
ABA和PP33对国兰低温胁迫及恢复中光合作用和叶绿素荧光参数的影响
2014-04-29 00:44:03于永畅张林王厚新李承秀牛田颜婷美王长宪
农学学报 2014年4期
于永畅 张林 王厚新 李承秀 牛田 颜婷美 王长宪
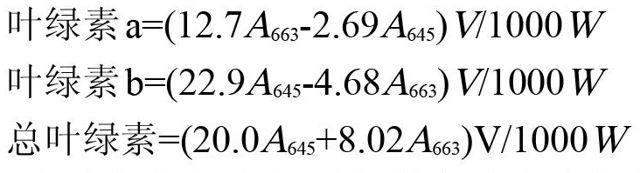
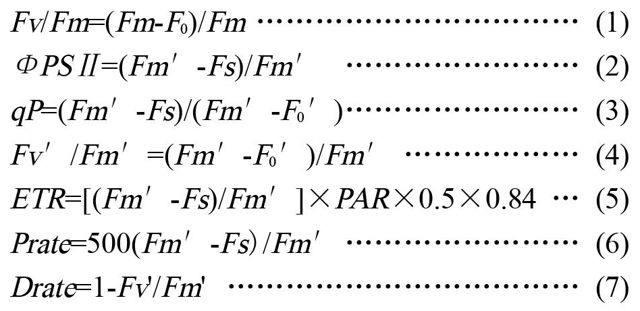
摘要:为探讨植物生长调节剂ABA和PP33对国兰低温胁迫下抗寒性的影响,以春兰名品‘宋梅为试材,采用20mg/LABA和300mg/L PP33对‘宋梅幼苗叶片进行叶面喷施和灌根处理,于光照培养箱[昼温/夜温=(5±0.5)℃/(0±0.5)℃]中进行低温胁迫9天,然后转入正常条件[昼温/夜温=(22±0.5)℃/(15±0.5)℃]恢复12天,研究不同低温处理及恢复时间对国兰光合作用和叶绿素荧光参数的影响。结果表明:低温导致春兰幼苗叶片的叶绿素含量、叶片净光合速率(Pn)、气孔导度(Gs)、气孔限制值(Ls)、PSⅡ最大光化学效率(Fv/Fm)、光化学猝灭系数(qP)、光合电子传递量子效率(φPSⅡ)、光适应下最大光化学效率(Fv'/Fm')、光合电子传递速率(ETR)和光化学耗散能量(Prate)下降,胞间CO2浓度(C1)、初始荧光(F0)和天线热耗散速率(Drate)则逐步上升,经ABA和PP33处理的植株指标波动幅度低于清水处理的植株,胁迫解除后能迅速恢复到对照水平,说明低温胁迫直接损伤光合机构使PSⅡ反应中心失活,而ABA和PP33可缓解PSⅡ反应中心受到的伤害,维持较高的光能捕捉能力和同化率,保证春兰幼苗叶片的光合作用能力。
关键词:春兰;抗寒性;光合作用;叶绿素荧光
中图分类号:S688
文献标志码:A
论文编号:2013-0754
0 引言
低温伤害是植物生长过程中常见的自然灾害,也是植物区域性分布和季节性生长的限制因素,它不仅导致植物产量的降低,严重时还会造成植株死亡。目前,在引种和培育抗寒品种过程中,评价植物抗寒性的常用方法是电导法、组织褐变法和2,3,5-氯化三苯基四氮唑还原法。近年来,有关光合和叶绿素荧光技术在植物抗逆性的研究领域逐渐增多,在植物抗寒性方面也显示出良好的应用前景。
兰花是中国十大传统名花之一,距今已有2500多年的栽培历史,国兰以其层出不穷的花艺、变幻莫测的叶艺受到爱花之士的喜爱。但是在日光温室国兰冬春季生产中,雨雪天气较多,温室内的兰花常处于低温的环境中,即白天温度常低于20℃、夜间温度常低于5℃。而兰花性喜温暖气候,一般国兰的生长适温为15-25℃,温度低于10℃生长缓慢,0℃低温便会造成不同程度的冻害发生,低温危害成为制约国兰在北方地区栽培生长的重要限制因素。通过一些途径提高植物的抗寒性,对于避免和减少植物寒害损失有重要意义。
植物抗逆化学诱导指的是利用外源化学物质模拟逆境信号或作为植物逆境信号传导过程中的中间信号物质,诱发植物启动抗性机制。植物激素在提高植物抗低温和微管抗冷稳定性方面有重要的作用,是植物抗寒基因表达的启动因子之一,同时外施植物激素类物质也能增强植物的抗寒性。内源脱落酸(ABA)是植物的“抗逆诱导因子”,诱导植物产生对不良生长环境的抗性。一般认为,ABA能提高植物的抗寒性,可能在于它能增强植物细胞保护系统的功能,提高细胞清除活性氧的能力,防止膜脂过氧化及稳定细胞膜结构,同时启动植物冷诱导基因的表达,提高植物抗寒性,脱落酸在植物抗寒性方面所起的作用已在葡萄、小麦等多种作物中得到证实。多效唑(PP333)也可提高多种植物的抗逆性,原因可能是多效唑可抑制植物体内赤霉素的合成,降低内源吲哚乙酸水平,提高吲哚乙酸氧化酶活性,增加细胞膜的稳定性,从而提高植物的抗逆性。目前,有关光合和叶绿素荧光机理在植物抗寒性方面的研究仅在黄瓜、水稻、番茄等少数作物上有相关报道,对于经过抗性处理的植物在低温胁迫下光合特性变化的研究较少,尤其在国兰方面尚未见相关报道。
光合作用是植物最基本的生命活动,低温胁迫不仅降低了叶绿体基质中进行的光合作用暗反应活性,还会引起类囊体膜介导的光反应活性降低,从而降低类囊体膜上的PS Ⅱ的光能传递效率和光能转换效率,导致CO2同化力降低,直接影响光合机构的性能和活性。光合机构中叶绿素吸收的光能主要有3个可能去向,即推动光合作用、转变成热散失和以荧光形式发射出来,由于这三者间存在此消彼长的关系,因此,通过荧光变化可以探测出光合作用和热耗散情况,逆境因子对植物光合作用的影响也可以通过它反映出来。本试验以春兰名品‘宋梅为试材,通过测定低温胁迫下经ABA和PP333处理的春兰幼苗光合作用和叶绿素荧光参数的变化,探讨低温胁迫对国兰光合机构的影响及ABA和PP333在提高国兰抗寒性方面的作用,为国兰在中国北方地区的选育栽培提供理论依据。
1 材料与方法
1.1试验材料及处理
试验于2012年10月-2013年2月在泰山林业科学研究院设施园艺中心和山东农业大学花卉研究室进行,供试材料为泰山林业科学研究院选育的春兰名品‘宋梅1年生幼苗。将兰花幼苗定植于装有植兰宝的花盆中,在日光温室中进行培养,当株高长至20cm左右,挑选生长健壮、长势一致的植株,用脱落酸(ABA,购白天津凯通化学试剂有限公司,为国产分析纯)、多效唑(PP333,购白天津凯通化学试剂有限公司,为国产分析纯)进行叶面喷施和灌根处理,ABA处理浓度为20mg/L、PP333处理浓度为300mg/L(前期试验发现20mg/L ABA和300mg/L PP333对提高国兰抗寒性方面具有显著影响)。采用随机区组设计,设3次重复,每小区10盆。为提高植株对生长调节剂的吸收效果,在第1次喷施后10天再进行第2次喷施,每次喷施以叶面附带小液珠为准,水肥管理按蕙兰的正常管理进行,共喷施5次。
处理完成后,将上述材料移入人工气候室,在昼温/夜温[(22±0.5)℃/(15±0.5)℃]适应2天,测定各参数值作为初始值,然后进行昼温/夜温[(5±0.5)℃/(0±0.5)℃]低温胁迫处理,光照时间为14h,光强为3000lx,相对湿度为80%。低温胁迫9天后转入昼温/夜温[(22±0.5)℃/(15±0.5)℃]中进行恢复生长,选取植株叶片的中上部,在低温胁迫处理后的第3、5、7、9天及恢复处理后的第3、6、9、12天测定植物叶片叶绿素含量及光合、荧光参数,以昼温/夜温[(22±0.5)℃/(15±0.5)℃]处理为CK,每项指标测定3株,3次重复。
1.2测定指标及方法
1.2.1叶绿素含量的测定 采用丙酮法测定叶绿素含量,称取叶片0.5g于研钵内,加入少量CaCO3和5mL80%的丙酮,将材料充分研磨细碎直至变白,转移至10mL具塞离心管中,定容至刻度,摇匀后置于4℃冰箱中静置12h,离心10min(2000r/min),取上清液在波长663、645和652mm下测定吸光度,以80%丙酮为空白对照,计算叶绿素a、b含量和叶绿素总量。
1.2.2光合参数的测定 选择健康叶片的中上部并做标记,采用英国PP-Systems公司产CIRAS-1型便携式光合测定仪测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(C1)。测定时设置内置光源光合有效辐射为500μmol/(m2·s),CO2浓度为400μmol/mol,温度为25℃,每日9:00-11:00进行测定。
1.2.3叶绿素荧光参数的测定 采用英国Hansatech公司生产的FMS-2型便携式调制荧光仪测定各项荧光参数,测定时间、叶片部位与光合作用相同。测定前先暗适应30min,然后照射检验光[<0.05μmol/(m2·s)]测定叶片的初始荧光(F0),再用饱和白光[7200μmol/(m2·s)]照1s测最大荧光(Fm)及光适应[500μmol/(m2·s)]下的稳态荧光(Fs)、初始荧光(F0')、最大荧光(Fm')。根据仪器测定的参数值和Adams等和Schreiber等的方法,结合式(1)~(7)计算各参数。
1.3数据处理
采用Microsoft Excel进行数据处理和制图,采用DPS 7.05软件的LSD法进行差异显著性检验。
2 结果与分析
2.1ABA、PP333处理在低温胁迫及恢复中对叶绿素含量的影响
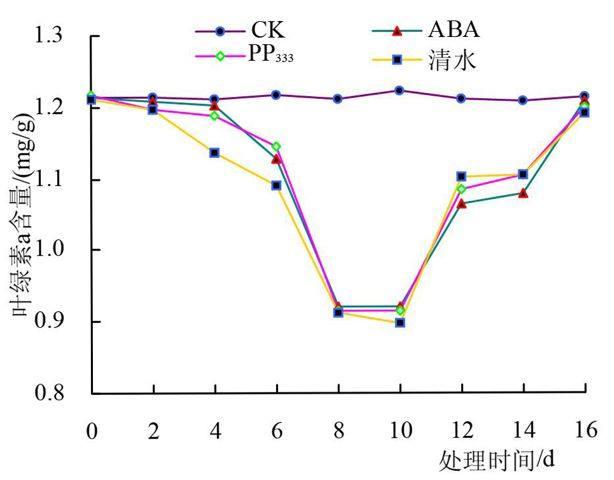
叶绿素是植物进行光合作用的主要色素,其含量的多少直接影响植物的生长。在本试验中,‘宋梅幼苗的叶绿素a含量、叶绿素b含量、叶绿素总量均随低温胁迫时间的延长而降低(图1)。低温胁迫下,各处理间叶绿素含量下降趋势不同,经ABA和PP333处理的幼苗叶绿素含量缓慢降低,而清水处理的幼苗叶绿素含量急剧下降,说明此低温下清水处理的幼苗受到的伤害较为严重。ABA处理和PP333处理比较,在胁迫初期(3~5天),经ABA处理的叶片叶绿素含量高于经PP333处理的叶片叶绿素含量,随胁迫时间的进一步延长(5~9天),经PP333处理的叶片叶绿素含量反而高于经ABA处理的叶片叶绿素含量,原因可能是不同植物激素对植物产生作用的条件不同。低温处理9天后,各处理的叶片叶绿素含量均降到了最低,叶绿素a含量分别降低了32.34%、32.83%、33.08%;叶绿素b含量分别降低了23.43%、23.65%、32.82%;叶绿素总量分别降低了29.33%、29.49%、32.99%。在恢复阶段,经ABA和PP333处理的植株叶片叶绿素含量恢复速度明显高于清水处理的叶片,在处理后的第21天(即恢复处理第12天),经ABA处理的叶片叶绿素a、b、a+b含量分别恢复到了对照的91.41%、86.19%、89.66%;经PP333处理的叶片叶绿素a、b、a+b含量分别恢复到了对照的86.19%、84.22%、77.46%清水处理的叶片叶绿素a、b、a+b含量分别恢复到了对照的89.66%、88.57%、85.75%。可见,经ABA和PP333处理后植株受到的低温伤害要小于清水处理,说明ABA和PP333处理可以提高植株的抗寒性。
2.2ABA、PP333处理在低温胁迫及恢复中对光合参数的影响
净光合速率是植物光合系统工作正常与否的重要指标。低温胁迫处理均降低了‘宋梅叶片的净光合速率(Pn),其中低温处理9天后的净光合速率降低最明显(图2A),ABA、PP333和清水处理的植株净光合速率与对照相比分别降低了53.12%、59.37%、62.5%,说明低温胁迫对春兰的光合系统功能产生了严重破坏。进入恢复期后净光合速率逐渐恢复,其中PP333处理的植株恢复最快,恢复12天后为对照的68.57%,而ABA和清水处理恢复到了对照的62.85%和54.28%。叶片的气孔导度和细胞间隙CO2浓度是影响光合速率的重要因素。本试验中,‘宋梅幼苗的气孔导度(Gs)随处理温度的降低而下降(图2B),其中低温处理9天后降幅最大,ABA、PP333和清水处理的植株气孔导度与对照相比分别降低了60.31%、61.91%、65.08%。而各处理的细胞间隙CO2浓度(Ci,图2C)都逐渐上升,与气孔限制值(Ls,图2D)呈相反的变化趋势。清水处理植株的Pn、Gs和Ls始终低于ABA和PP333处理,且均远低于对照。转入恢复条件后,各处理的Pn、Gs和Ls均开始回升,而Gi则降低。说明ABA和PP333处理对增强低温胁迫下‘宋梅的光合作用具有重要作用。
2.3ABA、PP333处理在低温胁迫及恢复中对叶绿素荧光参数的影响
初始荧光(F0)是PS Ⅱ反应中心处于完全开放时的荧光产量,它反映了PS Ⅱ天线色素受激发后的电子密度,与叶片的叶绿素浓度有关。Fv/Fm是PSⅡ最大光化学效率,其变化程度可以用来鉴别植物抵抗逆境胁迫的能力。在本试验中,各处理植株F0随着低温胁迫时间的延长而上升,Fv/Fm则逐步递减(图3)。ABA和PP333处理的植株在处理和恢复过程中F0和Fv/Fm波动较小,而清水处理的植株的F0则明显上升,Fv/Fm显著下降,且随着胁迫时间的延长,相邻处理时间之间升高或降低的幅度越大,低温胁迫第9天时F0较对照升高了44.15%,Fv/Fm较对照降低了55.24%,ABA和PP333处理的植株F0较对照分别升高了29.87%和11.95%,Fv/Fm较对照分别降低了11.95%和9.51%。表明低温对PSⅡ反应中心造成了伤害,使其光能转化能力显著下降,导致了春兰幼苗叶片光合能力的降低,而经ABA和PP333处理的植株受伤害程度低于清水处理的植株,说明ABA和PP333处理对提高春兰幼苗的抗寒能力具有一定的作用。在温度恢复阶段,恢复9天后基本接近对照水平,说明该伤害可在一定时间能得到修复,属于可逆损害。
春兰叶片的qP、φPSⅡ和Fv'/Fm'均随低温胁迫时间的延长显著降低。处理9天时,ABA、PP333和清水处理的植株qP与对照相比分别降低了53.7%、50%和70.37%,φPSⅡ与对照相比分别降低了82.75%、75.86%和89.65%,Fv'/Fm'与对照相比分别降低了58.49%、50.94%和69.81%。ABA和PP333处理植株的qP、φPSⅡ和Fv'/Fm'的下降幅度始终小于清水处理植株的下降幅度,且恢复处理后迅速恢复到对照水平,恢复12天后ABA和PP333处理基本恢复到了对照水平,而清水处理仍显著低于对照,说明低温胁迫对经ABA和PP333处理的植株未造成严重伤害,而导致了未处理的植株光合机构的部分失活或破坏。
低温胁迫处理显著降低了春兰叶片的光合电子传递速率(ETR)和光化学耗散能量(Prate),提高了天线热耗散速率(Drate)。处理9天时,ABA、PP333和清水处理的植株ETR分别降低了81.93%、75.98%和91.11%,ABA和PP333处理的植株在胁迫初期下降幅度较小,第3天开始才明显下降,恢复13天后达到对照水平的78.21%和79.41%,而此时清水处理的植株仅恢复到对照水平的55.09%,说明低温胁迫对未作处理的植株造成的伤害较大,导致了部分光合机构的失活或破坏。而处理9天时,ABA、PP333和清水处理的植株Prate分别为对照的18.61%、24.01%和8.32%,Drate分别比对照升高了65.95%、57.44%和78.72%。恢复处理后,各处理Prate仍低于对照水平,说明低温胁迫已对植物的光合机构造成了破坏,ABA和PP333处理的植株相比清水处理的植株受伤害较轻,说明ABA和PP333处理对提高春兰的抗寒性具有一定作用。
3 结论与讨论
低温胁迫使植物产生大量的自由基,损伤膜系统,最终造成低温伤害。应用植物生长调节剂改善植物抗寒性,保护植物免受低温伤害,是简便易行、经济适应的一种方法。植物体内的保护系统有2类:一是酶促系统,包括过氧化物酶(POD)、过氧化氢酶(CAT)等;二是非酶促系统,包括游离脯氨酸(Pro)、可溶性糖等。因此,对抗寒性的评价必须是多方面的。植物生长调节剂在提高植物抗寒性方面多有报道,但多集中于大田作物,对提高观赏植物抗寒性方面未见报道。
光合作用是植物受低温影响最明显的生理过程之一。当春兰幼苗受到低温胁迫时,叶绿体结构受到严重破坏。本试验中,低温胁迫处理9天后,ABA、PP333和清水处理的植株叶片叶绿素a含量分别降低了32.34%、32.83%、33.08%;叶绿素b含量分别降低了23.43%、23.65%、32.82%;叶绿素总量分别降低了29.33%、29.49%、32.99%。可见,低温胁迫造成了叶绿素含量的普遍降低,但清水处理的幼苗叶绿素含量始终低于经ABA和PP333处理的幼苗,说明ABA和PP333对保护叶绿体结构起到了一定作用。
通常认为逆境胁迫导致植物净光合速率(Pn)下降的原因主要包括气孔因素和非气孔因素。本研究中,随着低温胁迫时间的延长,春兰叶片净光合速率(Pn)、气孔导度(Gs)和气孔限制值(Ls)均明显降低,细胞间隙CO2浓度(C1)上升,说明低温胁迫下春兰叶片净光合速率的降低不是由于气孔导度下降导致的CO2供应量减少,而是由于非气孔因素阻碍了CO2的利用,导致了细胞间隙CO2的积累。
低温环境中的叶片受光可导致光合细胞器的受损,这种现象就是光合作用中的光抑制。光抑制启动了活性氧的诱导,这个过程可能会导致光合系统的退化,类囊体膜的膜脂过氧化,以及碳代谢过程中酶的失活。大量研究表明,植物体内的ABA在低温下所起的作用很重要,Chen等研究后提出假设,冷驯化是由ABA的作用启动的,外源施用ABA可以增加某些经过冷驯化植物种类的抗寒性。
低温胁迫对植物光合作用的影响是多方面的,叶绿素荧光测定技术被认为是探知植物体内生理状态与环境变化的理想探针,通过荧光变化可以探测光合作用和热耗散情况。F0是经暗适应的光合机构PS Ⅱ反应中心完全开放时的荧光强度,它与激发光的强度和叶绿素浓度有光,而与光合作用的光反应无关,Fv/Fm反映了PSⅡ反应中心的最大光能转换效率,在非胁迫条件下,Fv/Fm的变化极小,但在逆境条件下,植物受到胁迫或其他原因猝灭了荧光,这一效率值则会明显下降。在本试验中,随着低温胁迫时间的延长,各处理均造成了F0的升高和Fv/Fm的降低,原因应该是低温使植物受到了光抑制且导致了PS Ⅱ反应中心的失活或破坏。而经ABA和PP333处理的植株F0和Fv/Fm变化不大,这表明此时的低温胁迫并未对其造成明显的光抑制,而清水处理的植株F0和Fv/Fm变化较大,原因可能是低温严重抑制了植物叶片的碳同化能力,并使光能过剩,导致低温光抑制,这与Fracheboud在抗寒小麦品种中的研究相似。在恢复阶段,ABA和PP333处理植株的F0和Fv/Fm在第21天基本恢复到了对照水平,可见此条件并未造成PS Ⅱ反应中心的损害,只是低温下的可逆性失活。Fv'/Fm'在低温胁迫阶段一直处于下降趋势,说明低温处理下兰花叶片的PS Ⅱ反应中心随胁迫时间的延长而逐渐关闭,天线色素光能传递效率下降,这可能是植物为防止过剩光能破坏光合机构而形成的一种保护性机制。qP反映的是光能中用于光化学反应的部分,反映了光合活性的高低,φPSⅡ反映了光合电子传递量子效率的强弱。本试验中,各处理的Fv'/Fm'、qP、φPSⅡ和ETR均随低温胁迫时间的延长呈下降趋势,说明低温造成了PSⅡ活性中心的潜在活性和原初光能转换效率的减弱,使光合电子传递受阻,抑制为光合碳同化提供ATP和NADPH,引起碳同化降低,从而影响了兰花正常的光合作用。但经ABA和PP333处理的植株下降幅度较低,保持了较高的Fv'/Fm'、qP、φPSⅡ和ETR值,说明ABA和PPP333处理提高了春兰低温下对光抑制的抗性,这与刘鹏在冷锻炼甜椒品种叶绿素荧光中的研究结果相似,可以说ABA和PP333处理相当于对春兰进行了冷锻炼。本试验观察到,低温下春兰分配于光化学反应的能量减少,分配于天线耗散的能量增加,说明低温胁迫下春兰光合速率下降的同时,光合机构的过剩激发能增多,在光化学反应耗能减少的情况下,天线系统的热耗散增加,但经ABA和PP333处理的春兰光反应中心在低温下受到的伤害小,光化学效率高,分配于天线系统的能量耗散少。
植物的抗寒性在生理生化方面的表现,最终由植物体基因表达来控制,所以基因方面的研究有利于从根本上改善植物的抗寒性。春兰幼苗叶片PS Ⅱ反应中心是否存在不可逆失活,哪些植物生长调节剂能够启动植物的冷驯化,如何对环境低温做出应答以及植物周围的环境信号对植物抗寒性的影响,都是今后植物抗寒性的热点,有待于进一步研究。
猜你喜欢
快乐作文(1.2年级)(2023年2期)2023-03-30 01:09:26
健康体检与管理(2022年4期)2022-05-13 18:42:17
山东农业科学(2017年2期)2017-03-15 18:55:09
新疆农垦科技(2016年2期)2016-08-21 13:50:26
现代园艺(2016年2期)2016-03-15 16:05:02
中国果菜(2016年9期)2016-03-01 01:28:40
江苏农业科学(2015年11期)2016-01-27 14:59:30
中国马铃薯(2015年5期)2016-01-09 06:11:11
江苏农业科学(2015年8期)2015-09-10 16:07:34
小小说月刊·下半月(2015年2期)2015-05-14 15:23:17
