
小麦结实小穗数QTL分析
2014-04-29 00:44马昊翔董凡凡梁增浩王曙光王绘艳景蕊莲黛珍
农学学报 2014年4期
关键词:小麦
马昊翔 董凡凡 梁增浩 王曙光 王绘艳 景蕊莲 黛珍
摘要:为利用分子标记辅助选择技术选育抗旱高产小麦品种提供基础,以小麦‘旱选10号/‘鲁麦14DH群体为材料,在干旱胁迫及正常灌溉两种水分条件下于2010年和2011年考察小麦主茎结实小穗数,通过采用基于混合线性模型的复合区间作图法分析其QTL,研究控制小麦结实小穗数的数量性状基因。共检测到7个加性QTLs和2对上位性QTLs。加性QTL的LOD值介于2.56~5.08之间,分别位于2D、4A、6A和6B染色体上,对表型变异的贡献率为7.19%-13.65%;上位性QTL的LOD值分别为5.24和5.47,分别位于3A和6B与4B和5B染色体上,对表型贡献率分别为13.71%和17.93%。其中OFns-6A-2于2010年正常灌溉和2011年2种水分条件下均被检测到,可用于分子标记辅助育种。
关键词:小麦;结实小穗数;DH群体;QTL
中图分类号:S512.1+1
文献标志码:A
论文编号:2013-0715
0 引言
小麦是中国的主要粮食作物之一,提高小麦产量是保障中国粮食安全的重要途径。提高结实小穗数可增加穗粒数,从而改善产量构成因素。蒲定福等的研究表明,结实小穗数的遗传较大程度偏离加性-显性效应模型,且存在其他效应。海燕等认为,结实小穗数是由多对基因控制的数量性状,遗传基础复杂。目前,DNA分子标记和遗传连锁图谱已广泛应用于数量性状的研究。控制小麦小穗数的QTL已被定位于除2D、3D、4B之外的所有染色体上。对于结实小穗数QTL,不同研究者将其定位在不同染色体上。杨睿等利用波兰小麦×普通小麦品系中13RIL群体将有效小穗数QTL定位于3B染色体上。张国华等以128份黄淮麦区小麦品种(系)为材料检测到5个与可育小穗数显著关联的QTL,分布在1B、2A、4B和7A染色体上。张坤普等利用小麦花培3号/豫麦57DH群体将可育小穗数QTL定位在4A和5D染色体上。Li等利用‘Chuan 35050בShannong 483的RIL群体将结实小穗数QTL定位在2A、5D、6B和7D染色体上。Ma等利用136个‘Nanda2419בWangshuibaiRIL家系重排列互交产生的永久F2群体将结实小穗数QTL定位在1A、2D、3B、6A、7A和7D染色体上。但目前关于干旱胁迫环境条件下小麦结实小穗数QTL的报道较少。鉴于此,本研究对干旱胁迫与正常灌溉2种水分条件下的结实小穗数QTL进行定位分析,检测与之紧密连锁的分子标记,为利用分子标记辅助选择技术选育抗旱高产小麦品种提供基础。
1 材料与方法
1.1试验材料
供试材料为‘旱选10号/‘鲁麦14杂种F1花药离体培养创建的一个具有150个家系的DH群体,群体内各家系间变异广泛。该群体的2个亲本‘旱选10号与‘鲁麦14在抗旱性上的遗传差异较大,母本‘旱选10号(晋麦5号)是优异抗旱资源;父本‘鲁麦14是山东省烟台市农科所育成的水地高产品种。
1.2试验设计
DH群体及2个亲本于2010年及2011年分别种植于山西农业大学农作站,行长2m,行距25cm,每行均匀播60粒种子。试验采用裂区设计,3次重复。水分条件分为干旱胁迫(drought-stress,DS)和正常灌溉(well-watered,WW)。播前统一浇足底墒水,播种后干旱胁迫的材料全生育期只依靠自然降水,2010年与2011年小麦整个生育期降雨量分别为100mm和110mm;正常灌溉的材料在越冬前、拔节期、抽穗期、灌浆期分别灌溉,灌溉量650m3/(hm2·次)。
1.3测定内容及统计分析
成熟后每个家系及亲本从小区中部随机选取10株考种,记录主茎结实小穗数(fertile spikelet number,FSN)。利用SPSS V20软件对小麦DH群体各家系以及亲本的主茎结实小穗数进行数据统计分析。
1.4QTL分析
QTL定位分析采用基于混合线性模型的复合区间作图法的QTL作图软件QTL IciMapping V3.2,检测小麦主茎结实小穗数在干旱胁迫与正常灌溉2种条件下的加性QTL、加性×加性上位性QTL。以LOD≥2.5作为阈值判断QTL的存在与否,加性效应和上位性互作效应QTL分析以P<0.005、P<0.001为显著水平进行显著性检验。QTL的命名按照http://www.graingenes.org的方法进行,即“QTL+性状名称缩写+染色体编号”。
2 结果与分析
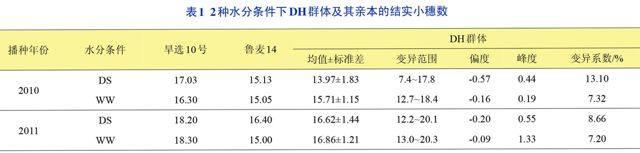
2.1DH群体及亲本间的表型差异
DH群体及亲本间的结实小穗数见表1。2年两种水分条件下母本‘旱选10号的结实小穗数均显著大于父本‘鲁麦14,表明2个亲本的遗传基础存在较大差异。DH群体结实小穗数2010年干旱胁迫下的平均值小于亲本,其余条件下均介于两亲本之间。不同环境条件下,小麦DH群体各家系的结实小穗数均表现为超亲分离,各家系之间存在明显的遗传差异,分布范围广泛,表现为连续的正态分布(图1),符合QTL定位的要求。
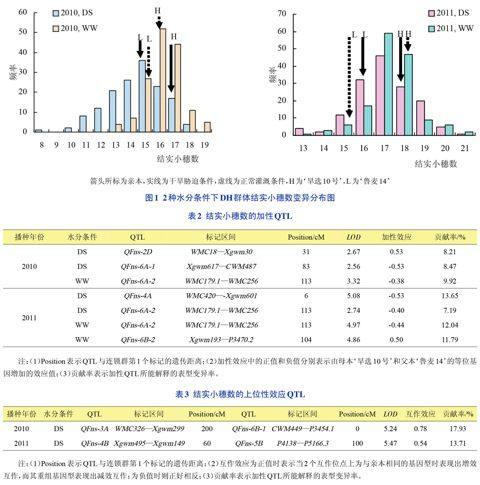
2.2结实小穗数OTL分析
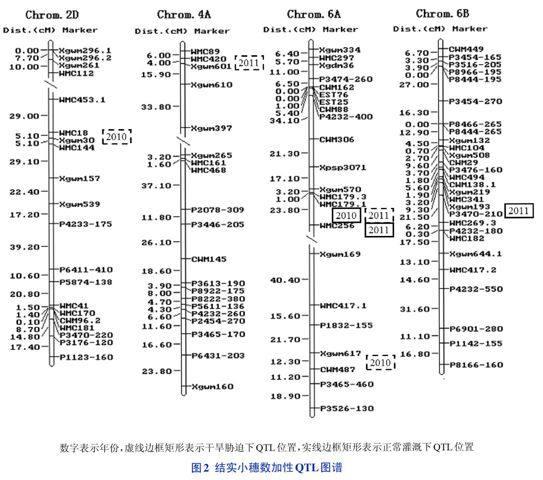
2年2种水分条件下,共检测到7个加性QTLs(表2和图2),分别位于2D、4A、6A和6B染色体上,LOD值介于2.56~5.42之间,对表型变异的贡献率为7.19%~13.65%。其中位于标记区间WMC179.1-WMC256上的OFns-6A-2在2010年正常灌溉及2011年2种水分条件下均被检测到,增效等位基因均来自父本‘鲁麦14,为稳定表达的QTL。而QFns-2D(2010)、QFns-6A-1(2010)和QFns-4A(2011)仅在干旱胁迫下检测到,QFns-6B-2仅在2011年正常灌溉下检测到,表明这些QTLs受环境影响较大。
共检测到2对上位性QTLs(表3)。2010年干旱胁迫下检测到的QFns-3A和QFns-6B-1为A和B基因组间的互作,互作效应值为0.78,表现为正效应,即重组型上位性效应小于亲本型上位性效应,对表型的贡献率为17.93%;2011年干旱胁迫下检测到的QFns-4B和QFns-5B为B基因组内的互作,互作效应值为0.54,表现为正效应,即重组型上位性效应小于亲本型上位性效应,对表型的贡献率为13.71%。不同年份正常灌溉条件下均未检测到结实小穗数的上位性QTLs。
3 讨论
数量性状受多基因控制,遗传基础复杂,且易受环境影响,因此只有在多种环境条件或多个年份重复鉴定才能探知目的性状的环境钝感QTL,从而用于分子标记辅助选择育种。本研究利用‘旱选10号/鲁麦14DH群体,2年2种水分条件下,共检测到7个加性QTLs。其中位于6A染色体标记区间WMC179.1-WMC256的QFns-6A-2在2010年正常灌溉及2011年2种水分条件下均被检测到,为稳定表达的QTL,可用于分子标记辅助选择育种。
利用相同的群体,An等于低氮条件下在6A染色体上相同标记区间检测到一个氮吸收量QTL,Su等检测到一个磷吸收量QTL。此外,Zhang等利用硬粒‘小麦UC1113בKofa的RIL群体在相同标记区间检测到一个灰分含量QTL。另一方面,本研究检测到的QFns-2D与刘秀林等利用相同群体检测到的控制根直径的QRD.cgb-2D以及Su等检测到的旗叶greenness(叶绿素含量)QTL均位于2D染色体标记区间WMC18-Xgwm30,表明这些QTLs(基因)或一因多效,或紧密连锁。Yang等利用相同群体在相邻标记区间WMC453.1-WMC18定位了1个茎可溶性糖QTL和1个成熟期千粒重QTL。这2个标记区间可能形成了根部性状、产量性状、氮磷吸收利用率、持绿性状QTL热点区域,在这些区域内发掘可靠的分子标记,一次可将控制多个优良性状的基因转移至同一品种中,从而大大加快育种改良进程,提高改良的效率。
基于Somers等的小麦遗传连锁图谱,比较本研究定位的QTLs与先前定位的小麦结实小穗数QTL研究结果,发现一些QTLs位于相近的位置。例如,在6B染色体上检测到的QFns-6B-2与Li等利用‘Chuan 35050בShannong 483的RIL群体检测到的QFss.sdau-6B.e4相距约1cM,可能为同一基因。在2D染色体上检测到的QFns-2D与Ma等利用136个‘Nanda2419בWangshuibaiRIL家系重排列互交产生的永久F2群体检测到的2个QTLs相距21~30cM;在4A染色体上检测到的QFns-4A与张坤普等检测到的qFsps4A相距约50cM,表明2D和4A染色体上确实存在结实小穗数QTL。
对于本研究检测到的稳定表达的QTL以及QTL热点区域,通过加密目标区间的连锁图谱或利用更大的作图群体可实现结实小穗数QTL的精细定位和区分热点区域QTL的一因多效性或紧密连锁性,为图位克隆提供基础。
4 结论
本研究在2年2种水分条件下,共检测到7个加性小麦结实小穗数QTLs,其中QFns-6A-2在2010年正常灌溉及2011年2种水分条件下均被检测到,增效等位基因均来自父本‘鲁麦14,为稳定表达的QTL,连锁的分子标记可用于辅助选择育种。其余4个QTL,包括QFns-2D、QFns-6A-1、QFns-4A和QFns-6B-2仅在1年1种水分环境条件下检测到,受环境影响较大。
猜你喜欢
今日农业(2022年16期)2022-11-09
今日农业(2021年12期)2021-11-28
今日农业(2021年8期)2021-11-28
今日农业(2021年8期)2021-11-28
金桥(2021年10期)2021-11-05
今日农业(2021年13期)2021-08-14
河北农机(2020年10期)2020-12-14
今日农业(2020年13期)2020-08-24
作文小学中年级(2020年4期)2020-06-11
今日农业(2019年11期)2019-08-13
