
拉鲁湿地自然保护区有壳肉足虫物种多样性与空间分布特征
2014-05-04 14:06:04巴桑普布
西藏大学学报(自然科学版) 2014年1期
巴桑 普布
(西藏大学理学院 西藏拉萨 850000)
拉鲁湿地自然保护区有壳肉足虫物种多样性与空间分布特征
巴桑 普布
(西藏大学理学院 西藏拉萨 850000)
摘要:2009年7月~2011年7月,用定点随机采样、非淹没培养皿法和活体观察法,对拉鲁湿地自然保护区有壳肉足虫物种多样性和空间分布特征进行了研究。共鉴定到有壳肉足虫61种(包括7个未定名种),隶属于3纲3目11科17属。其中表壳目44种,占有壳肉足虫物种总数的72.13%,为优势类群;网足目16种,占26.23%,为次优势类群;单室目1种,占有壳肉足虫总数的1.63%,为罕见类群。优势种类仅为盘状匣壳虫(Centropyxis discoides)。结果表明:拉鲁湿地有壳肉足虫群落结构较为复杂,各样点有壳肉足虫的物种相似性系数在0.44~0.70之间,为中等相似,有壳肉足虫群落特征区域差异性不明显;多样性指数在2.40~5.42之间,多样性指数具有明显的差异性。
关键词:有壳肉足虫;物种多样性;空间分布;拉鲁湿地
有壳肉足虫(Testate amoebae)是一类广泛分布于河流、湖泊、土壤和海洋乃至极端环境如极地等地的自由生活单细胞真核生物。其分类地位属原生动物亚界(Protozoa),肉鞭门(Sarcomastigophora),肉足亚门(Sarcodina),根足总纲(Rhizopoda)[1-3],对陆地和水生生态系统的结构和功能具有重要作用[4]。国际上对有壳肉足虫的研究始于1815年,迄今已描述2000多种。我国对淡水和土壤有壳肉足虫的工作分别始于1925年和1983年[5-6]。近80年来,学者们在有壳肉足虫的分类与区系方面作了一些工作,积累了一定的资料,已报道淡水种类233种和土壤种类64种[7-19]。西藏作为“地球三极之一”,特殊的地理环境孕育了丰富且独特的生物多样性,使该地区的生态系统对于全球变化特别是气候变化十分敏感,是研究全球气候变化生态效应的关键地区,因此西藏地区的生物多样性备受国内外专家的关注。有关西藏地区有壳肉足虫的研究开始于20世纪初,到目前只有极个别亚洲学者的报道[20-24]。但对拉鲁湿地自然保护区的有壳肉足虫未见专门的研究报道。2009年7月~2011年7月,对拉鲁湿地自然保护区有壳肉足虫的物种多样性及空间分布特征进行了较为系统的研究,目的在于初步摸清该湿地有壳肉足虫的物种组成、分布和空间变化特点,为保护该湿地的环境提供基础数据,也为我国淡水有壳肉足虫的生物多样性研究积累基础资料,同时对该地区自然资源开发与利用、水环境维持与改良等方面有着重要意义。
1 研究样区与方法
1.1 样区概况
拉鲁湿地位于拉萨河流域下游,拉萨市区西北角(N 29°39′46″~29°41'06″,E 91°03'49″~91°06'51″),平均海拔3645m。南北宽约2.5km,东西长约4.7km,总面积6.2km2。湿地气候为高原温带半干旱、半湿润季风气候,年日照时数2 811~3 157 h,年太阳总辐射量7 000~7 800 MJ/m2,多年平均降水量为439.3 mm,多年平均气温7.6℃,历史极端最高气温为29.9℃,极端最低气温-16.5℃。来自夺底、娘热两大沟的水系(泥沙河)、天然降水以及纳金电站的尾水为湿地主要补给水源。该湿地是海拔最高的天然内陆湿地,也是我国面积最大的城市天然湿地。拉鲁湿地具有丰富的动植物资源,为当地社会发展带来了巨大的经济、社会和生态效益[25-27]。拉鲁湿地植被以沼泽和草甸为主,其次是荒漠。沼泽湿地约占湿地总面积的60%,草甸沼泽湿地约占拉鲁湿地总面积的20%,草甸是由沼泽逐渐退化演变而形成。植被以芦苇为优势群落,其生长水域深0.5~1.4m,盖度80%~95%,群落生物量482.9~1 027.0g/m2;苔草和嵩草群落最复杂,其中苔草群落分布水深0.05~0.30m,盖度95%以上,群落生物量490.5~677.5g/m2;嵩草群落分布水深0~0.1m,盖度80%~90%,群落生物量294~434.9g/m2;最单调的是穗状狐尾藻群落,其生长域水深0.2~0.4m,群落盖度90%以上,群落生物量149.5~378.0g/m2。随着城市建设的发展、资源过度开发等原因,拉鲁湿地面积缩小了近6km2。这种状况如果继续加剧,会直接导致拉鲁湿生态环境退化,这将会对拉萨市城市环境发展产生不利的影响。
1.2 研究方法
1.2.1 样点设置及采样
根据环境特点共设置了5个采样站点。1#站点为湿地水源入口;2#~4#站点为湿地核心区域,其附近为牲畜主要集聚区;5#站点受人为干扰小,自然环境保持较相对良好。采样点分布图和生境特征详见文献[28]。
1.2.2样品采样与处理
水样采取随机采样法,用250ml的广口瓶随机采样,拿回实验室在显微镜下鉴定物种。采样时用便携式红外温度计(Fluke 62 Mini Infrared Thermometer,美国产)现场测试气温,用PCSTestr35型仪器(新加坡产)测得水体pH值、水温(T水)、电导率(EC)、总溶解盐 (TDS)以及盐度(Sal.)。
沼泽湿泥有壳肉足虫观察:取湿泥加土壤浸出液,充分搅拌,取悬浮液在显微镜下鉴定物种。
土壤有壳肉足虫观察:每份风干土样取10~50g于培养皿中,用“非淹没培养皿法”(non-flooded Petridish method)于恒温光照培养箱中在25℃左右的温度下培养。每份土样重复培养若干次,直到未出现新见物种为止。培养后第4d开始镜检。
1.2.3 种类鉴定
用活体镜检的方法和技术在显微镜下对有壳肉足虫种类进行鉴定并分类,种类鉴定和分类依据见文献[29-33]。
1.2.4 群落结构分析
(1)优势类群划分:优势类群和罕见类群的划分[34]。
(2)指数选取:包括Gleason-Margalef物种多样性指数、丰富度指数[35]和Jaccard群落相似性系数[36]。
(3)数据处理:采用SPSS15.0和Excel软件进行数据分析。
2 结果与讨论
2.1 水体理化指标
拉鲁湿地5个站点的主要理化指标详见文献[37]。
2.2 物种组成及分布
拉鲁湿地5个采样站点,共鉴定出有壳肉足虫61种,其中7个未定名种(见表1)。
从表1可以看出,有壳肉足虫物种在各个样点的分布存在明显差异。5#站点分布的种类最多,为37种,占物种总数的60.66%;1#站点分布的种类次之,为35种,占物种总数的57.38%;2#、3#和4#分布的种类相对较少,分别为32种、29和18种,占物种总数的52.46%、47.54%和29.51%。这正好反应1#与5#站点受人为干扰小,分布的种类相对丰富,而2#、3#和4#站点受人类活动干扰较多(如放牧和人为踩踏等活动),所以分布的种类较单一。数据表明,拉鲁湿地小环境存在着明显的空间差异性。
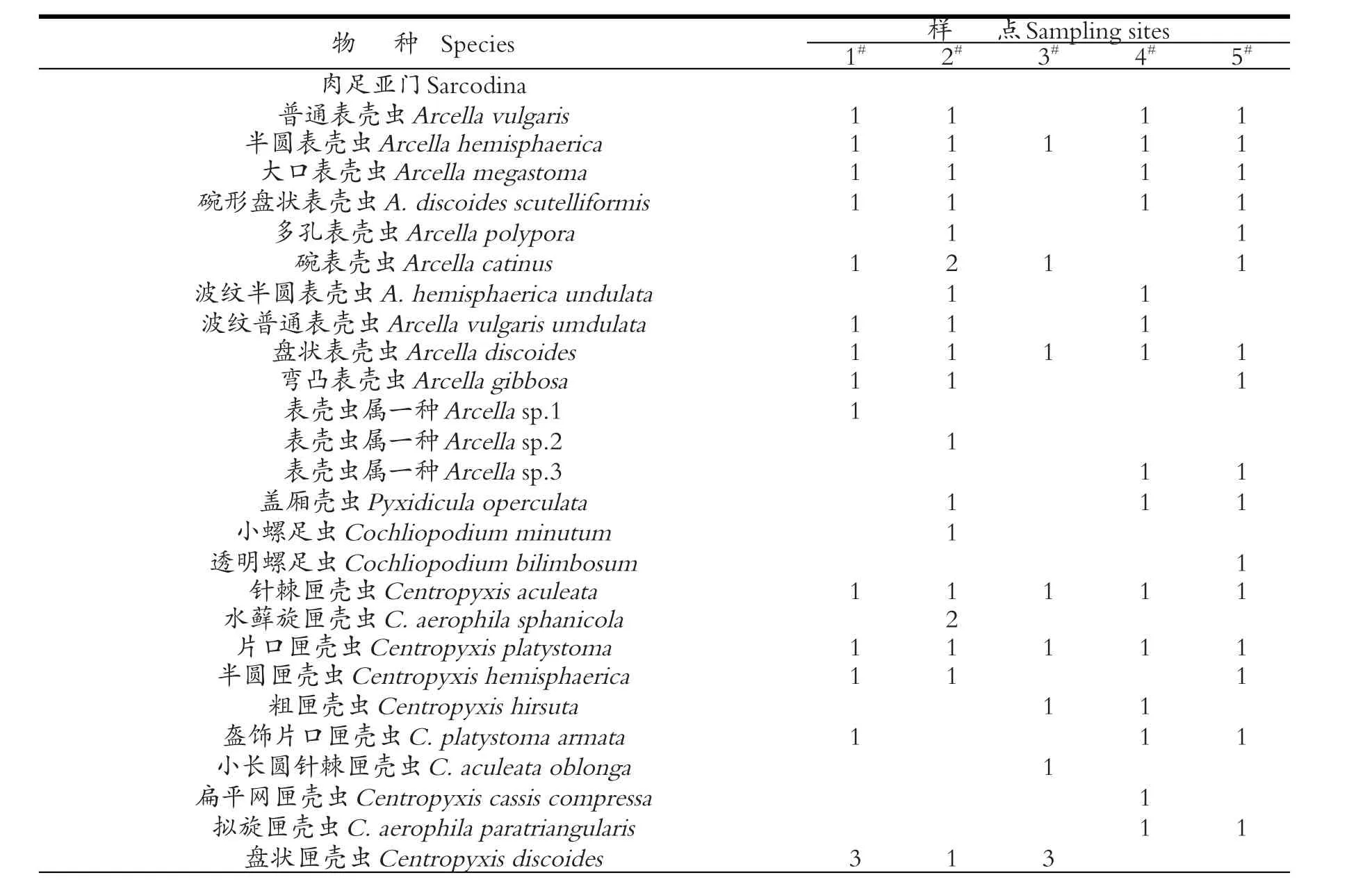
表1 拉鲁湿地有壳肉足虫种类及分布
2.3 群落结构特征
鉴定的61种有壳肉足虫,隶属于3纲3目11科17属。其中表壳目有44种,占有壳肉足虫物种总数的72.13%,为优势类群;网足目有16种,占26.23%,为次优势类群;单室目有1种,占有壳肉足虫总数的1.63%,为罕见类群。(见表2)。

表2 拉鲁湿地有壳肉足虫群落结构
2.4 群落多样性指数
根据Gleason-Margalef多样性性系数公式计算得到物种多样性系数(d),可以看出,拉鲁湿地1#样点的d值在最高,为5.42;3#样点d值最低,为2.40。各样点多样性指数依次为5#(5.42)>4#(4.86)>2#(4.63)>1#(4.61)>3#(2.40)。各样点d值在2.40~5.42之间,表明拉鲁湿地各样点多样性指数具有明显的差异性。
2.5 群落相似性系数
2.5.1 拉鲁湿地有壳肉足虫各样点群落相似性分析
对1#~5#站点分布的有壳肉足虫群落相似性(J)进行分析(见表3),可见,J值变化在0.4~40.70之间,为中等相似,结果表明拉鲁湿地有壳肉足虫群落特征区域性差异不明显。
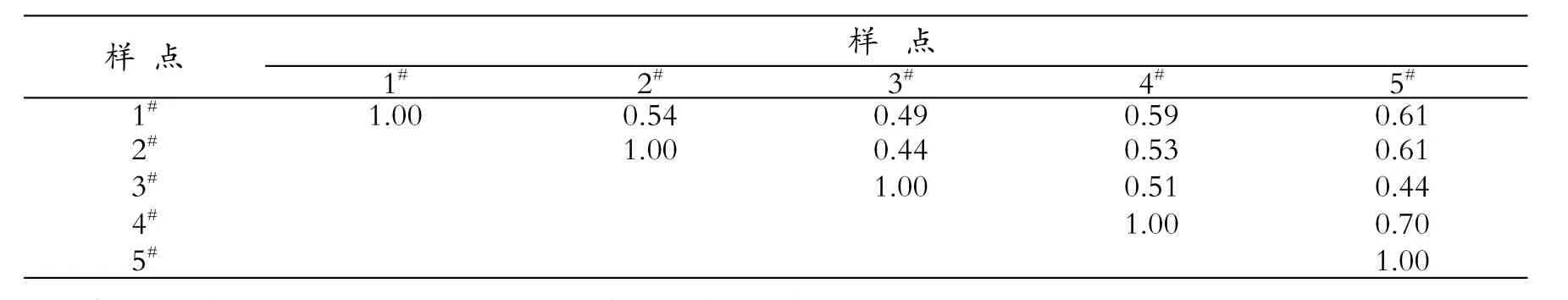
表3 拉鲁湿地各样点有壳肉足虫群落相似性系数
2.5.2 拉鲁湿地有壳肉足虫与内地湿地相似性系数比较
与吉林长白山、四川康定瓦斯河、上海崇明明珠湖、武汉南湖、甘肃黄河首曲湿地、新疆阿拉尔地区和北京野鸭湖湿地的淡水有壳肉足虫[38-44]相似性系数进行比较(见表4),可以看出,该湿地分布的有壳肉足虫与国内其他地区极不相似,表明拉鲁湿地自然保护区有壳肉足虫的物种组成具有其自身的独特性。

表4 拉鲁湿地自然保护区与中国其他地区淡水有壳肉足虫物种相似性系数
3 结论
3.1 种类组成与分布
在拉鲁湿地自然保护区5个采样站点共鉴定出61种有壳肉足虫(包括7个未定名种),未定名种占总种数的11.8%。各样点的物种组成各不相同,如5#样点的物种数最多(37种),占总种数的60.66%;3#样点的物种数最少(18种),占总数的29.51%;其余各样点分别占57.38%、52.46%和47.54%。表明拉鲁湿地各采样站点间的物种组成存在着明显的差异。各采样站点间的有壳肉足虫Jaccard群落相似性系数介于0.44~0.70之间,表现出较低的差异性。
3.2 群落结构与物种多样性
研究结果表明,拉鲁湿地自然保护区群落有壳肉足虫群落多样性普遍较高。调查研究中共鉴定出61种有壳肉足虫分别隶属于3纲3目11科17属61种。其中表壳目有44种,占有壳肉足虫物种总数的72.13%,为优势类群;网足目有16种,占26.23%,为次优势类群;单室目有1种,占有壳肉足虫总数的1.63%,为罕见类群。说明拉鲁湿地有壳肉足虫群落结构复杂,物种丰富度相对较高。多样性指数比较结果表明,各采样点间多样性具有一定的差别(2.40~5.42),表明拉鲁湿地有壳肉足虫物种多样性指数具有明显的差异。
3.3 物种多样性与环境理化因子间的关系
复杂的环境条件是丰富物种多样性的基础条件,拉鲁湿地自然保护区各采样站点间环境因子的差异决定了有壳肉足虫群落结构的复杂性及物种的多样性。在5个样点中,5#样点的pH值接近中性(7.22),水温相对较高,再加上受人类和家畜干扰因素较少,湿地保护较好,因此所鉴定出的有壳肉足虫物种最为丰富(37种),物种多样性指数也最高(5.42),而3#样点受人为干扰影响较大(家畜活动频繁区),所以物种数最少(18种),多样性指数最低(2.40)。通过理化因子的数据分析可看出,拉鲁湿地有壳肉足虫物种多样性受众多环境因素的影响,各样点之所以出现不同的数值,主要与人为干扰有直接的关联。
参考文献
[1][29]沈韫芬.原生动物学[M].北京:科学出版社,1999:231-293.
[2]Lee J J,Hutner,S H,Bovee E C1An Illustrated Guide to the Protozoa[M].1Lawrence,KS,USA:Allen Press,1985,158-252.
[3]Lee J J,Leedale GF,Bradbury P.The illustrated Guide to the protozoa(2nd Ed)[M].1Lawrence KSUSA:Allen Press,2000: 827-860,1054-1986.
[4]Wanner MA1reviewon the variability of testate amoebae:methodological approaches,environmental influences and taxonomical implication[J].Acts Protozool,1999,38:15-29.
[5]Wang C C.Study of the protozoa of Nanking,Part I[J].Contributions from the Biological Laboratory of the Science Society of China,1925,1(3):1-60.
[6]崔振东.长白山森林生态系统土壤原生动物初步研究[J].森林生态系研究,1983,3:144-153.
[7][20]王家楫.珠穆朗玛峰地区科学考察(1966-1968)生物与高山生理——珠穆朗玛峰地区的原生动物[M].北京:科学出版社,1974:145-1721.
[8]王家楫,沈韫芬,龚循矩.废水生物处理微型动物图志[M].北京:中国建筑工业出版社,1974:63-70.
[9][21]王家楫.西藏高原部分地区的原生动物[J].动物学报,1977,23(2):131-160.
[10]沈韫芬.西藏原生动物肉足虫一新种和两新亚种的描述[J].海洋与湖沼,1981,12(4):159-361.
[11][22][30]蒋燮治,沈韫芬,龚循矩.西藏水生无脊椎动物[M].北京:科学出版社,1983:48-100.
[12]尹文英.中国土壤动物检索图鉴[M].北京:科学出版社,1998:7-42,393-431.
[13]尹文英.中国土壤动物[M].北京:科学出版社,2000:209-220.
[14]宁应之,沈韫芬.中国典型地带土壤原生动物区系特征和物种分布[J].动物学报,1998,44(1):5-10.
[15]宁应之,沈韫芬.中国土壤原生动物新记录种——肉鞭虫门[J].动物学杂志,1999,34:2-5.
[16]王士正,马正学.西宁市工业废水纤毛虫类及三种高山、亚高山指示性有壳肉足虫[J].西北师范大学学报(自然科学版), 1994,30(1):60—65.
[17][31]杨军,冯伟松,缪炜.中国淡水与土壤有壳肉足虫最新分类名录及其区系分析[J].水生生物学报,2004,28(4):426-433.
[18][23]张文静,杨军,沈韫芬.中国有壳肉足虫(原生动物)五新纪录描述[J].动物分类学报,2009,34(3):686-690.
[19]杨军,Humphrey G.Smith,David M.Wilkinson.北极、南极和西藏有壳虫区系与分布[J].生物多样性,2010,18(4):373-382.
[20][24]袁显春,郑绵平,赵文,等.西藏阿里地区盐湖浮游生物生态调查[J].地质学报,2007,81(12):1754-1763.
[21][25]巴桑,次仁,普布,等.拉鲁湿地国家级自然保护区鸟类资源及其保护对策[J].资源科学,2009,31(7):1238-1243.
[22][26]陈德来,马正学,普布,等.拉鲁湿地夏季土壤动物的群落特征[J].动物学杂志,2011,46(5):1-7.
[23][27]刘汉武,立春.高原湿地研究——拉萨拉鲁地区湿地研究[M].北京:气象出版社,2005.
[24][28][37]巴桑,普布,马正学,等.西藏拉鲁湿地夏季和秋季浮游藻类群落特征[J].湿地科学,2012,10(4):404-416.
[25][32]沈韫芬,章宗涉,龚循矩,等.微型生物监测新技术[M].北京:中国建筑工业出版社,1990.
[26][33]Levine N.D,Corliss J O,COX F E G,et al.A newly revised classification of the protozoa[J].Journal of Protozoology, 1980,27(1):37-58.
[27][34]宁应之,李琦路,李晓鸿,等.甘肃白水江国家级自然保护区土壤肉鞭虫物种多样性[J].动物学杂志,Chinese Journal of Zoology,2007,42(4):81-88.
[28][35]Margalef R.Diversity and Stability in Ecological Systems[M].Upton:Brookhaven National Laboratory,1969.
[29][36]Jaccard P.Nouvelles recherches sur la distribution florale[J].Bulletin de la Société Vaudoise des Sciences Naturelles,1908, 44:223-270.
[30][38]李鸿凯,王升忠,卜兆君,等.长白山区典型泥炭藓沼泽的有壳变形虫[J].湿地科学,2010,8(3):249-255.
[31][39]吴志邦,谢嗣光,兰宝明.四川康定县瓦斯河龙洞水电站浮游动物调查初报[J].四川动物,2007,26(4):886-888.
[32][40]陈立婧,顾静,胡忠军.上海崇明明珠湖原生动物的群落结构[J].水产学报,2010,34(9):1404-1413.
[33][41]呼光富,刘红,马徐发.武汉南湖原生动物的初步研究[J].水利渔业,2007,27(3):77-78.
[34][42]程晓,刘汉成,杜海峰.甘肃黄河首曲湿地省级自然保护区肉鞭虫物种多样性研究[J].西北师范大学学报(自然科学版)2009,45(6):92-97.
[35][43]任道泉,程军,程勇.阿拉尔地区淡水原生动物调查初报[J].塔里木农垦大学学报,2002,14(1):18-20.
[36][44]高思佳.北京野鸭湖湿地浮游动物群落生态研究[D].北京:首都师范大学,2008.
2010年度教育部科学技术研究重点项目“西藏拉鲁湿地微型生物多样性与环境质量的生物效应研究”(项目号:210214);
2012年度教育部科学技术研究重点项目“拉萨河(拉萨段)表层沉积物金属污染时空特性研究”(项目号:212165)阶段性成果。
中图分类号:Q959
文献标识码:A
文章编号:1005-5738(2014)01-001-07
收稿日期:2013-10-24
基金项目:2014年度国家自然基金委地区基金项目“拉萨河流域原生动物群落结构特征及区系研究”(项目号:31360509);
第一作者简介:巴桑,男,藏族,西藏日喀则人,西藏大学理学院副教授,主要研究方向为湿地环境生态学。
[责任编辑:索郎桑姆]
Species diversity and spatial distribution of testate amoeba in the National Nature Reserve of Lhalu wetland,Lhasa
Ba Sang Pu Bu
(School of Science,Tibetan University,Lhasa 850000,Tibet)
Abstract:Species diversity and spatial variations during July 2009 to July 2011of testate amoeba was studied in Lhalu Wetland and randomly sampling,non-flooded petri dish and directly observation(in vivo)methods were applied.A total of 61 species(including 7 uncertain species)of testate amoeba were identified belonging to 17 genera,11 families,3 orders,3 classes.Amongst,44 species of arcellinida accounting for 72.13%of the total testate amoeba found in the study area was the dominant group;16 species of gromiida accounting for 26.23%of the total was the subdominant group;only one species(1.63%)of monothalamia found in the study area was occasional group.Centropyxis discoides was the only dominant species.The results showed that the community structure of testate amoeba in the wetland was relatively complex.The species similarity index of testate amoeba from different sampling sites in the wetland ranging between 0.44 and 0.70 implied that the spatial variations of testate amoeba was insignificant.However,diversity index,range between 2.40 and 5.42,suggested a significant spatial variability in diversity of testate amoeba in the study area.
Keywords:testate amoeba;species diversity;spatial distribution;Lhalu wetland
猜你喜欢
安徽农学通报(2016年24期)2017-01-12 21:15:16
绿色科技(2016年20期)2016-12-27 17:34:13
绿色科技(2016年21期)2016-12-27 10:53:26
现代农业科技(2016年20期)2016-12-20 09:17:04
科技创新导报(2016年21期)2016-12-17 12:53:50
中国民族民间医药·上半月(2016年10期)2016-11-19 11:41:11
科技视界(2016年18期)2016-11-03 23:51:58
中国科技博览(2016年22期)2016-11-01 16:58:26
现代经济信息(2016年13期)2016-06-17 21:16:45
科技视界(2016年13期)2016-06-13 08:06:36
