
长江口及其邻近陆架区夏季网采浮游植物及其影响因素
2014-06-01 12:30:09江志兵曾江宁李宏亮周锋陈全震朱根海张东声严小军
海洋学报 2014年6期
江志兵,曾江宁,李宏亮,周锋,陈全震,朱根海,张东声,严小军
(1.宁波大学海洋学院应用海洋生物技术教育部重点实验室,浙江宁波 315211;2.国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州 310012;3.国家海洋局第二海洋研究所卫星海洋环境动力学国家重点实验室,浙江杭州 310012)
长江口及其邻近陆架区夏季网采浮游植物及其影响因素
江志兵1,2,曾江宁2,李宏亮2,周锋3,陈全震2,朱根海2,张东声2,严小军1*
(1.宁波大学海洋学院应用海洋生物技术教育部重点实验室,浙江宁波 315211;2.国家海洋局第二海洋研究所国家海洋局海洋生态系统与生物地球化学重点实验室,浙江杭州 310012;3.国家海洋局第二海洋研究所卫星海洋环境动力学国家重点实验室,浙江杭州 310012)
2009年6、8月长江口及其邻近陆架区的网采样品中检出浮游植物9门395种(含223种硅藻与125种甲藻)。浮游植物丰度8月(3 077.15×104cells/m3)显著高于6月(107.80×104cells/m3)。随长江冲淡水势力增强,位于长江口的丰度高值区8月较6月更偏外侧。种类丰富度8月高于6月,但多样性和均匀度指数略低于6月。6月尖刺伪菱形藻和三角角藻占绝对优势,8月优势种主要有尖刺伪菱形藻、笔尖形根管藻和铁氏束毛藻。骨条藻虽非优势种,但在长江冲淡水区丰度较高。相似性分析和多维尺度分析表明,浮游植物群落组成时空差异显著。典范对应分析表明,温度和盐度是分别影响6、8月群落分布的首要因子。根据水动力和化学参数,该区浮游植物群落分布与环流变化和水团消长密切相关。
东海;长江口;浮游植物;时空分布;环境因子
1 引言
海洋浮游植物作为最主要的初级生产者,可为浮游动物等消费者提供食物和必需养分,但其引发的有害水华也会影响近海生态系统的结构、功能、服务和产出[1]。随着长江流域人口的快速增长和经济的迅猛发展,氮磷营养盐入海通量剧增[2—5],导致长江口及其邻近水域富营养化加剧、有害藻华频发[3—4,6]。因此,该海区浮游植物的群落特征及其影响因素的研究备受学术界关注,历年来大范围的调查[7—10]也较多。但这些调查均在三峡水库蓄水(2003年)前进行,而在蓄水后,因径流量、入海泥沙、营养盐和水动力等变化的影响[11—12],浮游植物的群落组成和结构可能已发生改变。另外,近来该区夏季浮游植物网样[13—16]和水样[17—20]的研究也存在空间(集中于29°~32°N,123°E以西的长江口海域,研究范围有限)与时间(仅有一个航次)尺度上的局限性。夏季不同月份长江口及其邻近海域的水文状况存在明显的时空变化,尤其是长江冲淡水的转向与扩展及西南季风盛行下台湾暖流的入侵[21—24]。因此,以往对夏季东海浮游植物分布的变化及其影响因素的认识可能存在不足。
2009年6、8月拖网采集了长江口及其邻近陆架区(27.75°~33.25°N,122.0°~125.0°E)的浮游植物,研究其分布特征和影响因素。本次调查范围更广,从长江口外延伸至其邻近的东海内陆架,同时选择在夏季的不同月份实施两次调查来探讨浮游植物与环境因子的耦合关系及浮游植物的演替状况,弥补了以往调查在时空尺度上的欠缺。本文还对比了近30年来该区浮游植物的相关资料,结合理化环境因素,分析了网采浮游植物的长期变化。研究结果可更好地了解该区浮游植物的生态现状和群落演替趋势及其对环境变化的响应,为其它相关学科的研究提供参考。
2 材料与方法
2.1 调查区域与站位
2009年6月1日至11日和8月14日至24日,随“北斗”号调查船对长江口及其邻近陆架区进行了2个航次调查(图1)。该海域夏季主要受长江冲淡水、苏北沿岸流、浙闽沿岸流和台湾暖流等环流控制,调查海域东北部及东南部外侧海域分别还受黄海冷水团和黑潮的影响[22,24]。

图1 长江口及其邻近陆架区的调查断面和站位设置及其夏季环流
2.2 样品采集及分析
在每站用孔径为76μm、网口附Hydro-Bios流量计的浮游生物网垂直拖网采集浮游植物。样品用4%中性甲醛固定,回实验室经浓缩后用Leica DM2500显微镜观察和计数[25]。用CTD测定温度、盐度和水深。用10 L采水器采集表层(2 m)水样,测定溶解氧(Dissolved oxygen,DO)、溶解无机氮(DIN:++)、活性磷酸盐(DIP)、硅酸盐(DSi)和悬浮物浓度。
2.3 数据分析
用PRIMER 5.0版软件计算浮游植物群落的Shannon-Weiner多样性指数(H′)和Pielou均匀度指数(J′)。优势度(Y)计算公式为Y=ni·fi/N× 100%,式中:N为样品的总丰度;ni为样品中第i种的丰度;fi为该种浮游植物在样品中的出现频率。当Y≥2%则认定为优势种。用SPSS13.0软件对相关数据进行统计检验。用PRIMER软件对物种丰度进行log(x+1)转换后,建立Bray-Curtis相似性聚类分析,并采用非度量多维尺度分析(non-Metric multidimensional scaling,n MDS)分析浮游植物群落的空间分布。浮游植物群落分布的时空比较采用单因素相似性分析(Analysis of similarity,ANOSIM)。选取优势种,用CANOCO 4.5软件进行CCA排序,物种丰度和环境参数(p H除外)均经log(x+1)转换。
3 结果与分析
3.1 群落组成
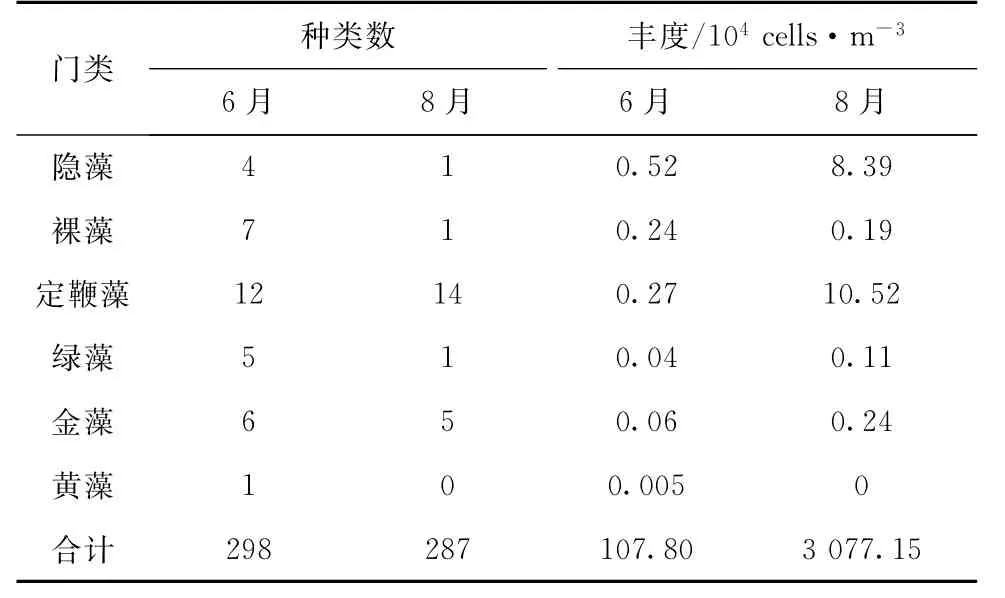
2个航次共鉴定出浮游植物9门395种(含变种和变型及未定种)。其中硅藻和125种甲藻。尽管6、8月总物种数相差不大,但硅藻、蓝藻、甲藻、隐藻和定鞭藻丰度月份差异较大(表1),且前两者在6、8月间均差异显著(p<0.05)。6、8月甲藻物种组成类似,均以角藻Ceratium和原多甲藻Protoperidinium居多。6、8月浮游植物总丰度分别为1.64×104~972.63×104和57.40×104~3 4201.85×104cells/m3,8月平均丰度显著(p<0.05)高于6月(表1)。无论是种类组成还是细胞数量组成,6、8月浮游植物均以硅藻占主导优势。

表1 6、8月浮游植物的种类数和丰度
续表1
3.2 丰度
硅藻、甲藻和浮游植物丰度分布在6、8月间存在明显差异(图2)。6月总丰度高值区位于长江口海域,次高值区位于长江口外海域(图2a);8月总丰度高值区位于长江口海域,较6月东移,次高值区位于浙江中部近海(图2b)。6、8月硅藻丰度高值区均对应于总丰度高值区(图2c和2d)。6月甲藻丰度高值区在长江口外呈哑铃形分布(图2e),8月位于长江口北侧海域(32.5°N,122.5°E)、长江口(31°N,123.5°E)和浙江中部近海(29°N,122.5°E)(图2f)。

图2 6月(a、c和e)、8月(b、d和f)浮游植物、硅藻和甲藻丰度(104cells/m3)的分布
从垂直岸线的断面分布看,6月B、C断面丰度远高于其余断面(见图3a),而8月北部断面(A、B和C)丰度高于南部断面(D、E和F)(见图3b),2个航次长江口主断面(C)丰度均明显高于其余断面。从平行于岸线的断面分布看,调查海域中部断面高于近岸和离岸断面(见图3),6月Ⅱ断面丰度最高,而8月则Ⅲ断面丰度最高,较6月东移。
3.3 优势种
6、8月浮游植物优势种组成和平面分布存在明显差异(见表2和图4)。6月尖刺伪菱形藻Pseudo-nitzschia pungens(Y=22.2%)和三角角藻Ceratium tripos(Y=16.1%)占绝对优势,其丰度分别为32.67×104和18.60 ×104cells/m3;8月除铁氏束毛藻Trichodesmium thiebautii为蓝藻外,其余优势种均为硅藻,包括尖刺伪菱形藻和笔尖形根管藻Rhizosoleniastyliformis等。

图3 6月(a)、8月(b)浮游植物丰度(Mean±SE)在不同断面的分布

表2 6、8月网采浮游植物优势种的丰度(104cells/m3)和优势度(Y,%)

图4 6月(a、c、e和g)、8月(b、d、f和h)浮游植物主要优势种和骨条藻的丰度(104cells/m3)分布
6月:三角角藻密集区呈哑铃形分布于长江口外(见图4b);具槽帕拉藻Paralia sulcata和铁氏束毛藻密集区分别分布于东海外侧和东南外海外,其余优势种高值区均位于近岸(或偏近岸),特别是在长江口附近海域,如尖刺伪菱形藻(见图4a)和蛇目圆筛藻Coscinodiscus argus(见图4c)。8月:除铁氏束毛藻外(见图4f),其余优势种高值区位于调查海域北部和西部,尤其在长江口外和浙江中部近海,如尖刺伪菱形藻(见图4b)和笔尖形根管藻(见图4d)。骨条藻S犽eletonema spp.虽然不是调查海域的优势种,但其在长江口海域的丰度较高,分布与冲淡水主体高度吻合。6、8月骨条藻的平均丰度分别为7.34×104cells/m3和56.51×104cells/m3,6月高值区位于长江口以北海域(见图4g),8月高值区位于长江口和浙江中部近海(见图4h)。
3.4 种类数、多样性和均匀度
6、8月浮游植物的种类数(S)、H′和J′之间无显著差异。8月S(52.1)高于6月(46.8),而H′和J′(分别为2.08和0.51)略低于6月(分别为2.12和0.56)。S高值区6月位于东南侧外海(图5a),8月位于浙江中部近海和东北侧外海(图5b);H′和J′高值区6月位于调查海域北部和浙江中部近海(图5c和5e),8月位于调查海域中部和浙江中部近海(图5d 和5f)。

图5 6月(a、c和e)、8月(b、d和f)浮游植物种类数、多样性指数和均匀度指数分布
3.5 群落分布差异
根据ANOSIM,6、8月浮游植物群落组成差异显著(p=0.001,R=0.48)。6月北部A、B和C断面与南部E和F断面群落组成差异显著(p<0.01),8月仅A和B断面与F断面群落组成差异显著(p<0.05);6月近岸Ⅰ、Ⅱ和Ⅲ断面与离岸Ⅳ和Ⅴ断面差异显著(p<0.05),8月Ⅰ和Ⅱ断面与Ⅴ断面有显著(p< 0.05)差异。聚类分析和n MDS表明,浮游植物群落均可分为3组(见图6)。其中:组a和d基本为河口半咸水类群和近岸低盐类群(代表种有骨条藻、尖刺伪菱形藻和夜光藻Noctiluca scintillans等),主要受长江冲淡水和沿岸水影响;组b主要为海洋广布类群(如具槽帕拉藻和三角角藻等),受陆架混合水和黄海冷水团影响;组c和f为外海高盐类群(如铁氏束毛藻),主要受台湾暖流水与黑潮次表层水的影响;组e为近岸低盐的暖水类群(如洛氏角毛藻Chaetoceros lorenzianus和笔尖形根管藻等),主要受台湾暖流和长江冲淡水的共同影响,且位于冲淡水锋面附近。
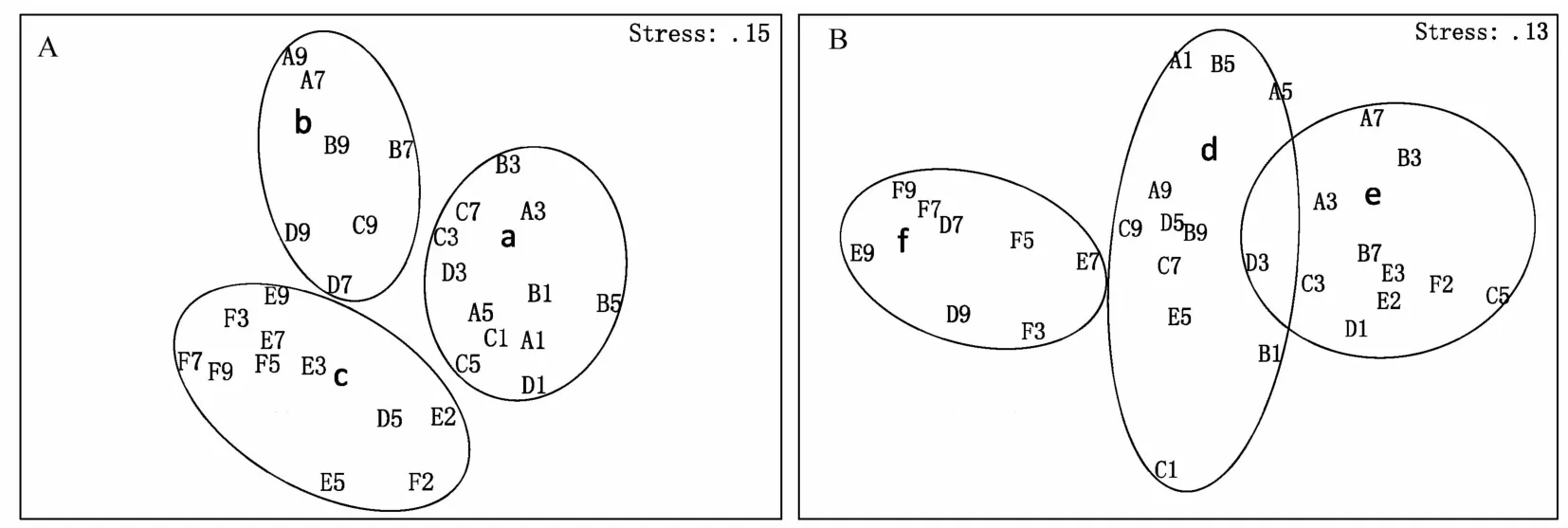
图6 基于聚类和多维尺度分析的6(A)、8月(B)浮游植物群落空间分布
3.6 环境因子
6月在浙江海域外侧受台湾暖流外侧分支影响,有一支盐度超过34的暖水舌(温度超过24℃)向北扩展,同时在调查区东北部有一支向南的冷水舌分布,最低温度小于18℃,表层呈现为闭合等值线分布(图7a)。8月,南方的暖水舌(台湾暖流)势力明显增强(图7b),使暖水舌北上到达31°N附近。调查区东北侧仍盘踞着陆架混合水,水温较低,其底层水温仅12℃,说明该区下层水体仍受黄海冷水团的影响。8月温度(28.1℃)显著(p<0.001)高于6月(21.4℃)。6月冲淡水的势力还较弱,舌形较宽,羽状锋有向东北扩展的趋势(见图7c)。8月冲淡水的势力增强,舌形较窄,向外海延伸(图7d),且低盐(≤31)水舌主要是向东(越过124°E)、向南扩展(至28.5°N)。尽管8月冲淡水势力强于6月,但表层平均盐度8月(31.0)高于6月(30.5),说明8月东海陆架受台湾暖流影响增大。
6、8月DIN分布与低盐水舌分布类似(图8a和8b)。DIN、DIP和DSi高值区均位于长江口附近海域(见图8),但8月DIP有一明显次高值区,正中心位于30°N,123°~123.5°E。8月平均DIN与DIP(分别为14.62和0.30μmol/L)高于6月(分别为13.41和0.25μmol/L),但DSi则表现为6月(14.78μmol/L)高于8月(10.31μmol/L)。
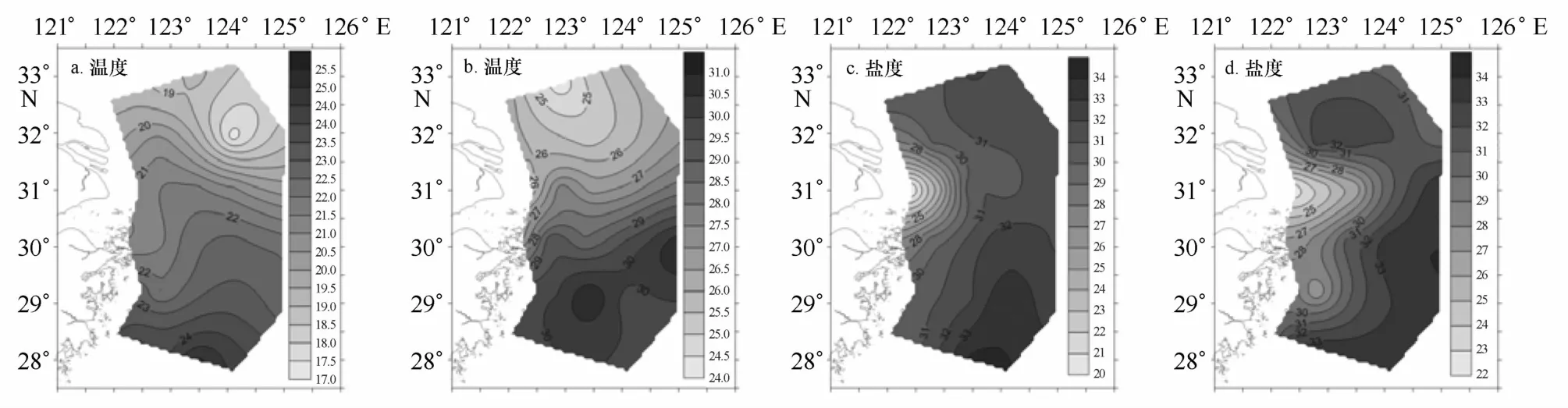
图7 6月(a、c)、8月(b、d)表层温度(℃)和盐度的分布
3.7 典范对应分析(CCA)
蒙特卡罗检验表明,6、8月CCA中的第1轴和全部轴均为显著(p<0.01),故排序结果是可信的。图9表明,6月的温度、营养盐(DIN、DSi和N/P)和水深以及8月的盐度、营养盐和水深是依次影响各自月份浮游植物群落的主要环境因子,且营养盐浓度随盐度和水深的增加而明显降低。骨条藻、尖刺伪菱形藻、蛇目圆筛藻、扁面角毛藻Chaetoceros compressus、洛氏角毛藻、三角藻和夜光藻等与营养盐密切相关,而与盐度负相关,表明这些种类适宜长江冲淡水低盐、营养盐丰富的环境;铁氏束毛藻与温度、盐度和水深正相关,而与营养盐和悬浮物浓度负相关,表明该种在外洋暖水(台湾暖流)中占优势;伏氏海毛藻Thalassiothrix frauenfeldii、笔尖形根管藻和伪菱形藻与营养盐和盐度无明显相关,而与温度负相关,表明这些种类在冲淡水锋面附近(盐度和营养盐适中)相对低温的环境中占优势;图9a中心的柔弱伪菱形藻属相对广温广盐种,说明其对环境适应性较好,且在东海的分布也较广。
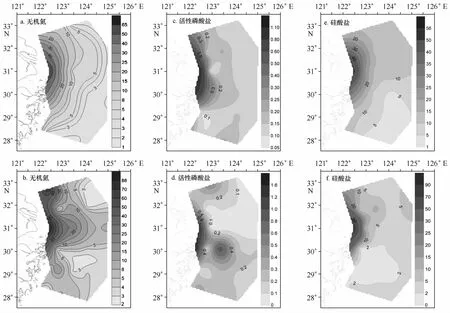
图8 6月(a、c和e)、8月(b、d和f)表层营养盐(μmol/L)的分布
4 讨论
4.1 浮游植物与环境因子的关系
研究海域夏季主要受长江冲淡水、沿岸流、台湾暖流、黑潮次表层水和黄海冷水团等影响(见图1),浮游植物种类组成丰富(见表1),共395种,包括淡水种(如颤藻Oscillatoria和裸藻)、河口半咸水种(如克尼角毛藻Chaetoceros犽nipowitschi)、沿岸低盐种(如骨条藻和夜光藻等)和外海高盐种(如鸟尾藻Ornithocercus和束毛藻等)。受上述水团和环流影响[22,24],夏季浮游植物呈现高度的空间异质性[7—9]。根据聚类分析和nMDS,6、8月浮游植物均可分三大类(见图6),即受控于低盐的长江冲淡水与沿岸水以及高盐的台湾暖流与黑潮次表层水,故ANOSIM显示该区夏季浮游植物群落组成和分布具有显著的时空差异。随台湾暖流向北和向近岸入侵加剧,8月南、北断面之间的差异较6月小,离岸、近岸断面亦是如此。
6月西南季风盛行,自南向北的台湾暖流和浙闽沿岸流带来了丰富的暖水种,使得南部海域的S、H′和J′均明显高于受控于长江冲淡水、苏北沿岸流和黄海冷水团的北部海域(见图5)。尤其是在调查海域东南部,受台湾暖流外侧分支影响,S最高,这与郭玉洁等[7]调查结果相符。因束毛藻占绝对优势,该区H′和J′相对较低。8月随台湾暖流入侵加剧,在长江口附近被上层长江冲淡水阻挡,其锋面(以盐度31为界)外侧S、H′和J′均较高(见图5)。尽管8月S高于6月,但因8月浮游植物个别优势种大量繁殖,降低了群落的多样性和均匀度,导致8月H′和J′均低于6月。

图9 6月(a)、8月(b)浮游植物群落与环境因子间的CCA排序(物种代码见表2;SS:悬浮物)
夏季,长江冲淡水在向东北扩展时,在近口门处分向东南、东北两侧[21—22]。2股的相对强弱受径流量影响,径流量大,向东南扩展的一股强,反之则朝东北扩展的一股强[23]。本文6、8月长江冲淡水的扩展也是如此(见图7)。6月,长江冲淡水出口门后指向东北一侧(见图7c),并在123.5°E附近形成锋面,使得优势种尖刺伪菱形藻、蛇目圆筛藻和骨条藻等大量繁殖(见图4)。而甲藻(主要为三角角藻)高值区(图2e)紧邻硅藻高值区(见图2c)。在浙江外侧海域,海表温度和盐度分别超过24℃和34(见图7),说明该处受台湾暖流外侧分支影响较强[22,24],适应高温高盐、低营养的束毛藻丰度较高。8月,冲淡水势力增强(大通站同期月均径流量由6月的36 000 m3/s增至8月的41 000 m3/s),舌形分向东北、东南两侧(见图7d),并在长江口以东的124°E以及浙江中部近海的29°N附近形成锋面(见图2),导致洛氏角毛藻、扁面角毛藻、尖刺伪菱形藻和骨条藻等硅藻在这两处海域大量繁殖(见图2和图4)。这与以往调查资料基本相符[7—10]。因此,长江口和浙江近海也是夏季东海赤潮的高发区[3,6]。台湾暖流在表层被长江冲淡水阻挡[21—22],该水文特征与优势种的分布密切相关。譬如铁氏束毛藻低值区刚好对应于长江冲淡水的低盐水舌,而笔尖形根管藻和伏氏海毛藻等外洋广布种高值区则刚好位于低盐水舌两侧(见图7d)。浮游植物丰度高值区与长江冲淡水、沿岸流和台湾暖流锋面重合,形成了高度的多样性。
6、8月营养盐高值区均位于长江口附近,随盐度升高而向外海逐渐降低,且DIN、DIP和DSi分布与长江冲淡水(低盐水舌)的分布和转向密切相关(见图8)。CCA排序也表明营养盐浓度与盐度呈显著负相关(图9),这符合该区的营养盐分布规律[4,26]。随长江冲淡水势力增强,8月DIN与DIP明显高于6月,但DSi反之,这或许与8月硅藻大量繁殖消耗掉有关。张传松等[26]观察到东海春、夏季硅藻水华时DSi浓度快速下降。8月DIP有一明显次高值区,正中心位于30°N,123°~123.5°E,这可能与浙江近岸上升流带来丰富DIP有关。根据文献[21—22]报道,夏季在浙江近海28°~31°N,124°E以西的狭长区块有明显的上升流,呈低温、高盐高磷特征。由于底层台湾暖流水涌升,大量暖水种被带至上层,使该狭长海域出现高S、H′和J′的现象(见图5)。因太阳辐照和冲淡水势力增强,8月水温和营养盐较6月明显升高,导致8月浮游植物丰度显著高于6月(见表1)。长江口主断面受长江冲淡水控制,其丰度远高于其余断面。而调查海域中部断面刚好位于锋面附近,其丰度高于近岸和离岸一侧(见图3)。6月Ⅱ断面丰度最高,而8月Ⅲ断面丰度最高。这表明长江冲淡水势力增强使得锋面东移,导致浮游植物丰度高值区也随之向东移动(见图2)。
长江口及其邻近陆架区的浮游植物群落受到水动力、光照、温度、盐度和营养盐等环境因子[3,11,19]的多重影响。CCA结果显示,6月的温度、营养盐和盐度以及8月的营养盐、盐度和水深是依次影响各自月份浮游植物群落的主要环境因子。CAA排序较好的展示了浮游植物空间分布与环境因子的密切联系(图9)。例如:嗜营养的低盐种骨条藻与营养盐密切正相关,而与盐度负相关,高值区均位于长江冲淡水主体内(见图4h和图4g);高温高盐种的束毛藻与温度、盐度和水深密切正相关,而与营养盐和悬浮物密切负相关,低值区与冲淡水低盐水舌高度吻合(见图4f和图7d)。6、8月硅藻在群落中占绝对优势,其丰度高值区与藻类总丰度高值区相一致,均位于冲淡水的锋面附近(见图2)。锋面邻近海域营养盐丰富、透明度相对较高、温度和盐度适宜(见图7和图8),适合浮游植物生长和繁殖。甲藻优势种三角角藻高值区则位于硅藻高值区外侧,这与2003年6月异帽藻Heterocapsa在锋面外侧丰度较高类似[23]。
4.2 浮游植物群落演变及其环境指示作用
表3列出了近30年来长江口及其邻近陆架区夏季网采浮游植物丰度和主要优势种组成。因调查区域、时间差异以及个别年份骨条藻大量繁殖,导致长江口浮游植物丰度变异性较大,但总体而言,近年来该区夏季网采丰度呈上升趋势。优势种虽仍主要由链状硅藻(包括骨条藻、伪菱形藻和角毛藻等)组成,但本次调查发现大、中型甲藻(三角角藻和夜光藻)和蓝藻(束毛藻)也成为该海域的优势种,而骨条藻优势度有所下降。

表3 夏季长江口及其邻近陆架区网采浮游植物的平均丰度(104cells/m3)和主要优势种
近年来水样分析表明,硅藻和小型甲藻在本调查海域占共同优势[17-20,23]。这与网采结果并不一致,原因是拖网采集浮游植物通常会漏掉部分个体小于76μm的物种,特别是一些营单个细胞生活的小型甲藻。故在今后的工作中最好同时采集网样和水样进行分析。近50年来长江口浮游植物群落的物种组成、丰度及优势种发生了很大变化,赤潮发生频率剧增,且大规模藻华优势种也呈从硅藻到硅藻与甲藻(东海原甲藻Prorocentrumdonghaiense和米氏凯伦藻Karenia mi犽imoto)并存的演变趋势[2—5]。东海近海有害藻华的发生亦是如此[2,6]。
长江口及其邻近陆架区浮游植物群落的异常演替与营养浓度上升和结构(N/P和Si/N)改变有关[2-5]。Zhou等[3]研究表明:长江口口门处硝酸盐已从1960s的20.5μmol/L升至1980s的59.1μmol/L,到1990-2004年其浓度已达80.6μmol/L;DIP变化相对较小,其浓度仅由1980s的0.59μmol/L升至1990-2004年的0.77μmol/L。而Li等[2]根据大通站资料发现,因水库数量增加,长江入海DSi浓度和通量自1950s以来急剧下降。同样,余立华等[29]研究表明,长江口海域夏季DSi浓度由1959年的42.12μmol/L降至1999年的20.98μmol/L,到三峡水库蓄水后,进一步降至14.47μmol/L,导致N/P和N/Si不断升高。在这种营养状况下,甲藻等非硅质微藻较硅藻竞争优势明显[30]。特别是长江截留后,氮磷入海通量持续升高[4],硅酸盐通量和入海泥沙量却急剧下降[12],导致长江口及其邻近陆架区富营养化加剧,且浊度有所降低,这也有利于浮游植物旺发和有害水华持续爆发[11,31—32]。
近年来长江口及其邻近陆架区浮游植物群落中除甲藻优势度升高外,外海暖水种(如细长翼鼻状藻Proboscia alata f.Gracillima、束毛藻和笔尖形根管藻等)优势度也明显升高(见表3)。这种演变趋势可能与外海水入侵加剧和海水温升有关[32]。自2000后,长江径流量呈下降趋势[4,32],导致外海水的入侵加剧[31—32],从而带来更多的外海种。另外,受全球气候变暖影响,东海海表温度在1945—2006年间已升高了0.9℃[33],而长江口邻近海域8月海表温度在1982—2009年间更是升高了约1.85℃[32],温度的升高有利于暖水种的生长。因此,外海水入侵和海水温升也可能使该区浮游植物分布格局发生改变。在全球气候变化的背景下,异常天气增多和气候波动可通过海、气影响近海环流路径、水体层化强度和冲淡水锋面分布,从而使物理环境出现明显波动[34],而浮游植物恰恰对这种扰动尤为敏感,故近年来海洋环境演变也可能引起该区浮游植物群落改变。
5 结论
长江口及其邻近陆架区受长江冲淡水、台湾暖流、沿岸流、黄海冷水团和黑潮次表层水等影响,物种组成丰富,多样性较高。6、8月网采浮游植物群落以硅藻为主,甲藻和蓝藻丰度也较高。从空间分布来看,在富营养盐的近岸主要由硅藻和甲藻组成,而在寡营养的远岸则为蓝藻占优势。浮游植物高值区位于长江口外侧和浙江中部近海,且与长江冲淡水主体位置高度吻合。优势种主要有尖刺伪菱形藻、笔尖形根管藻、三角角藻和铁氏束毛藻等。6、8月的首要影响因素分别为温度和盐度,营养盐和水深均为次要因素。该海域浮游植物群落组成与分布受控于环流变化和水团消长,尤其是长江冲淡水和台湾暖流。近年来该区夏季网采浮游植物数量呈明显上升趋势,虽然优势种仍主要由链状硅藻组成,但甲藻和蓝藻优势度有所升高;外海暖水种/广布种也逐渐在长江口占据优势,与外海水入侵及海水温升密切关联。浮游植物的异常演替可较好指示长江口及其邻近海域生态环境的演变。
致谢:感谢“北斗”号全体船员在样品采集时的大力协助!
[1] Anderson D M,Glibert P M,Burkholder J M.Harmful algal blooms and eutrophication:nutrient sources,composition,and consequences[J].Estuaries and Coasts,2002,25:704-726.
[2] Li M,XuK,Watanabe M,et al.Long-term variations in dissolved silicate,nitrogen,and phosphorus flux from the Yangtze River into the East China Sea and impacts on estuarine ecosystem[J].Estuarine,Coastal and Shelf Science,2007,71:3-12.
[3] Zhou M J,Shen Z L,Yu R C.Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang(Yangtze)River [J].Continental Shelf Research,2008,28:1483-1489.
[4] 俞志明,沈志良,陈亚瞿,等.长江口水域富营养化[M].北京:科学出版社,2011:1-549.
[5] Jiang T,Yu Z M,Song X X,et al.Long-term ecological interactions between nutrient and phytoplankton community in the Changjiang estuary[J]. Chinese Journal of Oceanology and Limnology,2010,28:887-898.
[6] Wang J,Wu J.Occurrence and potential risks of harmful algal blooms in the East China Sea[J].Science of the Total Environment,2009,407:4012 -4021.
[7] 郭玉洁,杨则禹.1976年夏季东海陆架区浮游植物生态的研究[J].海洋科学集刊,1982,19:11-32.
[8] 钱树本,陈国蔚.长江口及济州岛邻近海域综合调查研究报告[J].山东海洋学院学报,1986,16:26-85.
[9] 郭玉洁,杨则禹.长江口浮游植物的数量变动及生态分析[J].海洋科学集刊,1992,33:167-189.
[10] 王云龙,沈新强,李纯厚,等.中国大陆架及邻近海域浮游生物[M].上海:上海科学技术出版社,2005:35-67.
[11] Chen CS,Zhu J R,Beardsley R C,et al.Physical-biological sources for dense algal bloom near the Changjiang River[J].Geophysical Research Letters,2003,30:151-1518.
[12] XuK,Milliman J D.Seasonal variations of sediment discharge from the Yangtze River before and after impoundment of the Three Gorges Dam [J].Geomorphology,2009,104:276-283.
[13] 王云龙,袁骐,沈新强.长江口及邻近海域夏季浮游植物分布现状与变化趋势[J].海洋环境科学,2008,27(2):169-172.
[14] 林锋竹,吴玉霖,于海成,等.2004年长江口浮游植物群落结构特征分析[J].海洋与湖沼,2008,39(4):401-410.
[15] 王俊,陈瑞盛,左涛.三峡工程截流后长江口邻近海域的网采浮游植物群落结构特征[J].水生态学杂志,2009,2(2):80-87.
[16] 何青,孙军.长江口及其邻近水域网采浮游植物群落[J].生态学报,2009,29(7):3928-3938.
[17] Luan Q S,Sun J,Shen Z L,et al.Phytoplankton assemblage of Yangtze River estuary and the adjacent East China Sea.Journal of Ocean University of China[J].2006,5(2):123-131.
[18] 李云,李道季,唐静亮,等.长江口及毗邻海域浮游植物的分布与变化[J].环境科学,2007,28(4):719-729.
[19] 栾青杉,孙军.2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系[J].生态学报,2010,30:4967-4975.
[20] 王丹,孙军,周锋,等.2006年6月长江口低氧区及邻近水域浮游植物[J].海洋与湖沼,2008,39(6):619-626.
[21] 朱建荣,丁平兴,胡敦欣.2000年8月长江口外海区冲淡水和羽状锋的观测[J].海洋与湖沼,2003,34(3):249-255.
[22] 苏纪兰,袁业力.中国近海水文[M].北京:海洋出版社,2005:207-245.
[23] Zhu J R,Wang J H,Shen H T,et al.Observation and analysis of the diluted water and red tide in the sea off the Changjiang River mouth in middle and late June,2003[J].Chinese Science Bulletin,2005,50(3):59-65.
[24] Chen Y L L,Chen H Y,Gong G C,et al.Phytoplankton production during a summer coastal upwelling in the East China Sea[J].Continental Shelf Research,2004,24:1321-1338.
[25] 国家海洋局.海洋生物生态调查技术规程[S].北京:海洋出版社,2006:1-250.
[26] 张传松,王江涛,朱德弟,等.2005年春夏季东海赤潮过程中营养盐作用初探[J].海洋学报,2008,30(2):153-159.
[27] 顾新根,袁骐,杨蕉文,等.长江口羽状锋海区浮游植物的生态研究[J].中国水产科学,1995,2(1):1-15.
[28] 吴玉霖,傅月娜,张永山,等.长江口海域浮游植物分布及其与径流的关系[J].海洋与湖沼,2004,35(3):246-251.
[29] 余立华,李道季,方涛,等.三峡水库蓄水前后长江口水域夏季硅酸盐、溶解无机氮分布及硅氮比值的变化[J].生态学报,2006,26(9):2817 -2826.
[30] 李瑞香,朱明远,陈尚,等.围隔生态系内浮游植物对富磷的响应[J].生态学报,2001,21(4):603-607.
[31] Jiao N Z,Zhang Y,Zeng Y H,et al.Ecological anomalies in the East China Sea:impacts of the Three Gorges Dam[J].Water Research,2007,41:1287-1293.
[32] Jiang Z B,Liu J J,Chen J F,et al.Responses of summer phytoplankton community to drastic environmental changes in the Changjiang(Yangtze River)Estuary during the past 50 years[J].Water Research,2014,54:1-11.
[33] 冯琳,林霄沛.1945—2006年东中国海海表温度的长期变化趋势[J].中国海洋大学学报,2009,39(1):13-18.
[34] 蔡榕硕,陈际龙,黄荣辉.我国近海和邻近海的海洋环境对最近全球气候变化的响应[J].大气科学,2006,30(5):1019-1033.
Distribution of net-phytoplankton community in summer and its influence factors in the Changjiang Estuary and adjacent continental shelf
Jiang Zhibing1,2,Zeng Jiangning2,Li Hongliang2,Zhou Feng3,Chen Quanzhen2,Zhu Genhai2,Zhang Dongsheng2,Yan Xiaojun1
(1.Key Laboratory of AppliedMarine Biotechnology,Ministry of Education,Marine College of Ningbo University,Ningbo 315211,China;2.Key Laboratory of Marine Ecosystem andBiogeochemistry,SecondInstitute of Oceanography,State Oceanic Ad-ministration,Hangzhou 310012,China;3.StateKey Laboratory of Satellite Ocean Environment Dynamics,SecondInstitute of Oceanography,State Oceanic Administration,Hangzhou 310012,China)
Net-phytoplankton samples were collected from the Changjiang Estuary(CE)and adjacent continental shelf,East China Sea in June and August 2009.A total of 395 taxa(9 phyla)were identified,including 223 diatom and 125 dinoflagellate species.The phytoplankton abundance in August(3 077.15×104cells/m3)was significantly higher than that in June(107.80×104cells/m3).The high-density zone in the CE moved eastward from 31°N,123°E in June to 31°N,123.5°E in August due to the enhancement of Changjiang Diluted Water.The species richness of net-phytoplankton in August was much higher than that in June,while species diversity and evenness were slightly lower than that in June.Pseudo-nitzschia pungens and Ceratium tripos were the dominant species in June. However,P.pungens,Rhizosolenia styliformis and Trichodesmium thiebautii dominated in August.S犽eletonema spp.had high abundance in the CE in both June and August,although it was not a dominant species.The analysis of similarity and non-metric multidimensional scaling showed a significantly spatio-temporal difference in phytoplankton community.Canonical correspondence analysis indicated that temperature and salinity were the primary variables affecting the community distribution in June and August,respectively.These data demonstrated that the distribution of phytoplankton community in the CE and adjacent continental shelf was intensively influenced by the circulations and water masses.
East China Sea;Changjiang Estuary;phytoplankton,spatio-temporal distribution;environmental factors
Q178.53
A
0253-4193(2014)06-0112-12
2013-05-15;
2013-12-08。
国家重点基础研究发展规划项目(2010CB428903);海洋公益性行业科研专项(201305009,201205015,201105014);国家自然科学科学基金(41206103,41206104,41176142);国家海洋局青年海洋科学基金(2011106);国家海洋局第二海洋研究所基本科研业务费专项(JG1222)。
江志兵(1983—),男,浙江省温岭市人,助理研究员,主要从事海洋浮游植物研究。E-mail:jzb1217@126.com
*通信作者:严小军,博士,研究员,主要从事海洋生物技术研究。E-mail:yanxiaojun@nbu.edu.cn
江志兵,曾江宁,李宏亮,等.长江口及其邻近陆架区夏季网采浮游植物及其影响因素[J].海洋学报,2014,36(6):112—123,
10.3969/j.issn.0253-4193.2014.06.014
Jiang Zhibing,Zeng Jiangning,Li Hongliang,et al.Distribution of net-phytoplankton community in summer and its influence factors in the Changjiang Estuary and adjacent continental shelf[J].Acta Oceanologica Sinica(in Chinese),2014,36(6):112—123,doi:10.3969/j.issn.0253-4193.2014.06.014
猜你喜欢
吉林大学学报(地球科学版)(2024年3期)2024-06-03 13:22:07
大自然探索(2023年7期)2023-11-14 13:07:36
课堂内外(小学版)(2023年9期)2023-10-11 14:39:26
黑龙江水利科技(2020年8期)2020-01-12 06:26:00
学与玩(2019年8期)2019-10-29 03:32:16
资源节约与环保(2019年10期)2019-01-21 00:22:48
上海建材(2018年1期)2018-04-18 12:15:16
海洋渔业(2016年6期)2016-04-16 03:00:35
海洋渔业(2016年6期)2016-04-16 03:00:32
中国酿造(2014年9期)2014-03-11 20:21:07
