
三疣梭子蟹(Portunus trituberculatus)胞内氯离子通道蛋白基因克隆及其表达分析*
2014-03-19 12:17:39吕建建高保全
海洋与湖沼 2014年6期
王 渝 吕建建 刘 萍① 高保全 李 健 陈 萍
(1. 农业部海洋渔业可持续发展重点实验室 中国水产科学研究院黄海水产研究所 青岛 266071;2. 上海海洋大学水产与生命学院 上海 201306)
三疣梭子蟹(Portunus trituberculatus), 俗称梭子蟹, 隶属于甲壳纲(Crustacea)、十足目(Decapoda)、梭子蟹科(Portunidae)、梭子蟹属(Portunus), 是中国沿海的重要经济蟹类, 其可以存活于盐度为13.7—47.7的水环境中(隋延鸣等, 2012), 属典型广盐性蟹类(孙颖民等, 1984)。在三疣梭子蟹的育苗过程中, 盐度微小的变化都可能导致变态的推迟、活力变差, 乃至病害的发生(季东升, 2005)。在三疣梭子蟹养成过程盐度也起到了重要作用, 对其存活、摄饵和生长均具有重要影响(吕富等, 2010; 路允良等, 2012)。然而, 在三疣梭子蟹养殖过程中, 降雨会改变水体的盐度, 因此养殖过程中产生的盐度胁迫是不可避免的。一般认为造成盐度胁迫的主要物质是NaCl, 在其胁迫下三疣梭子蟹体内的Cl–含量会超出正常需要, 进而对机体产生胁迫作用(康敏华等, 2011)。然而, 目前在三疣梭子蟹中渗透压相关的研究主要集中在酶活、免疫、蜕皮和能量收支等方面(郑萍萍等, 2010; 江山等, 2011),涉及Cl–的渗透压调节分子机理的研究则相对较少。因此, 从分子水平研究三疣梭子蟹氯离子通道对Cl–运输、吸收和排除显得尤为重要(康敏华等, 2011)。
氯离子通道是分布于细胞膜或细胞器质膜上的一类能够转运氯离子及其它阴离子的通道蛋白(陈丽娥等, 2010)。研究表明, 氯离子通道可调控不同的生理过程和细胞功能, 如: Cl–运输、渗透压调节、离子稳态、胞内pH、细胞容积调节、电兴奋性、金属耐性和信号识别与转导(Uchidaet al, 1993; Adachiet al,1994; Hechenbergeret al, 1996), 甚至在细胞迁移、细胞增殖和分化等过程中也起到一定作用(Jentschet al,1999)。氯离子通道分为三种类型: 氯通道(chloride channel, CLC)、囊性纤维化跨膜传导调解因子(cystic fibrosis transmembrane conductance regulator, CFTR)和胞内氯离子通道(chloride intracellular channel,CLIC) (戴寒晶, 2006)。CLIC家族成员大多数为单次跨膜蛋白, 与谷胱甘肽-S-转移酶(glutathione-S-transferase, GST)具有一定的同源性(Dulhuntyet al,2001)。Valenzuela等(1997)从人类单核细胞中克隆了第一个CLIC1基因。现在已经发现了六个CLIC家族成员, 包括CLIC1 (Valenzuelaet al, 1997)、CLIC2(Heisset al, 1997)、CLIC3 (Qianet al, 1999)、CLIC4(Edwards, 1999)、CLIC5 (Berrymanet al, 2000)和CLIC6 (Perluigiet al, 2002), 各成员之间高度保守。Berry等(2003)在线虫中发现了编码一种CLIC样蛋白的基因EXC-4。目前关于氯离子通道蛋白的研究主要集中在哺乳动物、两栖类、昆虫和植物(Cornejoet al,2001; Shorninget al, 2003; Donget al, 2013), 在水产动物中研究较少, 而在甲壳动物中更是尚未见报道。
本研究从本实验室构建的三疣梭子蟹转录组文库(Lvet al, 2013)中筛选到胞内氯离子通道蛋白基因EST序列; 采用RACE技术克隆获得该基因全长cDNA序列; 借助相关生物信息学软件分析PtCLIC基因及其翻译的氨基酸序列的结构; 通过Blast程序比对其与其它物种的CLIC基因氨基酸序列同源性;利用实时荧光定量技术研究其组织表达模式、低盐胁迫进程中的表达规律以及低盐胁迫下低盐耐受和低盐敏感家系中表达规律的差异。本研究为PtCLIC基因的生物学功能研究及其在渗透压调节机理方面的研究奠定理论基础, 同时也为三疣梭子蟹耐盐新品系的选育提供理论指导。
1 材料与方法
1.1 材料
健康三疣梭子蟹取自昌邑市海丰水产养殖有限公司, 体质量(5.78±1.11)g。暂养于3m3的室内水泥池中, 每池90只, 共6池, 暂养3d。养殖水温26°C, 盐度33, pH 8.7, 持续充氧, 养殖用水每天更换1/3, 于18:00定时喂食蓝蛤。
Trizol Reagent购自Invitrogen公司; SMARTTMRACE Amplification Kit和Advantage 2 PCR Kit购自Clontech公司; DNA胶回收试剂盒购自生工生物工程(上海)股份有限公司; SYBR Premix Ex Taq Ⅱ荧光定量试剂盒、PrimeScript RT reagent Kit、PMD18-T载体和DH5α感受态细胞均购自TaKaRa公司。
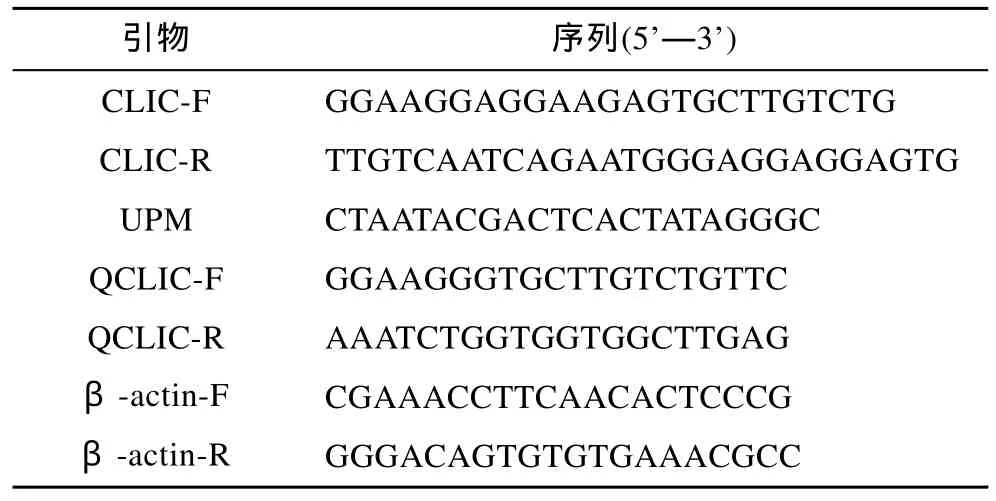
表1 实验所用的引物序列Tab.1 All sequence of primers for the experiment
1.2 总RNA的提取及cDNA第一链的合成
Trizol试剂分别提取鳃、肝胰腺和肌肉组织的总RNA, 核酸定量仪与1.0%的琼脂糖凝胶电泳检测RNA的质量及完整性; 分别取等量的上述3种组织的总RNA均匀混合, 按照SMARTTM RACE Amplification Kit说明书上的方法分别合成3’和5’RACE的cDNA第一链。
1.3 PtCLIC基因全长cDNA的克隆及测序
3’RACE和5’RACE特异性引物利用从三疣梭子蟹转录组文库中获得的一段胞内氯离子通道蛋白基因EST序列和Primer Premier 5.0软件设计, 并且委托生工生物工程(上海)股份有限公司合成。3’和5’末端的快速扩增使用试剂SMARTTMRACE Amplification Kit, Advantage 2 PCR Kit和两对特异性引物对CLIC-F和UPM、CLIC-R和UPM进行(表1)。反应程序: 94°C 30s, 68°C 30s, 72°C 3min, 30个循环。
利用1.0%的琼脂糖凝胶电泳分别检测3’RACE和5’RACE的扩增产物, 使用胶回收试剂盒回收目的片段, 并连接入PMD18-T载体, 转化入大肠杆菌DH5α感受态细胞, 挑取阳性克隆进行菌落PCR鉴定, 将含有目的条带的菌液送往上海桑尼生物科技有限公司进行测序。
1.4 PtCLIC基因的生物信息学分析
利用Vector NTI 11.0软件去除所测序列中的载体, 并与已知序列拼接。利用DNAStar的EditSeq程序预测开放阅读框(ORF)并将其翻译为氨基酸序列。使用BLAST (http://www.blast.ncbi.nlm.nih.gov/Blast.cgi)程序分别进行PtCLIC基因的核苷酸序列和推导的氨基酸序列同源性比对。利用ProtParam tool软件进行蛋白质理化性质预测, 利用InterproScan软件进行蛋白质功能结构域分析, 利用TMHMM 2.0软件进行蛋白质跨膜结构域分析。使用Clustal X软件对三疣梭子蟹PtCLIC氨基酸序列与其它物种的CLIC氨基酸序列进行多序列比对, 然后使用MEGA 4.0软件,采用邻接法构建系统进化树。
1.5 盐度胁迫实验
随机挑取暂养3d的三疣梭子蟹分为2组, 对照组(33)和低盐组(11), 每个处理组设3个平行, 每个平行90只蟹。使用地下卤水、自然海水和淡水配制各盐度实验用水, 使用YSI盐度仪进行盐度校准, 具体方法见隋延鸣等(2012)。各组分别于胁迫0、3、6、12、24、48和72h取鳃和肝胰腺, 放置液氮中冷冻保存, 用于后续RNA的提取, 每个时间点取3只。另取3只健康三疣梭子蟹的鳃、肝胰腺、肌肉、心脏、眼柄和血细胞, 放置液氮中冷冻保存, 用于后续RNA的提取。
1.6 家系低盐胁迫实验
从本实验室构建的低盐耐受家系和低盐敏感家系中随机选取三疣梭子蟹各90只, 胁迫盐度为三疣梭子蟹半致死浓度(11), 每个家系设置3个平行, 每个平行30只。各组分别于胁迫0h、12h、72h取鳃组织, 放置液氮中冷冻保存, 用于后续RNA提取。
1.7 PtCLIC基因表达分析
使用Trizol提取不同实验组三疣梭子蟹鳃和肝胰腺组织的总RNA, 使用PrimeScript RT reagent Kit反转录合成cDNA。
根据已获得的三疣梭子蟹内参基因β-actin和PtCLIC基因全长序列, 分别设计两对引物(β-actin-F和β-actin-R、QCLIC-F和QCLIC-R)(表1), 对不同时间点盐度胁迫的三疣梭子蟹鳃和肝胰腺中PtCLIC基因的相对表达量进行检测。使用SYBR Premix Ex TaqⅡ试剂在ABI 7500 Real Time PCR仪上利用Real-time PCR方法对各个组织中以及胁迫进程中PtCLIC基因的表达情况进行分析。荧光定量PCR反应体系为20μL, 包括10μL SYBR Premix Ex TaqTMⅡ(2×), 0.8μL 10μmol/L的引物QCLIC-F, 0.8μL 10μmol/L的引物QCLIC-R, 0.4μL ROX Reference DyeⅡ(50×), 2.0μL cDNA模板, 6.0μL无菌ddH2O。反应程序为: 95°C 30s; 95°C 5s, 60°C 34s, 40个循环;95°C 15s, 60°C 1min, 95°C 15s。采用2–△△CT方法计算PtCLIC基因的相对表达量, 使用SPSS 17.0软件进行单因素方差分析。
2 结果
2.1 总RNA提取
利用Trizol试剂提取获得三疣梭子蟹鳃、肝胰腺和肌肉组织的总RNA, 经核酸定量仪检测, 其OD260/OD280均在1.9—2.0之间, 表明总RNA纯度较高; 经1.0%琼脂糖凝胶电泳检测, 28S、18S、5S rRNA条带清晰, 表明总RNA完整性较好, 质量较高。
2.2 PtCLIC基因的全长cDNA克隆
以三疣梭子蟹的鳃、肝胰腺和肌肉组织的等量总RNA混合物为模板, 反转录得到3’RACE和5’RACE的cDNA第一链。以特异引物CLIC-F和CLIC-R分别与通用引物UPM配对, 进行3’和5’RACE扩增。将测序获得的3’和5’片段与已知EST序列进行拼接,得到三疣梭子蟹CLIC基因的全长cDNA序列, 命名为PtCLIC。该基因全长为2000 bp (GenBank登录号为KJ186099), 包括76 bp的5’端非编码区(UTR),1162 bp的3’端非编码区和762 bp的开放阅读框(ORF)。3’端含有一个PolyA尾和一个多聚腺苷酸加尾信号(AATAAA) (图1)。
2.3 PtCLIC基因序列分析及结构域预测
DNAstar软件分析表明,PtCLIC基因编码一个由253个氨基酸组成的蛋白质, 分子量为29.2kDa, 理论等电点为5.93。ProtParam tool软件分析表明,PtCLIC蛋白带有负电的氨基酸残基为36个(Asp和Glu), 带有正电的氨基酸残基为31个(Arg和Lys), 不稳定系数为46.34, 亲水性平均数为-0.449, 属于不稳定蛋白。InterProScan和TMHMM 2.0软件分析表明,PtCLIC基因翻译的蛋白质结构中无跨膜结构域。
2.4 PtCLIC基因的同源性分析
利用BLAST程序对三疣梭子蟹PtCLIC基因的氨基酸序列进行同源性比对, 发现三疣梭子蟹PtCLIC基因与蚤状溞(Daphnia pulex)、点蜂缘蝽(Riptortus pedestris)、地中海实蝇(Ceratitis capitata)、埃及伊蚊(Aedes aegypti)和果蝇(Drosophila virilis)的同源性分别为83%、84%、82%、83%和82%。通过与上述物种的CLIC氨基酸序列比对发现,PtCLIC氨基酸序列保守性较高(图2)。

图1 三疣梭子蟹PtCLIC基因cDNA全长及其编码的氨基酸序列Fig.1 Complete cDNA sequence and deduced amino acids sequence of P. trituberculatus PtCLIC gene
利用MEGA 4.0软件进行系统进化分析表明, 所有物种的CLIC共聚为两大类群: 脊椎动物和无脊椎动物。在无脊椎动物中, 三疣梭子蟹与拟穴青蟹紧密聚为一支, 然后与埃及伊蚊、蚤状、捕食螨聚为一大支; 在脊椎动物中, 野猪与白眉翁聚为一支(图3)。
2.5 PtCLIC基因表达分析
2.5.1PtCLIC基因的组织表达分析 采用实时荧光RT-PCR分析了三疣梭子蟹PtCLIC基因在不同组织中的表达分布特征。结果显示,PtCLIC基因在鳃、肝胰腺、肌肉、眼柄、血液和心脏中均有表达。其中,在肝胰腺中的表达量最高, 且显著高于其它组织(P<0.05)(图4)。
2.5.2PtCLIC基因在低盐度胁迫进程中的表达分析低盐胁迫后PtCLIC在鳃和肝胰腺中的相对表达量分别如图5、图6所示。在鳃中, 与对照组相比, 低盐胁迫0—6h后,PtCLIC出现显著下调现象, 6h的相对表达量为对照组的0.61倍(P<0.05), 12—48hPtCLIC的相对表达量出现显著上调, 并于48h达到最大值,相对表达量为对照组的4.18倍(P<0.05), 48—72h又出现显著下调趋势(图5)。在肝胰腺中, 与对照组相比, 低盐胁迫0—6h后,PtCLIC的相对表达量组间差异不显著, 6—24hPtCLIC的相对表达量出现显著上调, 并于24h达到最大值, 相对表达量为对照组的4.48倍(P<0.05), 24—72h又出现显著下调(图6)。
2.5.3PtCLIC基因在低盐耐受与低盐敏感家系中的表达分析 实时荧光定量RT-PCR结果显示, 在低盐耐受家系中, 0—12hPtCLIC基因相对表达量出现上调, 随后12—72h显著下调, 三个时间点低盐耐受家系的表达量分别为低盐敏感家系的2.31、2.53和1.90倍。低盐敏感家系0—72h之间PtCLIC基因相对表达量差异不显著, 保持初始水平几乎没有变化,且显著低于低盐耐受家系各时间点的相对表达量。见图7。

图2 三疣梭子蟹PtCLIC氨基酸序列与其它物种的CLIC氨基酸序列比对Fig.2 The amino acid sequence of P. trituberculatus PtCLIC alignment with other species’ CLIC
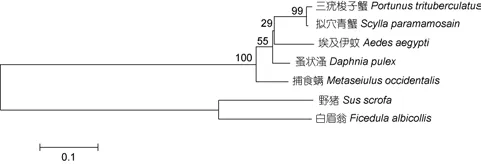
图3 利用MEGA 4.0软件NJ法构建的系统进化树Fig.3 NJ phylogenetic tree made by MEGA 4.0
3 讨论
CLIC家族成员在体内分布非常广泛, 主要存在于真核细胞质膜和各种细胞器上。研究发现(Heisset al, 1997; Wartonet al, 2002), 人类CLCI1和CLIC2分别编码241和243个氨基酸, 随后Qian等(1999)发现小鼠CLIC3编码208个氨基酸, Berryman等(2000)发现人类CLIC5编码251个氨基酸。本研究采用RACE技术首次克隆得到762 bp的PtCLIC基因cDNA开放阅读框, 预测蛋白质由253个氨基酸组成。蛋白质结构域分析表明, 在PtCLIC基因编码的氨基酸中不存在跨膜结构域, 且分子量较小, 推测该蛋白本身并不是氯通道, 而仅仅起到氯通道活化蛋白的功能(Valenzuelaet al, 1997)。另外, 在与PtCLIC同源关系较近的蚤状(Daphnia pulex)和地中海实蝇(Ceratitis capitata)等物种的CLIC氨基酸序列上亦不存在跨膜结构域, 这与本实验结果一致。同源性分析表明,PtCLIC与蚤状溞等物种CLIC的同源性均高于80%,确认该基因为三疣梭子蟹CLIC基因。多氨基酸序列比对分析表明,PtCLIC氨基酸序列保守性较高, 与NCBI数据库中氨基酸序列比对结果未能确认本研究所得PtCLIC基因具体亚型, 因此PtCLIC基因的分型工作有待于深入研究。系统进化分析表明,PtCLIC与同属的拟穴青蟹亲缘关系较近与野猪等脊椎动物的亲缘关系较远。验证了亲缘关系近的同源性高, 亲缘关系远的同源性低的规律, 符合遗传进化规律。
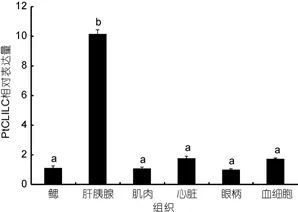
图4 三疣梭子蟹不同组织中PtCLIC基因的表达Fig.4 Expression of PtCLIC gene in different tissues of P.trituberculatus

图5 低盐胁迫下三疣梭子蟹PtCLIC基因在鳃中的表达情况Fig.5 Expression of PtCLIC gene in P. trituberculatus gill tissue under low salinity stress

图6 低盐胁迫下三疣梭子蟹PtCLIC基因在肝胰腺中的表达情况Fig.6 Expression of PtCLIC gene in P. trituberculatus hepatopancreas tissue under low salinity stress
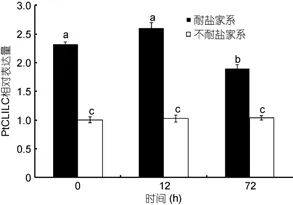
图7 低盐胁迫下低盐耐受家系和低盐敏感家系中PtCLIC基因在鳃中的表达情况Fig.7 Expression of PtCLIC gene in low salinity tolerant family and low salinity susceptive family in P. trituberculatus gill tissue under low salinity stress
实时荧光RT-PCR证实PtCLIC基因在检测的所有组织中均有表达。在肝胰腺中的表达量最高, 且显著高于其它组织中的表达量。这表明三疣梭子蟹PtCLIC基因的表达具有组织特异性。Liu等(2011)的研究表明, 肝脏是甲壳动物离子储存和代谢的中心,由此推测三疣梭子蟹的肝胰腺在氯离子转运中起到了重要作用。
目前关于甲壳类动物胞内氯离子通道蛋白对盐度胁迫的应答机制还未见报道。鳃是甲壳动物与外界环境进行气体和离子交换的介质(Freireet al, 2008),而肝脏又是离子储存和代谢的中心(Liuet al, 2011),因此研究低盐胁迫进程中PtCLIC在鳃和肝胰腺中的表达规律十分必要。Freire等(2008)研究了甲壳动物中NaCl的吸收和排出过程, 证实了Cl–的平衡是甲壳动物渗透压调节的关键。在Cl–的吸收过程中,首先Cl–与HCO3–交换通过鳃上皮顶部质膜, 继而在基底侧质膜上通过Cl–通道进入血淋巴中。而在Cl–的排出过程中, Cl–首先以Na+/K+/2Cl–形式协同转运通过基底侧质膜, 继而在鳃上皮顶部质膜上通过Cl–通道排到水环境中。本实验结果显示低盐胁迫下PtCLIC基因在鳃和肝胰腺中的表达总体呈现先上调然后下调的规律。低盐胁迫下PtCLIC基因的表达量上升, 可增强基底侧质膜上Cl–通道蛋白的活性(Valenzuelaet al, 1997), 进而增强三疣梭子蟹对Cl–的吸收能力。然而当体内的Cl–水平被调节到正常水平时,PtCLIC基因又基本恢复到初始水平, 出现下调表达。
低盐胁迫下低盐耐受家系和低盐敏感家系差异显著, 低盐耐受家系中PtCLIC的相对表达量存在上调和下调的过程, 表达量最大值为低盐敏感家系的2.53倍, 而低盐敏感家系中PtCLIC的相对表达量基本不变且维持在较低水平。此差异证明在低盐耐受家系中PtCLIC存在一个积极的渗透压调节过程, 而低盐敏感家系几乎被动接受应激。该研究结果可辅助选育三疣梭子蟹耐低盐品系。
本研究首次成功克隆了三疣梭子蟹PtCLIC基因全长cDNA序列, 通过分析低盐度胁迫下三疣梭子蟹鳃和肝胰腺中PtCLIC基因的表达规律以及低盐耐受和低盐敏感家系中表达规律的差异, 进一步认定PtCLIC基因在渗透压调节中发挥了重要作用, 为三疣梭子蟹耐盐新品系的选育提供了理论指导。在后续的研究工作中, 关于三疣梭子蟹PtCLIC基因在渗透压调控方面的功能研究以及相关SNP位点筛选等将重点进行展开。
吕 富, 黄金田, 於叶兵, 2010. 盐度对三疣梭子蟹生长、肌肉组成及蛋白酶活性的影响. 海洋湖沼通报, 4: 137—142
江 山, 许强华, 2011. 盐度胁迫对三疣梭子蟹鳃Na+/K+-ATPase酶活的影响. 水产学报, 35(10): 1475—1480
孙颖民, 闫 愚, 孙进杰, 1984. 三疣梭子蟹幼体发育. 水产学报, 8(3): 219—226
陈丽娥, 谢 浩, 2010. ClC型氯离子通道的研究. 生命的化学,30(4): 545—549
季东升, 2005. 三疣梭子蟹池塘养殖技术. 特种经济动植物, 3:12—13
郑萍萍, 王春琳, 宋微微, 2010. 盐度胁迫对三疣梭子蟹血清非特异性免疫因子的影响. 水产科学, 29(11): 634—638
康敏华, 陈 暖, 杜喜玲等, 2011. 水稻氯离子通道蛋白基因的克隆及表达分析. 西北植物学报, 31(4): 0647—0653
隋延鸣, 高保全, 刘 萍, 2012. 三疣梭子蟹“黄选1号”盐度耐受性及适宜生长盐度分析. 大连海洋大学学报, 27(5):398—401
路允良, 王 芳, 赵卓英, 2012. 盐度对三疣梭子蟹生长、蜕壳及能量利用的影响. 中国水产科学, 19(2): 237—245
戴寒晶, 2006. 胞内氯离子通道蛋白功能的初步研究. 合肥:安徽医科大学硕士学位论文, 49—50
Adachi S, Uchida S, Ito Het al, 1994. Two isoforms of a chloride channel predominantly expressed in thick ascending limb of Henle’s loop and collecting ducts of rat kidney. Journal of Biological Chemistry, 269: 17677—17683
Berry K L, Bulow H E, Hall D Het al, 2003. AC. elegansCLIC-like protein required for intracellular tube formation and maintenance. Science, 302(5653): 2134—2137
Berryman M, Bretscher A, 2000. Identification of a novel member of the chloride intracellular channel gene family(CLIC5) that associates with the actin cytoskeleton of placental microvilli. Molecular Biology of the Cell, 11(5):1509—1521
Cornejo I, Niemeyer M I, Sepulveda F V, 2001. Cloning cellular distribution and functional expression of small intestinal epithelium guinea pig ClC-5 chloride channel. Biochimica et Biophysica Acta-Biomembranes, 1512(2): 367—374
Dong Y X, Chen Y, Wei Qet al, 2013. Cloning and polymorphism analysis of glutamate-gated chloride channel gene ofLaodelphax striatellus(Hemiptera: Delphacidae).Florida Entomologist, 96(3): 1168—1174
Dulhunty A F, Gage P W, Curtis Set al, 2001. The glutathione transferase structural family includes a nuclear chloride channel and a ryanodine receptor calcium release channel modulator. Journal of Biological Chemistry, 276(5): 3319—3323
Edwards J C, 1999. A novel p64-related Cl–channel: subcellular distribution and nephron segment-specific expression.American Journal of Physiology, 276(3): 398—408
Freire C A, Onken H, McNamara J C, 2008. A structure-function analysis of ion transport in crustacean gills and excretory organs. Comparative Biochemistry and Physiology Part A,151: 272—304
Hechenberger M, Schwappach B, Fischer W Net al, 1996. A family of putative chloride channels from A rabidopsis and functional complementation of a yeast strain with a CLC gene disruption. Journal of Biological Chemistry, 271(52):33632—33638
Heiss N S, Poustka A, 1997. Genomic structure of a novel chloride channel gene, CLIC2, in Xq28. Genomics, 45(1):224—228
Jentsch T J, Friedrich T, Schriever A, 1999. The CLC chloride channel family. European Journal of Applied Physiology,437: 783—795
Liu H, Peatman E, Wang W Qet al, 2011. Molecular responses of calreticulin genes to iron overload and bacterial challenge in channel catfish (Ictalurus punctatus). Developmental and Comparative Immunology, 35: 267—272
Lv J J, Liu P, Wang Yet al, 2013. Transcriptome analysis ofPortunus trituberculatusin response to salinity stress provides insights into the molecular basis of osmoregulation.PloS One, 8(12): 1—15
Perluigi S, Addabbo D, Luca P Let al, 2002. Segmental paralogy in the human genome: a large-scale triplication on lp, 6p and 21q. Mammalian Genome, 13(8): 456—462
Qian Z, Okuhara D, Abe M Ket al, 1999. Molecular cloning and characterization of a mitogen-activated protein kinaseassociated intracellular chloride channel. Journal of Biological Chemistry, 274(3): 1621—1627
Shorning B Y, Wilson D B, Meehan R R, 2003. Molecular cloning and developmental expression of two Chloride Intracellular Channel (CLIC) genes inXenopus laevis.Development Genes and Evolution, 213: 514—518
Uchida S, Sasakl S, Furukawa Tet al, 1993. Molecular cloning of a chloride channel that is regulated by dehydration and expressed predominantly in kidney medulla. Journal of Biological Chemistry, 268: 3821—3824
Valenzuela S M, Artin D K, Por S Bet al, 1997. Molecular cloning and expression of a chloride ion channel of cell nucleus. Journal of Biological Chemistry, 272(19): 12575—12582
Warton K, Tonini R, Fairlie W Det al, 2002. Recombinant CLIC1 (NCC27) assembles in lipid bilayers via a pH-ependent two-state process to form chloride ion channels with identical characteristics to those observed in Chinese hamster ovary cells expressing CLIC1. Journal of Biological Chemistry, 277(29): 26003—26011
猜你喜欢
海峡姐妹(2018年11期)2018-12-19 05:18:30
北京广播电视报(2018年39期)2018-11-23 06:01:14
现代营销·经营版(2017年3期)2017-05-17 10:19:46
作文周刊·小学五年级版(2016年1期)2016-06-29 03:31:18
广东海洋大学学报(2015年4期)2016-01-13 08:39:30
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
广东海洋大学学报(2015年3期)2015-12-22 10:05:30
首都医科大学学报(2015年4期)2015-12-16 13:00:08
华南农业大学学报(2015年5期)2015-12-04 03:04:38
中国洗涤用品工业(2015年5期)2015-02-28 19:02:26

- 海洋与湖沼的其它文章
- 大菱鲆(Scophthalmus maximus)、星斑川鲽(Platichthys stellatus)及其杂交种的形态学分析*
- 半滑舌鳎(Cynoglossus semilaevis Günther)生长激素体外重组表达及活性分析*
- 香港牡蛎(Crassostrea hongkongensis)与长牡蛎(Crassostrea gigas)种间杂种遗传力评估*
- 褪黑激素对香港细首纽虫(Cephalothrix hongkongiensis)性腺发育的影响及羟基吲哚-O-甲基转移酶(HIOMT)的测定*
- 饲养空间大小对单体筐养养殖系统中三疣梭子蟹(Portunus trituberculatus)的摄食行为与生长特性的影响研究*
- 凡纳滨对虾(Litopenaeus vannamei)Runt转录因子的基因克隆及其功能分析*