CKLF1通过激活PLCγ/FAK信号途径促进SH-SY5Y细胞迁移
2014-05-18 08:07王真真苑玉和陈乃宏
中国药理学通报 2014年9期
王真真,苑玉和,韩 宁,张 毅,陈乃宏
(1.中国医学科学院神经科学中心,中国医学科学院·北京协和医学院药物研究所,天然药物活性物质与功能国家重点实验室,北京 100050;2.北京中医药大学基础医学院解剖学教研室,北京 100029)
趋化因子(chemotactic cytokines,chemokines)属于小分子的分泌蛋白超家族(Mr=8 000~12 000,含70~90个氨基酸),是具有趋化作用的一类细胞因子[1],根据肽链N端高度保守的半胱氨酸残基的数量和相对位置的差异,可将趋化因子及其受体分为4个亚家族:CXC、CC、C和 CX3C[2-4],其中 C代表半胱氨酸,X代表任意一种氨基酸。趋化因子最初被认识是基于其调节免疫细胞游走的能力,深入的研究发现趋化因子的生物学功能不仅仅限于免疫系统,在神经系统中也发挥了重要的作用,不但涉及到神经炎症过程,还参与了神经系统中与炎症无关的生理功能[5-7]。
趋化素样因子超家族(chemokine-like factor super family,CKLFSF)是北京大学人类疾病基因研究中心在国际上首次报道的一个新的基因家族[8-11],该家族包括9个基因,其中趋化素样因子1(chemokine-like factor 1,CKLF1;原名 UCK-1,U937 derived chemokine-1)是该家族中第一个发现的CC类趋化因子(GenBank编号:AF096895)[12]。研究发现,CKLF1在 免 疫[12-13]、造 血[14-15]、骨 骼[16]和 呼吸[17-18]等系统发挥重要的作用,那么CKLF1是否也参与了神经系统的功能呢?本研究旨在探索CKLF1在SH-SY5Y细胞迁移过程中所发挥的作用,以及其可能的分子机制。
1 材料与方法
1.1 细胞 SH-SY5Y细胞为本室保存。
1.2 药品及试剂 CKLF1多肽购自上海吉尔生化公司(共计27个氨基酸,经HPLC和MS鉴定,纯度≥95%);SDF-1α购自美国 PeproTech公司;FAK-pY397磷酸化抗体购自美国Upstate公司;β-actin多克隆抗体购自美国Santa Cruz公司;BSA购自美国Sigma公司;BCA蛋白定量试剂盒购自维格拉斯生物技术有限公司;胎牛血清、RPMI 1640培养基购自美国Gibco公司;其它各种化学试剂为进口或国产分析纯试剂。
1.3 细胞培养 将SH-SY5Y细胞置于含10%胎牛血清的RPMI1640培养基,于37℃、5%CO2的孵箱中培养。
1.4 CELLocate细胞定位实验 将刻有26个英文字母和10个阿拉伯数字的微网格玻片(购自美国Eppendorf公司)置于24孔板中,再将细胞以2.0×106·L-1接种于24孔板中,待细胞贴壁后分别加入溶剂对照、CKLF1(200 nmol·L-1)、SDF-1α(100 μg·L-1),观察并拍摄不同时间点(0.5、2、8、24 h)细胞的定位情况,利用每个小方格边界长度为175 μm,对细胞迁移运动的距离进行测量和计算,同时统计迁移细胞占所有细胞的百分比。
1.5 蛋白免疫印迹 细胞经PBS清洗后,用NP-40裂解液(50 mmol·L-1Tris-HCl,150 mmol·L-1NaCl,10 g·L-1NP-40,1 mmol·L-1EDTA,1 g·L-1SDS,1 mmol·L-1PMSF,10 mg·L-1异亮肽酶,20 mg·L-1抑肽酶,1 mg·L-1胃蛋白酶,1 mmol·L-1Na3VO4,10 mmol·L-1NaF)进行裂解,蛋白浓度用BCA试剂盒进行测定,细胞裂解物溶于上样缓冲溶液后经SDS-PAGE电泳分离,并电转到PVDF膜上,经30 g·L-1BSA封闭后,与一抗孵育,再与辣根过氧化物酶标记的二抗结合后,用ECL化学发光法进行检测。
1.6 趋化实验 采用Boyden趋化小室法行体外细胞趋化活性分析。向24孔板内加入600μl含不同浓度CKLF1的RPMI 1640培养基,同时以不含趋化因子的RPMI 1640培养基作为阴性对照,将覆盖有8μm孔径多聚碳酸酯膜的趋化小室插入24孔板内,用含0.1%BSA的无血清RPMI 1640培养基将SH-SY5Y细胞制悬,以5×108·L-1的密度接种于趋化小室的上层(100μl/孔),在 37℃、5%CO2条件下培养8~12 h。用棉签轻轻擦去膜表面残留的细胞,置于10%中性福尔马林溶液中固定。用1%草酸铵结晶紫染色10~15 min后双蒸水漂洗。倒置显微镜下观察,随机计数5个高倍视野(400×)下迁移至多聚碳酸酯膜下表面的SH-SY5Y细胞平均数,并按公式计算趋化指数(chemotactic index,CI),当 CI>2时认为有意义。

1.7 统计学处理 实验数据以¯x±s表示,各组间差异的比较应用GraphPad Prism 5.0软件进行单因素方差分析(One-Way ANOVA)或双因素方差分析(Two-Way ANOVA)。
2 结果
2.1 CKLF1能够促进SH-SY5Y细胞的迁移运动采用CELLocate细胞定位实验检测SH-SY5Y细胞在200 nmol·L-1CKLF1刺激下在CELLocate载玻片上的迁移能力。细胞在37°C孵箱中分别孵育0.5、2、8、24 h后镜下观察细胞位置的改变。如Fig 1所示,随着与CKLF1孵育时间的延长,细胞迁移距离和迁移细胞百分比均呈现上升趋势。与空白对照组相比,孵育8 h组和24 h组的细胞迁移距离差异有显著性(P<0.05,P<0.05),且与阳性对照组效应相当(P<0.05,P<0.05)。与空白对照组相比,孵育8 h组的迁移细胞百分比差异有显著性(P<0.05),且与阳性对照组效应相当(P<0.01)。结果表明,CKLF1(200 nmol·L-1)可促进 SH-SY5Y细胞的迁移运动,在刺激8 h时的效应最为明显。
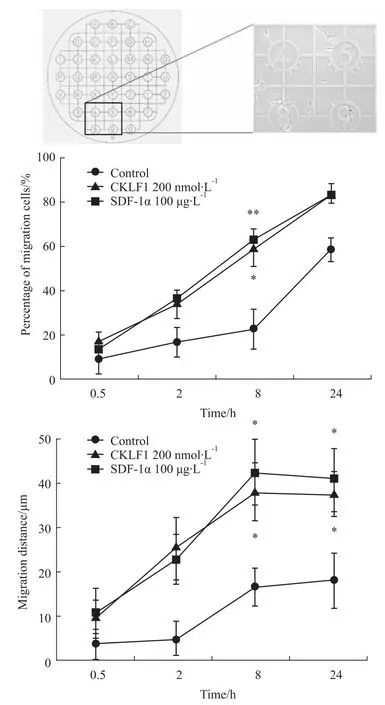
Fig 1 M igration distance and the percentage ofm igration cells(¯x±s)SH-SY5Y cells were stimulated with CKLF1 at the indicated time points.Exact relocation of the areawasenabled by CELLocate coverslipswith engraved alphanumerically labelled grids.SDF-1αwas regarded as the positive control.*P<0.05,**P<0.01 vs control.
2.2 抑制PLCγ可逆转CKLF1诱导的FAK磷酸化水平的升高 以1.0×106/孔的密度将SH-SY5Y细胞接种到6孔板中,待细胞贴壁后加入PLCγ抑制剂U73122(1μmol·L-1)或其溶剂对照预孵育1 h,再加入 CKLF1(200 nmol·L-1)刺激 8~12 h,阴性对照组不加CKLF1刺激,然后行Western blot检测。如Fig 2所示,与空白对照组相比,CKLF1刺激组的FAK-pY397磷酸化水平明显升高(P<0.05),而同时给予U73122和CKLF1处理组的FAK-pY397磷酸化水平,与CKLF1刺激组相比则明显降低(P<0.05)。结果表明,CKLF1能够通过激活PLCγ增强FAK-pY397位点的磷酸化水平。
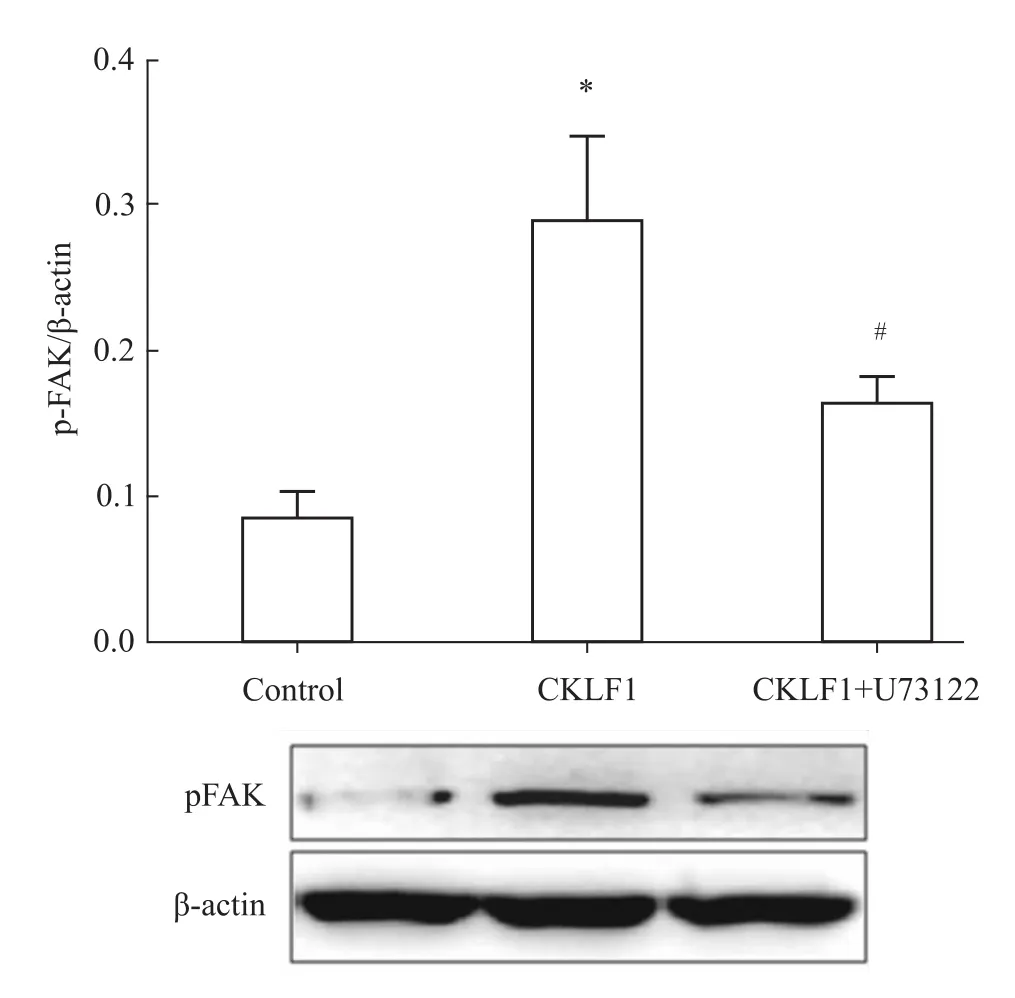
Fig 2 PLCγinhibitor U73122 inhibits CKLF1-induced phosphorylation of FAK(¯x±s)SH-SY5Y cellswere plated onto 6-well plates at a density of 1.0×106 cells per well.Cellswere pretreated with PLCγinhibitor U73122(1 μmol·L-1)or the vehicle for1 h before the stimulaton of CKLF1(200 nmol·L-1)for 8-12 h.*P<0.05 vs control;#P<0.05 vs CKLF1.
2.3 抑制FAK或PLCγ能够降低CKLF1的趋化活性 分别将 FAK抑制剂 PF-573228(10μmol·L-1)或 PLCγ抑制剂 U73122(1μmol·L-1)或其溶剂对照加入SH-SY5Y细胞预孵育1 h,然后将细胞制悬,加入趋化小室上层,趋化小室下层为含CKLF1(200 nmol·L-1)的培养基,阴性对照组的趋化小室下层则为不含CKLF1的培养基,37℃孵育8~12 h。然后按照趋化实验方法检测各组CI值。如Fig 3所示,与空白对照组相比,CKLF1刺激组CI值明显升高(P<0.01),而PF-573228或U73122预处理组的CI值与CKLF1刺激组相比则明显降低(P<0.01)。结果表明,CKLF1对SH-SY5Y细胞具有明显的趋化活性,而这一作用可被 FAK抑制剂或PLCγ抑制剂明显抑制。
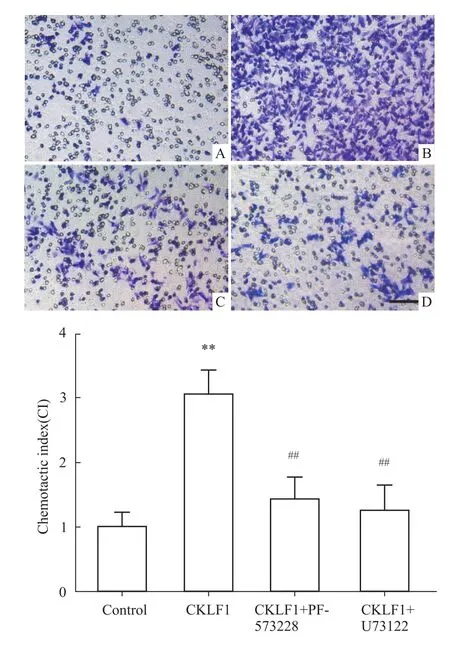
Fig 3 Effects of FAK inhibitor and PLCγinhibitor on CKLF1-induced m igration(¯x±s)Vehicle(B),PF-573228(C)or U73122(D)pre-treated SHSY5Y cells were allowed to migrate through an 8-μm pore membrane which immersed in the medium containing CKLF1(200 nmol·L-1).The transmigrated cellswere harvested after incubation 8-12 h at37℃.The negative control cells(A)were allowed tomigrate throughmembrane which immersed in themedium without CKLF1 instead.Scale bar=100 μm.Chemotactic index(CI)showed that CKLF1 significantly promoted the chemotaxis in SH-SY5Y cells,while themigrating cells obviously decreased in the PF-573228 and U73122 pre-treated groups.**P<0.01 vs control;##P<0.01 vs CKLF1.
3 讨论
哺乳动物最早期的神经系统起源于胚胎的神经外胚层,神经外胚层卷曲形成神经管[19-20]。神经管开始只有一层细胞,而成熟的大脑皮层有多层细胞。胚胎学家通过对胚胎的形态观察到细胞层数变化而提出在胚胎发育期间和出生后,包括人在内的哺乳动物神经系统的大部分神经细胞(也许是所有神经细胞)都要经过一定距离的迁移运动才能抵达它们发挥功能的部位[21-23]。神经细胞迁移不能正常进行时,会导致神经系统发育异常,可以产生多种疾病[24-26],一部分癫痫就是因为神经细胞迁移异常而造成的[27]。因此,研究神经细胞迁移对于我们了解神经系统功能是必不可少的。
我们前期的研究发现,趋化因子CKLF1在脑组织中的表达部位恰好与神经细胞的迁移路线相吻合,而且CKLF1的表达在时间上与脑发育阶段并行[7],这一有趣的现象是否只是巧合呢?虽然CKLF1对中性粒细胞、单核细胞、淋巴细胞等免疫系统细胞具有化学吸引作用,但是CKLF1在神经系统中的功能尚不完全清楚,因此,本文重点研究了CKLF1对神经系统细胞系SH-SY5Y细胞迁移运动的影响。结果发现,用CKLF1活性片段刺激SH-SY5Y细胞,能够诱导其迁移运动,表明CKLF1对神经细胞也有促进迁移的能力。
我们进一步对CKLF1促进SH-SY5Y细胞迁移可能的分子机制进行了研究。细胞迁移是细胞在接收到迁移信号或感受到某些物质的浓度梯度后而产生的移动,由4个循环往复的步骤组成,即头部伪足的形成和延伸、新黏附位点的建立、胞体的收缩以及尾部的退缩,通过不断重复的4个过程向前迁移[28]。我们前期已经发现,CKLF1可促进骨架蛋白的聚合和伪足的形成[6],因此本文对细胞迁移运动的第2个步骤新黏附位点的建立进行了研究,主要着眼于参与黏着斑形成的黏着斑激酶FAK。结果发现,CKLF1活性片段刺激SH-SY5Y细胞可明显升高FAK-pY397位点的磷酸化水平激活FAK,而FAK受到抑制后,CKLF1的趋化活性则明显降低,提示FAK-pY397位点磷酸化在CKLF1诱导的SH-SY5Y细胞迁移过程中发挥了重要的调节作用。我们进一步对FAK的上游分子进行了研究,结果发现,抑制PLCγ可明显逆转CKLF1所诱导的FAK-pY397位点磷酸化水平升高;此外,CKLF1的趋化活性也能够被PLCγ抑制剂U73122所抑制。这些结果表明,PLCγ可通过影响FAK的磷酸化水平调节CKLF1的趋化活性,CKLF1通过激活PLCγ/FAK信号途径促进SH-SY5Y细胞的迁移运动。
参考文献:
[1] Adler MW,Geller E B,Chen X,etal.Viewing chemokinesasa third major system of communication in the brain[J].AAPS J,2005,7(4):E865-70.
[2] Dijkstra IM,Hulshof S,van der Valk P,etal.Cutting edge:activity of human adultmicroglia in response to CC chemokine ligand 21[J].J Immunol,2004,172(5):2744-7.
[3] Murphy PM,BaggioliniM,Charo IF,et al.International union of pharmacology.XXII.Nomenclature for chemokine receptors[J].Pharmacol Rev,2000,52(1):145-76.
[4] Tran PB,Miller R J.Chemokine receptors:signposts to brain development and disease[J].Nat Rev Neurosci,2003,4(6):444-55.
[5] 王真真,胡金凤,李 刚,等.趋化因子及其受体在神经系统发育中的作用[J].生命科学,2007,19(5):536-42.
[5] Wang Z Z,Hu JF,Li G,et al.Effects of chemokines and their receptors on the developmentof nervous system[J].Chin Bull Life Sci,2007,19(5):536-42.
[6] Wang Z Z,Li G,Chen X Y,et al.Chemokine-like factor 1,a novel cytokine,induces nerve cellmigration through the non-extracellular Ca2+-dependent tyrosine kinases pathway[J].Brain Res,2010,1308:24-34.
[7] Wang Z Z,Zhang Y,Yuan Y H,et al.Developmental expression of chemokine-like factor1,a novelmember of chemokines family,in postnatal rat cerebral cortex[J].Neurosci Lett,2012,519(1):51-5.
[8] Han W,Ding P,Xu M,et al.Identification of eight genes encoding chemokine-like factor superfamily members 1-8(CKLFSF1-8)by in silico cloning and experimental validation[J].Genomics,2003,81(6):609-17.
[9] Shi S,Rui M,Han W,et al.CKLFSF2 is highly expressed in testis and can be secreted into the seminiferous tubules[J].Int J Biochem Cell Biol,2005,37(8):1633-40.
[10]Li T,Han W,Yang T,etal.Molecular cloning and identification ofmouse Cklfsf2a and Cklfsf2b,two homologues of human CKLFSF2[J].Int JBiochem Cell Biol,2006,38(3):420-9.
[11]Zhong J,Wang Y,Qiu X,et al.Characterization and expression profile of CMTM3/CKLFSF3[J].JBiochem Mol Biol,2006,39(5):537-45.
[12]Han W,Lou Y,Tang J,et al.Molecular cloning and characterization of chemokine-like factor 1(CKLF1),a novel human cytokine with unique structure and potential chemotactic activity[J].Biochem J,2001,357(Pt1):127-35.
[13]Lou Y,Xia D,Han W,etal.Molecular cloning and characterization of rat chemokine-like factor 1 and 2[J].Gene,2003,307:125-32.
[14]克晓燕,贾丽萍,景红梅,等.新的人趋化素样因子对骨髓造血干/祖细胞的体外刺激作用[J].中华血液学杂志,2002,23(6):301-3.
[14]Ke X Y,Jia L P,Jing H M,etal.Effects of novel human chemokine-like factor 1(CKLF1)on bone marrow hematopoietic stem cell/progenitor cell in vitro[J].Chin JHematol,2002,23(6):301-3.
[15]韩文玲,芮 珉,张颖妹,等.趋化素样因子(CKLF1)对骨髓细胞增殖活性的研究[J].中国医学科学院学报,2001,23(2):119-22.
[15]Han W L,RuiM,Zhang YM,etal.Stimulatory effectof chemokine-like factor 1(CKLF1)on the growth of bonemarrow cells[J].Chin Med Sci J,2001,23(2):119-22.
[16]程爱新,韩文玲,马大龙,等.趋化素样因子1(CKLF1)对关节软骨细胞增殖及代谢的影响[J].北京大学学报(医学版),2003,35(4):399-401.
[16]Cheng A X,Han W L,Ma D L,et al.Effects of chemokine-like factor 1(CKLF1)on proliferation and metabolism of chondrocytes[J].JPeking Univ(Health Sci),2003,35(4):399-401.
[17]Tan Y X,Han W L,Chen Y Y,etal.Chemokine-like factor1,a novel cytokine,contributes to airway damage,remodeling and pulmonary fibrosis[J].Chin Med J(Engl),2004,117(8):1123-9.
[18]李光艳,宋修云,胡金凤,等.CKLF1高表达激活NF-κB信号通路导致小鼠肺部炎症[J].中国药理学通报,2013,29(10):1359-62.
[18]Li G Y,Song X Y,Hu JF,et al.High expression of CKLF1-induced lung inflammation by NF-κB signal pathway in mice[J].Chin Pharmacol Bull,2013,29(10):1359-62.
[19]Kwon Y,Shin J,Park HW,etal.Dynamic expression pattern of Hoxc8 duringmouse early embryogenesis[J].Anat Rec A Discov Mol Cell Evol Biol,2005,283(1):187-92.
[20]Pankratz M T,Li X J,Lavaute TM,etal.Directed neural differentiation of human embryonic stem cells via an obligated primitive anterior stage[J].Stem Cells,2007,25(6):1511-20.
[21]Chittajallu R,Kunze A,Mangin JM,et al.Differential synaptic integration of interneurons in the outer and inner molecular layers of the developing dentate gyrus[J].J Neurosci,2007,27(31):8219-25.
[22]Fukumitsu H,Ohtsuka M,Murai R,et al.Brain-derived neurotrophic factor participates in determination of neuronal laminar fate in the developingmouse cerebral cortex[J].JNeurosci,2006,26(51):13218-30.
[23]Molnar Z,Metin C,Stoykova A,et al.Comparative aspects of cerebral cortical development[J].Eur JNeurosci,2006,23(4):921-34.
[24]Barth PG.Disorders of neuronalmigration[J].Can JNeurol Sci,1987,14(1):1-16.
[25]Toi A,Chitayat D,Blaser S.Abnormalities of the foetal cerebral cortex[J].Prenat Diagn,2009,29(4):355-71.
[26]Spalice A,Parisi P,Nicita F,et al.Neuronal migration disorders:clinical,neuroradiologic and geneticsaspects[J].Acta Paediatr,2009,98(3):421-33.
[27]Gordon N.Epilepsy and disorders of neuronalmigration.II:Epilepsy as a symptom of neuronal migration defects[J].Dev Med Child Neurol,1996,38(12):1131-4.
[28]Schaar B T,McConnell SK.Cytoskeletal coordination during neuronalmigration[J].Proc Natl Acad Sci USA,2005,102(38):13652-7.
猜你喜欢
生物化工(2022年4期)2022-09-20
数学年刊A辑(中文版)(2022年1期)2022-08-20
数学物理学报(2021年5期)2021-11-19
东坡赤壁诗词(2021年2期)2021-06-01
世界最新医学信息文摘(2021年55期)2021-01-07
环球时报(2019-01-29)2019-01-29
畅谈(2018年6期)2018-08-28
天津农业科学(2018年5期)2018-06-23
天津农业科学(2017年8期)2017-08-11
中国医药生物技术(2015年4期)2015-12-26