
淡色生赤壳菌(Bionectria ochroleuca)Bo-1菌株产生抗菌物质的发酵条件优化
2014-03-17 11:46刘琴英,蒋冬花,齐育平等
生物技术通报 2014年1期
淡色生赤壳菌(Bionectria ochroleuca)Bo-1菌株产生抗菌物质的发酵条件优化
刘琴英 蒋冬花 齐育平 孙蕾 陈璨 谢祥聪
(浙江师范大学 化学与生命科学学院,金华 321004)
以淡色生赤壳菌(Bionectria ochroleuca)Bo-1菌株发酵液乙酸乙酯粗提物对水稻白叶枯病菌(Xanthomonas oryzae pv. oryzae,Xoo)抑菌活性为检测指标,采用单因素试验优化Bo-1菌株产生抗菌物质培养所需的碳源、氮源、无机盐;通过正交试验优化培养基配方和摇瓶发酵条件。研究结果表明,Bo-1菌株产生抗菌物质适宜的碳源、氮源和无机盐分别为淀粉、蛋白胨、MgSO4·7H2O;优化的培养基配方为:淀粉30 g/L,蛋白胨2 g/L,MgSO4·7H2O 0.5 g/L;适宜的发酵条件为:温度30℃,转速150 r/min,装液量80 mL/250 mL,pH 6.5。
淡色生赤壳菌 正交试验 发酵 优化 抑菌活性
植物内生真菌(endophytic fungus)是指那些在其生活史中某一段时期生活在活的植物组织内、不引起植物组织产生明显病害症状的真菌[1]。研究发现,寄生在植物体内的内生真菌可以产生多种活性物质,如吡咯烷类生物碱、麦角碱、吲哚二萜等活性物质,可以提高植物的抗病菌、抗线虫的能力,如Istifadah等[2]研究发现,将内生真菌球毛壳菌(Chaetomium globosum)接种到小麦后,能够减轻由小麦德氏霉(Drechslera tritici-repentis)引起的小麦黄斑叶枯病的症状。因此,植物内生真菌具有作为新型抗菌药物筛选源的巨大潜力[3]。
生 赤 壳 属(Bionectria) 是1999年Rossman等[4]根据形态学及分子生物学特征将一部分丛赤壳科(Nectriaceae)菌种独立出来建立的一个属,被归类为生赤壳科(Bionectriaceae),肉座菌目(Hypocreales),子囊菌纲(Ascomycetes),子囊菌门(Ascomycota)。据相关研究报道,某些生赤壳菌(Bionectriaspp.)所产生的抗生素类物质可以对某些病原菌起特异性拮抗作用,如曹晋忠[5]从黄芩内分离出的淡色生赤壳菌N.SBA35对枯草芽孢杆菌
(Bacillus subtilis)、大肠埃希氏菌(Escherichia coli)、镰孢霉(Fusarium)等具有较好抑菌效果。
本实验室以水稻白叶枯病菌(Xoo)P6生理小种为指示菌,筛选得到一株对其抑菌效果明显的水稻内生真菌淡色生赤壳菌(Bionectria ochroleuca)Bo-1菌株。此外,其发酵液乙酸乙酯粗提物对大肠杆菌(Escherichia coli)、白色念珠菌(Monilia albican)、枯草芽孢杆菌(Bacillus subtilis)、李斯特氏菌(Listeria monocytogenes)、金黄色葡萄球菌(Staphyloccocus aureus)等也有较强抑菌活性。本研究以Bo-1菌株对白叶枯病菌的抑菌活性为检测指标,优化发酵工艺以期提高发酵液中抑菌活性物质的含量,并测定其次级代谢产物的最低抑菌浓度。
1 材料与方法
1.1 材料
1.1.1 供试菌株 淡色生赤壳菌(Bionectria ochroleuca)Bo-1菌株由本实验室从浙江省金华市水稻田健康水稻叶片内分离获得,保藏于中国典型培养物保藏中心,保藏号为CCTCC NO:M 2013027。
1.1.2 供试病原菌 水稻白叶枯病菌(Xoo)P6生理小种,由浙江师范大学生化学院遗传实验室提供。
1.1.3 主要培养基 马铃薯葡萄糖蛋白胨培养基(PDP):马铃薯200 g,葡萄糖20 g,蛋白胨1.0 g,水1 000 mL,pH6.0-6.5,(固体培养基加琼脂粉18 g);用于Bo-1菌株的培养与优化。
改良PD培养基:马铃薯300 g,蛋白胨5.0 g,蔗糖15.0 g,Ca(NO3)2·4H2O 0.5 g,NaHPO4· 12H2O 2.0 g,水1 000 mL,pH6.0-6.5,(固体培养基加琼脂粉18 g);用于XooP6生理小种的培养。
1.2 方法
1.2.1 Bo-1菌株发酵液粗提物的制备 采用液体发酵的方法,挑取已纯化菌丝体接种到装有约100 mL PDP液体培养基的250 mL锥形瓶中,于摇床上(220 r/min、28℃)振荡培养。培养约7 d后,获得发酵液。所得发酵液在室温下经纱布过滤后得滤液,滤液经乙酸乙酯萃取多次,直至上层乙酸乙酯层无色为止。将得到的乙酸乙酯层溶液合并,在旋转蒸发仪上蒸发浓缩至干即得Bo-1菌株发酵液粗提物。
1.2.2 Bo-1菌株抑菌活性的生物测定 参照文献[6,7],用牛津杯法测定发酵液粗提物对白叶枯病菌拮抗活性,具体操作如下:取保藏于-80℃冰箱XooP6生理小种接种于改良PD液态培养基摇床活化培养(120 r/min,28℃),当菌悬液浓度达到OD值0.6时,取100 μL菌液均匀涂布到PDA固体培养基上,待用;把发酵液粗提物用乙酸乙酯稀释成浓度为200 μg/mL的药液,待用;采用牛津杯法测定抑菌效果,用十字交叉法测量抑菌圈的大小。以乙酸乙酯溶液作为空白对照,每处理重复3次(以下所有试验均重复3次)。
1.2.3 发酵培养基优化
1.2.3.1 碳源优化 采用单因素试验[8]以PDP为基础培养基,将其中的碳源依次替换为蔗糖、乳糖、甘油、麦芽糖、淀粉,以基础培养基为对照。培养7 d后,按1.2.1方法处理发酵液,测定发酵液粗提物的抑菌活性。
1.2.3.2 氮源优化 采用单因素试验[8]以PDP为基础培养基,将其中的氮源依次替换为硝酸铵、胰蛋白胨、尿素、硝酸钾、牛肉浸膏,以基础培养基为对照。培养7 d后,处理发酵液,测定发酵液粗提物的抑菌活性。
1.2.3.3 无机盐优化 以优化的碳、氮源培养基作为对照,根据微生物对大量元素与微量元素的利用情况不同,按0.5 %的量分别加入NaCl和K2HPO4,按0.05%的量分别加入MgSO4·7H2O和CaCl2,按0.000 5% 的量分别加入FeSO4·7H2O,ZnSO4和CuSO4。培养7 d后,处理发酵液,按测定发酵液粗提物的抑菌活性。
1.2.3.4 培养基配方正交优化 在上述单因子筛选的基础上,以淀粉、蛋白胨、MgSO4·7H2O为考察因素,设置3个水平,用L9(34)正交设计试验(表1)[9]。培养7 d后,处理发酵液,测定发酵液粗提物的抑菌活性。
1.2.4 发酵条件正交优化 采用优化后的培养基,以装液量、初始pH、培养温度、摇床转速为考察因素,每个因素设计4个水平,用L16(45)正交设计试验(表2)[9]。培养7 d后,处理发酵液,测定发酵液粗提物的抑菌活性。

表 1 发酵培养基正交试验的因素和水平设计

表 2 发酵条件正交试验的因素和水平设计
1.2.5 优化验证试验 将活化后的Bo-1菌株分别接种到优化后培养基和原始培养基中,并对应采用优化后的培养条件和原始条件进行发酵。培养7 d后,处理发酵液,测定发酵液粗提物的抑菌活性。
1.2.6 最低抑菌浓度测定(MIC) 使用优化后的培养基及条件进行发酵,制备发酵液粗提物,称取一定质量的该粗提物溶于乙酸乙酯中,对倍稀释配制浓度分别为2 000、1 000、500、250、125、62.5、31.25、15.6、0 μg/mL的药液,琼脂稀释法[10,11]测定Bo-1菌发酵液粗提物对白叶枯病菌的MIC。以原始培养基及条件发酵所得的发酵液粗提物作为对照。
2 结果
2.1 发酵培养基优化
2.1.1 Bo-1菌株发酵的碳源优化 由表3可知,不同碳源对Bo-1菌株发酵液粗提物抑菌活性强弱明显不同。其中,以淀粉为碳源时,发酵液粗提物抑菌活性最强,抑菌圈直径达34.2 mm;其次蔗糖,抑菌圈直径为29.1 mm;在所有供试的碳源中,甘油最不适合该菌株产生抗菌物质,所得抑菌圈直径仅18.3 mm。由此可见,选用淀粉作为碳源最有利于Bo-1菌株产生抗菌物质。
2.1.2 Bo-1菌株发酵的氮源优化 由表4知,以不同氮源替代基础培养基蛋白胨后,其发酵液粗提物抑菌活性均不如蛋白胨(28.9 mm)。但各氮源间还是存在一定差异,牛肉浸膏仅次于蛋白胨,抑菌圈直径为27.5 mm;接着依次为胰蛋白胨、硝酸铵、硝酸钾、尿素。因此,选择蛋白胨作为氮源最有利于Bo-1菌株产生抗菌物质。

表 3 碳源对Bo-1菌株发酵液粗提物抑菌活性的影响

表 4 氮源对Bo-1菌株发酵液粗提物抑菌活性的影响
2.1.3 无机盐对Bo-1菌株发酵的影响 在优化后的碳、氮源培养基中,分别加入不同无机盐培养Bo-1菌株,所得发酵液粗提物抑菌活性有显著差异(表5)。与基础培养基相比,加入无机盐后,Bo-1菌株发酵液粗提物抑菌活性出现促进、抑制、无影响3种情况。当加入MgSO4·7H2O时,其抑菌能力略有升高,抑菌圈直径为37.6 mm(对照34.2 mm);当加入CuSO4和ZnSO4时,其抑菌活性显著下降,抑菌圈直径分别为22.1 mm、25.5 mm。其余4种无机盐(NaCl、K2HPO4、CaCl2、FeSO4·7H2O)的加入,对其抗菌活性均无明显影响。由此可见,在基础培养基中加入MgSO4·7H2O有利于Bo-1菌株产生抗菌物质。
2.1.4 优化的发酵培养基配方 正交优化试验结果见表6,由极差R值的大小可以看出,培养基配方的3种组分对Bo-1菌株抑菌活性能力影响的主次顺序为:B(蛋白胨)>A(淀粉)>C(MgSO4·7H2O)。对正交试验进行方差分析(表7)发现,仅B因素的P值小于0.05,因此,仅B因素对试验具有显著性的影响;F值由大到小依次为B>A>C,F数值越大表明该因素的变化对结果影响越大。由此可知,对Bo-1菌株发酵液粗提物抑菌活性的影响顺序为B、A、C,这与直观分析结果基本一致。综上分析,确定Bo-1菌株优化的发酵培养基配方为A1B1C2即淀
粉30 g/L,蛋白胨2 g/L,MgSO4·7H2O 0.5 g/L。

表 5 无机盐对Bo-1菌株发酵液粗提物抑菌活性的影响
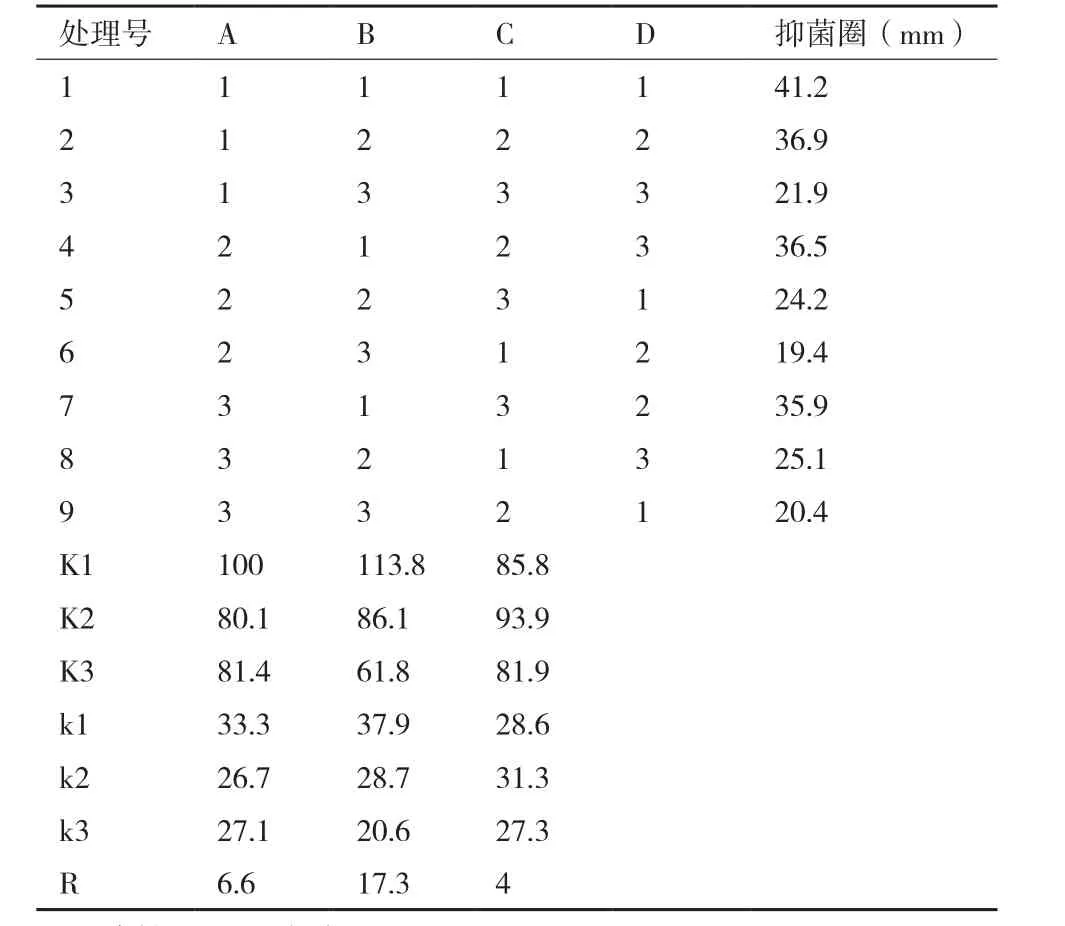
表 6 培养基配方正交试验结果
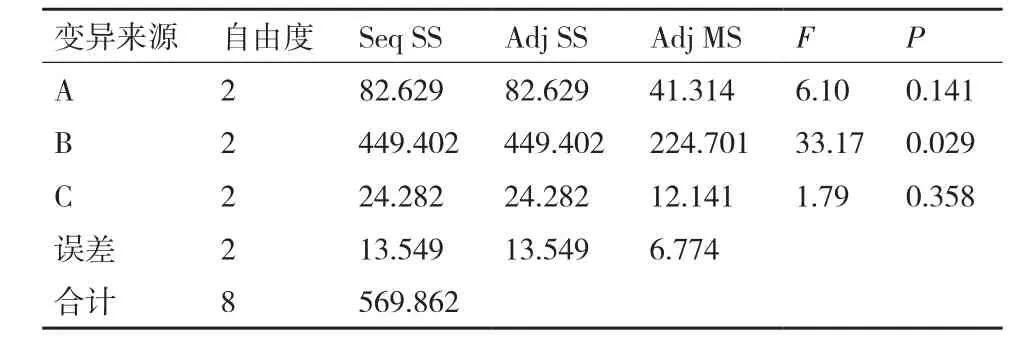
表 7 培养基配方正交试验方差分析
2.2 发酵条件的优化
从表8可以看出,发酵条件对Bo-1菌株发酵液粗提物抑菌活性有显著影响。由极差R值分析可见,4种因素对Bo-1菌抑菌活性能力影响的强弱顺序为:C(装液量)>A(温度)>B(转速)>D(pH)。装液量对其活性影响最显著,并与其活性呈负相关;pH对其活性影响最为微弱。对正交试验进行方差分析(表9)知,F值由大到小依次为C(装液量)>A(温度)>B(转速)>D(pH),这与直观分析结果基本一致。因此,最优发酵条件组合:A3B2C3D1即温度30℃,转速150 r/min,装液量80 mL/250 mL,pH6.5。
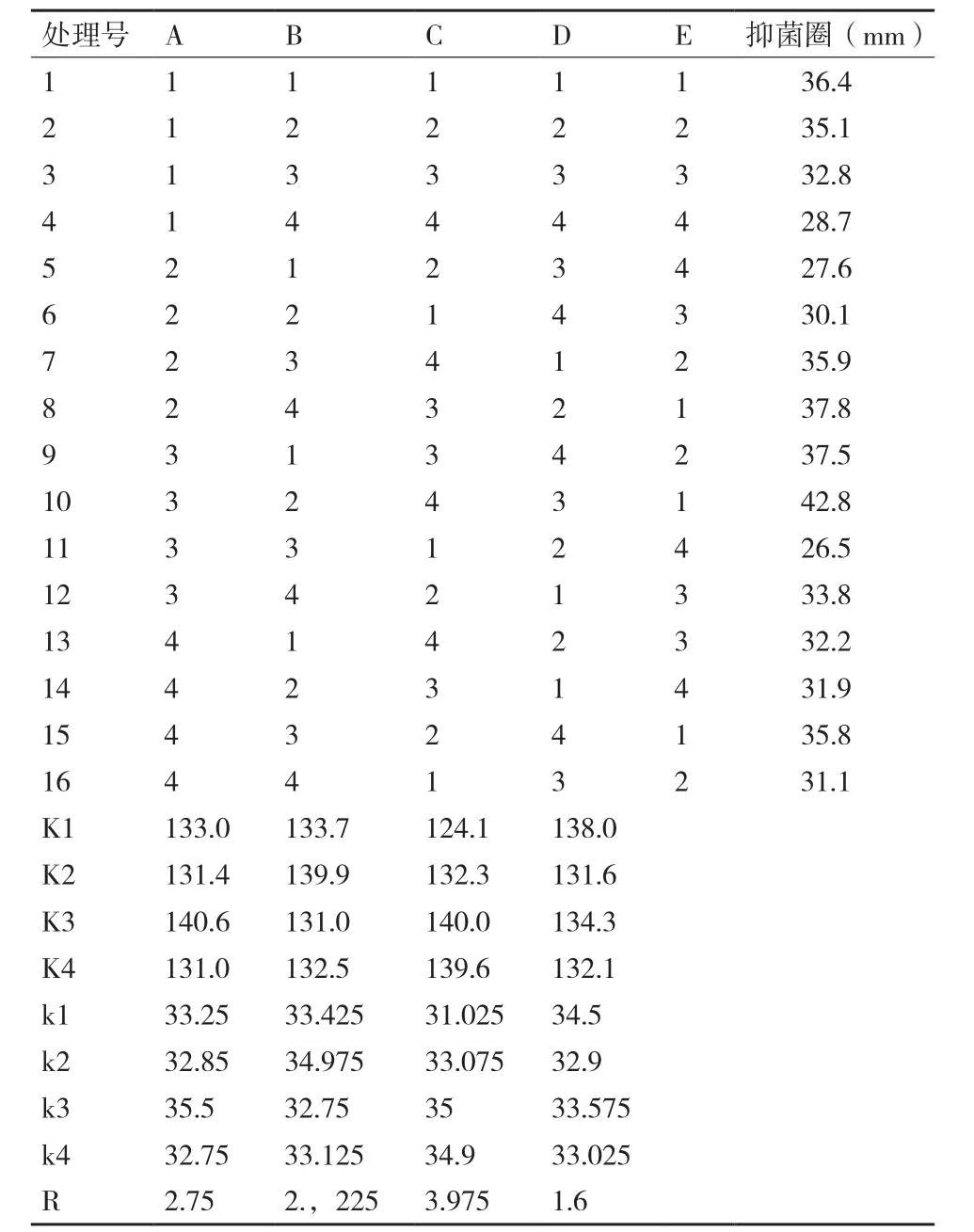
表 8 发酵条件正交试验结果

表 9 发酵条件正交试验方差分析
2.3 优化验证试验结果
采用优化过的培养基和条件发酵Bo-1菌株,其抑菌活性较原始抑菌活性(29.3 mm)显著增强,抑菌直径达42.5 mm(图1)。
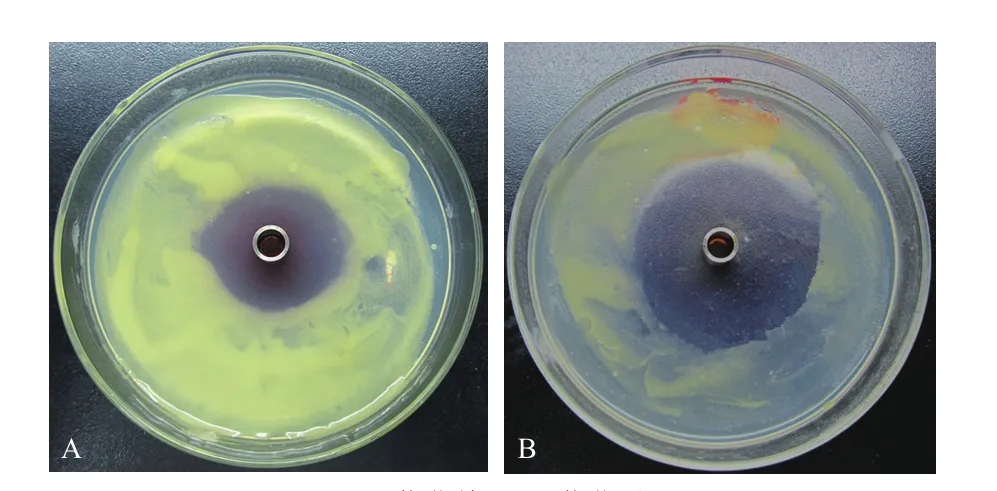
图 1 优化验证试验结果
2.4 最低抑菌浓度测定(MIC)
较原始Bo-1菌株发酵液粗提物对白叶枯病菌的MIC(250 μg/mL),优化后的MIC值有了显著的下降,其值仅为125 μg/mL。
3 讨论
微生物的发酵受外部环境条件的影响较大,不同的环境条件往往使得微生物具有不同的生长状态和不同的代谢途径,从而产生出不同的代谢产物[12]。在前期的研究中筛选得到了一株具有较强拮抗水稻白叶枯病菌活性的淡色生赤壳菌Bo-1菌株,本研究对其进行了一系列发酵条件优化试验,从而得到Bo-1菌株产生抗菌物质的最优培养条件,Bo-1菌株抑菌活性(42.5 mm)比优化前(29.3 mm)提高了45.1%。类似的报道,如胡小媛等[13]通过单因子法及响应面法优化布拉酵母培养基及培养条件,经验证发现,该菌在优化后的条件下培养,菌体湿重(32.2 g/L)比优化前(20.9 g/L)提高了54.1%,而活菌数(8.3×108CFU/mL)比优化前(5.5×108CFU/mL)提高50.9%;刘冬梅等[14]通过单因素试验与正交试验法相结合,优化Lactobacillus caseisubsp. rhamnosus LCR 719培养基配方及发酵条件后,其对S. aureus抑菌圈直径达42 mm,对枯草芽孢杆菌和肠炎沙门氏菌也有较好的抑菌效果。
4 结论
在优化条件下,Bo-1菌发酵液粗提物抗菌活性有了较大的提高,抑菌圈直径达42.5 mm,对水稻白叶枯病菌的MIC为125 μg/mL。
[1] Strobel G, Daisy B. Bioprospecting formicrobial endophytes and their natural products[J]. Microbiol Mol Biol Rev, 2003, 67(4):491-502.
[2] Istifadah N, McGee P. Endophytic Chaetomium globosum reduces development of tanspot in wheat caused by Pyrenophora tritici-repentis[J]. Australasian Plant Pathology, 2006, 35(4):411-418.
[3] 易晓华, 孙之潭, 胡美英, 等. 内生真菌的研究进展[J]. 仲恺农业技术学院学报, 2005, 18(10):61-65.
[4] Rossman AY, Samuels GJ, Rogerson CT, Lowen R. Genera of bionectriaceae, hypocreaceae and nectriaceae(Hypocreales, Ascomycetes)[J]. Stud Mycol, 1999, 42(7):1-260.
[5] 曹晋忠, 郑朝辉, 等. 4个黄芩内生真菌菌株的鉴定及抗菌活性研究[J]. 安徽农业大学学报, 2011, 38(3):393-399.
[6] 刘冬梅, 李理, 杨晓泉. 用牛津杯法测定益生菌的抑菌活力[J].食品研究与开发, 2006, 27(3):110-111.
[7] 董金甫, 李瑶卿, 洪绍梅. 茶多酚(TPP)对8种致病菌最低抑制浓度的研究[J]. 食品科学, 1995, 16(1):6-12.
[8] 段传人, 姚月良, 汪中文, 等. 海洋链霉菌Streptomyces sp. SCSIO 1666 活性代谢产物替达霉素A和B的发酵优化及分离鉴定[J]. 中国海洋药物, 2010, 12(6):12-20.
[9] 王如平, 吴婷, 等. 正交设计优选白芨多糖包合丹皮酚最佳工艺以及包合物的鉴定[J]. 食品科学, 2008, 29(12):259-263.
[10] 朱晓松, 贾林, 王蜜蜜, 等. 商陆提取物抑菌活性评价研究[J].中国现代中药, 2010, 12(12):33-39.
[11] 徐龙权, 陆军, 葛丽丽, 等. 红薯茎叶多糖提取物抑菌研究[J].食品科技, 2011, 36(1):163-166.
[12] 袁献温, 杨瑞丽, 曹雪. 抗肿瘤海洋放线菌ACMA006发酵条件的优化[J]. 中国海洋药物, 2008, 27(6):5-9.
[13] 胡小媛, 滕达, 张勇, 等. 防腹泻布拉酵母培养条件的响应面法优化[J]. 生物技术通报, 2013(6):194-199.
[14] 刘冬梅, 吴晖, 李理, 梁世中. Lactobacillus casei rhamnosus的抑菌作用及最佳抑菌作用发酵条件优化[J]. 食品科学, 2008, 6:237-242.
(责任编辑 李楠)
Fermentation Optimization of Antimicrobial Substance Produced by Bionectria ochroleuca Strain Bo-1
Liu Qinying Jiang Donghua Qi Yuping Sun Lei Chen Can Xie Xiangcong
(College of Chemistry and Life Science,Zhejiang Normal University,Jinhua 321004)
Based on the index of antimicrobial activity of ethyl acetate extracts from culture liquid of Bionectria ochroleuca strain Bo-1 to Xanthomonas oryzae pv. oryzae, the optimal carbon source, nitrogen source and salt for the production of antimicrobial substance by strain Bo-1 were tested by single factor experiments. The medium composition and fermentation conditions were optimized by orthogonal experiments. The results showed that the optimal carbon source, nitrogen source and salt were starch, peptone, MgSO4·7H2O, respectively. The optimal composition of the medium was 30 g/L starch, 2 g/L peptone, 0.5 g/L MgSO4·7H2O. The optimal fermentation conditions were the combination of temperature 30℃, agitation speed 150 r/min, medium volume 80 mL/250 mL, initial pH6.5.
Bionectria ochroleuca Orthogonal experiments Fermentation Optimization Antimicrobial activity
2013-07-29
国家自然科学基金项目(31270061)
刘琴英,女,硕士研究生;研究方向:应用微生物学;E-mail:rk371970746@163.com
女,教授,硕士生导师,研究方向:微生物学和植物病理学;E-mail:jdh@ zjnu. cn
猜你喜欢
江苏农业学报(2022年4期)2022-09-07
中国果业信息(2021年5期)2021-12-05
亚热带农业研究(2019年1期)2019-06-11
天然产物研究与开发(2018年4期)2018-05-07
中成药(2018年1期)2018-02-02
中国酿造(2016年12期)2016-03-01
中国酿造(2014年9期)2014-03-11
食品工业科技(2014年9期)2014-03-11
植物营养与肥料学报(2011年6期)2011-10-24
