携带大量氮硅营养盐输入湾内。然而,王湾表层水SW营养盐浓度值较低,特别是硝酸盐和磷酸盐(最低值分别为0.7和0.13μmol/L),其平均硝酸盐和硅酸盐浓度均低于河水与溪流水值。所以,这部分陆源水进入湾内将影响王湾表层水营养盐的平衡。
除了通过地表径流输入湾内外,入海冰川会以崩解的冰块的形式直接入海,并在湾内融化,对王湾表层水产生重要影响。2012年采集的冰川水营养盐浓度为NO3-N 1.4~2.2μmol/L;PO4-P 0.01~0.03 μmol/L;SiO3-Si 0.5~1.4μmol/L(未发表数据),与表层水的营养盐浓度接近或更低。然而,这些低营养盐的入海冰川通量也较大,其对表层水的营养盐稀释作用也不可忽视。
由于王湾在夏季存在较为强烈的层化作用(见图1),通过以上径流方式输入湾内的淡水将主要停留在表层水中;并且,冰雪融水形成的径流营养盐稍高,也是主要的淡水输入通量,这些淡水输入形成的表层水营养盐浓度应该高于冰川崩解融水的营养盐浓度,而实际是与它们相近,这说明除了淡水输入外,还有其他因素使得表层水呈现出低氮磷营养盐的特征。
对采集的上游溪流水与河水分析发现,其叶绿素a浓度只有约0.10μg/L。它们进入湾内不足以支持王湾表层水的初级生产(叶绿素a浓度最大值为1.24 μg/L);而且,叶绿素a的高值与营养盐的低值是对应的(见图3),这说明浮游植物的生长在一定程度上发生在表层水中,且该过程消耗了部分营养盐,使得表层水氮磷营养盐浓度进一步降低。这与春季发生浮游植物旺发时类似,其表层叶绿素a可以到5.00μg/L以上,硝酸盐和硅酸盐浓度则降低到接近1.0μmol/L,甚至更低[13]。
4.3 生物活动对王湾营养盐分布的影响
海水中无机氮、磷和硅等营养物质是海洋生物繁殖生长不可缺少的化学成分。浮游植物的吸收与释放,浮游动物的摄食与排泄,以及它们的再生循环都会对营养盐的分布产生影响。
由上一节知道,在王湾表层水中,浮游植物对营养盐的吸收与王湾表层水中硝酸盐和磷酸盐的减少密切相关。然而,当营养盐浓度降到较低值时,营养盐也会限制浮游植物的生长。浮游植物生长所需溶解无机氮、硅酸盐和磷酸盐最低阈值分别为1.0 μmol/L、2.0μmol/L和0.1μmol/L[21]。实际观测发现王湾表层水仅溶解无机氮浓度基本在该阈值以下,所以,根据最小因子定律,表层水中,低浓度氮营养盐限制了浮游植物的生长。Redfield比值可以用来表示浮游植物生长潜在的限制性营养盐。王湾表层水中氮磷比值较低,基本低于5,而氮硅比值也较低,小于0.5(见图4),同样表明该水体潜在氮限制因素。
浮游植物的生长也对王湾表层水中硅酸盐的分布产生影响。由图3可以知道,表层硅酸盐浓度(约2.5μmol/L)高于次表层值(约1.5μmol/L)(见图3),而硅藻也是王湾的主要浮游植物种[17],说明硅藻为主的浮游植物在真光层的生物生产消耗了部分硅酸盐,而表1中看到的陆源水较高的硅酸盐输入(约大于5.0μmol/L)补充了表层消耗的硅酸盐,造成了该营养盐特征。
浮游动物的摄食与排泄等代谢过程中也可能对营养盐的平衡产生影响。由图3可以发现,2010年7月,王湾在100 m水深处出现一个铵盐高值区域(约50 m厚,大于2.0μmol/L),其形成原因可能与生物活动有关。下面对其形成原因进行探讨。一般,在缺氧条件下,当硝酸盐都已被还原时,铵盐将是化合态无机氮的主要形式[22],但从图3看,硝酸盐没有显著降低,说明铵盐的高值不是还原环境下产生的。水生动物的排泄也可能造成铵盐大量形成。根据拖网与过滤等现场调查发现,王湾海水水柱在夏季不同层位中,都存在大量大型浮游动物(水蚤类桡足动物为主)以及粪球粒(张武昌,个人通信)。前人研究也证实,夏季是王湾浮游动物丰度较高的时期(比春季高一个数量级[23])。Wiborg在1957年与1958年对王湾浮游动物进行了连续采样调查(靠近本次调查站位K1),也发现7、8月的浮游动物丰度是全年中最高的,且以飞马哲水蚤Calanus finmarchicus为主[24]。铵盐通常是浮游动物排泄物中主要的可溶性无机含氮化合物,而且浮游动物排泄氮的速率相对于其他动物来说高很多[22],所以,浮游动物的排泄物,如粪球粒,在下沉过程中的不断释放,可能是造成该区域铵盐高值的重要原因。此外,王湾夏季生产力一般较高,但藻类生物的现存量从叶绿素a的数据看较小(基本小于1.00μg/L),藻类生物量在夏季的减少可能与摄食作用有关[17]。王湾外陆架上曾发现的铵盐浓度高值及对应的桡足类生物量的增加,也被认为是浮游动物摄食减少了浮游植物生物量[11]。然而,Corte等在2008年6月到7月对王湾进行了连续4周的海水调查,发现在20 m到100 m深度铵盐浓度平均值从1.07μmol/L上升到2.30μmol/L,并以高浓度保持了近3周[14],所以,2010年夏季王湾的高铵盐区域也可能形成于春季或者更早。除了浮游动物的新陈代谢外,细菌利用动物排泄的粒状有机氮和其他悬浮在海水中的含氮碎屑物质作为能源物质,生成铵盐[22],这可能是该高铵盐浓度持续的重要原因,但它们的贡献量有待进一步研究。
5 结语
根据温度、盐度以及营养盐分布特征的不同,可以由上到下将2010年夏季王湾水体划分为表层水(SW)、中层变异水(TIW)、本地变异水(TLW)和大西洋变异水(TAW),发现,表层水与中层变异水营养盐浓度较低,随着深度增加,本地变异水与大西洋变异水营养盐浓度都较高。2010年夏季王湾是大西洋水影响较弱的年份,以低温低盐高营养盐的本地变异水为主导。大西洋变异水主要影响200 m以深水柱,且营养盐浓度低于本地变异水深部值,而本地变异水中营养盐浓度的增加主要来自于颗粒物中营养盐的再生。表层水主要来源于地表径流与冰川入海融水,这些淡水输入与浮游植物的初级生产共同构成了表层水的营养盐分布特征。本地变异水中,在水深100 m上下出现的铵盐高值区域可能与浮游动物和/或微生物的代谢过程有关。
王湾在营养盐生态动力学方面的研究有待进一步的深入,如表层水营养盐是否浮游植物生长的主要限制因子,以及长时间序列的连续观测能够促进对生态变化机制机理的研究。此外,扩大研究的时间与空间尺度,将有助于解释环境与气候变化对王湾生态系统产生的影响。
致谢:感谢2010年北极黄河站度夏考察所有队员的帮助与支持;同时感谢国家海洋局极地考察办公室对现场考察以及研究项目的资助。
参考文献:
[1] Svendsen H,Beszczynska-Møller A,Hagen J O,et al.The physical environment of Kongsfjorden-Krossfjorden,an Arctic fjord system in Svalbard[J].Polar Research,2002,21(1):133-166.
[2] Tverberg V,Nøst O A.Eddy overturning across a shelf edge front:Kongsfjorden,west Spitsbergen[J].Journal of Geophysical Research,2009,114,C04024,doi:10.1029/2008JC005106.
[3] Hop H,Pearson T,Hegseth E N,et al.The marine ecosystem of Kongsfjorden,Svalbard[J].Polar Research,2002,21(1):167-208.
[4] Hop H,Falk-Petersen S,Svendsen H,et al.Physical and biological characteristics of the pelagic system across Fram Strait to Kongsfjorden[J].Progress in Oceanography,2006,71(2/4):182-231.
[5] Cottier F,Tverberg V,Inall M E,et al.Water mass modification in an Arctic fjord through cross-shelf exchange:The seasonal hydrography of Kongsfjorden,Svalbard[J].Journal of Geophysical Research,2005,110,C12005,doi:10.1029/2004JC002757.
[6] Cottier F R,Nilsen F,Inall M E,et al.Wintertime warming of an Arctic shelf in response to large scale atmospheric circulation[J].Geophysical Research Letters,2007,34,L10607,doi:10.1029/2007GL029948.
[7] Nilsen F,Gjevik B,Schauer U.Cooling of the West Spitsbergen Current:Isopycnal diffusion by topographic vorticity waves[J].Journal of Geophysical Research,2006,111,C08012,doi:10.1029/2005 JC002991.
[8] Cui S K,He J F,He P M.The adaptation of Arctic phytoplankton to low light and salinity in Kongsfjorden(Spitsbergen)[J].Oceanography,2006,71:182-231.
[9] Wang G,Guo C,Luo W,et al.The distribution of picoplankton and nanoplankton in Kongsfjorden,Svalbard during late summer 2006[J].Polar biology,2009,32(8):1233-1238.
[10] Guo C Y,He J F,Zhang F,et al.Spatial heterogeneity of a microbial community in Kongsfjorden,Svalbard during late summer 2006 and its relationship to biotic and abiotic factors[J].Advance in Polar Science,2011,22:55-66.
[11] Owrid G,Socal G,Civitarese G,et al.Spatial variability of phytoplankton,nutrients and new production estimates in the waters around Svalbard[J].Polar Research,2000,19(2):155-171.
[12] Hodal H,Falk-Petersen S,Hop H,et al.Spring bloom dynamics in Kongsfjorden,Svalbard:nutrients,phytoplankton,protozoans and primary production[J].Polar Biology,2012,35(2):191-203.
[13] Hegseth E N,Tverberg V.Effect of Atlantic water inflow on timing of the phytoplankton spring bloom in a high Arctic fjord(Kongsfjorden,Svalbard)[J].Journal of Marine Systems,2013,113/114:94-105.
[14] Corte D D,Sintes E,Yokokawa T,et al.Changes in viral and bacterial communities during theice-melting season in the coastal Arctic(Kongsfjorden,Ny-Ålesund)[J].Environmental Microbiology,2011,13(7):1827-1841.
[15] Willis K,Cottier F,Kwasniewski S,et al.Theinfluence of advection on zooplankton community composition in an Arctic fjord(Kongsfjorden,Svalbard)[J].Journal of Marine Systems,2006,61(1/2):39-54.
[16] Nilsen F,Cottier F,Skogseth R,et al.Fjord-shelf exchanges controlled by ice and brine production:Theinterannual variation of Atlantic Water in Isfjorden,Svalbard[J].Continental Shelf Research,2008,28(14):1838-1853.
[17] Keck A,Wiktor J,Hapter R,et al.Phytoplankton assemblages related to physical gradients in an Arctic,glacier-fed fjord in summer[J].ICES Journal of Marine Science,1999,56(Suppl.):203-214.
[18] MacLachlan SE,Cottier F R,Austin W E N,et al.The salinity:δ18O water relationship in Kongsfjorden,western Spitsbergen[J].Polar Research,2007,26(2):160-167.
[19] 李宏亮,陈建芳,刘子琳,等.北极楚科奇海和加拿大海盆南部颗粒生物硅的粒级结构[J].自然科学进展,2007,17(1):72-78.
[20] Hagen J O,Kohler J,Melvold K,et al.Glaciers in Svalbard:mass balance,runoff and freshwater flux[J].Polar Research,2003,22(2):145-159.
[21] Justic D,Rabalais N N,Turner R E.Stoichiometry nutrient balance and origin of coastal eutrophication[J].Marine Pollution Bulletin,1995,30:41-46.
[22] Riley J P,Skirrow G.Chemical oceanography[M].2nd ed.London:Academic Press,1975:297-316.
[23] Walkusz W,Kwasniewski S,Falk-Petersen S,et al.Seasonal and spatial changes in the zooplankton community of Kongsfjorden,Svalbard[J].Polar Research,2009,28(2):254-281.
[24] Wiborg K F.Investigations on Zooplankton in Norwegian Waters and in the Norwegian Sea during 1957-58[R]//Report on Norwegian Fishery and Marine Investigations.Bergen:A.S John Griegs Boktrykkeri,1960,12(6):1-19.
Nutrient distribution and the influencing factors of seawater in Arctic Kongsfjorden,summer 2010
Ji Zhongqiang1,Gao Shengquan1,Jin Haiyan1,2,He Jianfeng3,Bai Youcheng1,Wang Bin1,Yang Zhi1,Chen Jianfang1,2
(1.State Oceanic Administration Laboratory of Marine Ecosystem and Biogeochemistry,Second Institute of Oceanography,State O-ceanic Administration,Hangzhou 310012,China;2.State Key Laboratory of Satellite Ocean Environment Dynamics,Hangzhou 310012,China;3.Polar Research Institute of China,Shanghai 200136,China)
Kongsfjorden is located in the northwest of Svalbard,Arctic,and the water is affected by Arctic glacier and the Atlantic Ocean.In order to study the affects of glacier water and Atlantic water on distribution of nutrients and the ecosystem in Kongsfjorden,seawater and runoff water were sampled and analyzed for the nutrients and Chlorophyll a during the Yellow River Station expedition in summer 2010.The results showed that,it was a weak year of the Atlantic water in kongsfjorden 2010,and the transformed local water with low temperature,low salinity and high nutrients was in control.The Kongsfjorden water in July 2010 could be divided into 4 water masses,including surface water(SW),transformed intermediate water(TIW),transformed local water(TLW)and transformed Atlantic water(TAW),of which,the nutrient concentration were low in SW and TIW,while,as the depth went deeper,nutrient concentrations were higher in TLWand TAW.The water column below 200 m was mainly controlled by TAW,and its nutrient concentration was lower than those in the bottom of TLW.The higher nutrients of TLW were mainly derived from the nutrient regeneration of the particulates in the water column.The glacier melting water and runoff water controlled the SW,and these freshwater inputs and the primary production of the phytoplankton together determined the nutrient concentrations and structures in SW.In the water column at about 100 m of TLW,the high ammonia concentration was connected with the metabolic processes of the zooplankton and/or microorganism.
Arctic;Kongsfjorden;nutrients;water mass
P734.2+1
A
0253-4193(2014)10-0080-10
2014-04-10;
2014-07-15。
极地专项项目(CHINARE2013-02-01,CHINARE2013-04-03-07);国家海洋局基金项目(2013331,2011327);海洋公益性行业科研专项经费项目(201105022);中国极地科学战略研究基金项目(20120104,20120305)。
季仲强,男(1984—),博士,助理研究员,主要从事海洋生物与有机地球化学研究。E-mail:jizq@sio.org.cn
金海燕,研究员,主要从事海洋生物地球化学研究。E-mail:jinhaiyan@sio.org.cn
季仲强,高生泉,金海燕,等.北极王湾2010年夏季水体营养盐分布及影响因素[J].海洋学报,2014,36(10):80—89,
10.3969/j.issn.0253-4193.2014.10.009
Ji Zhongqiang,Gao Shengquan,Jin Haiyan,et al.Nutrient distribution and the influencing factors of seawater in Arctic Kongsfjorden,summer 2010[J].Acta Oceanologica Sinica(in Chinese),2014,36(10):80—89,doi:10.3969/j.issn.0253-4193.2014.10.009
猜你喜欢
草业学报(2022年5期)2022-07-08
草业学报(2021年3期)2021-03-22
草业学报(2018年10期)2018-10-19
草业科学(2018年7期)2018-07-26
小哥白尼(野生动物)(2018年3期)2018-06-15
小哥白尼(趣味科学)(2018年2期)2018-05-25
学生天地(2017年11期)2017-05-17
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
