
传统发酵牦牛酸奶中马克斯克鲁维酵母菌的分离鉴定及系统发育分析
2014-03-08 05:40王远微张诚民索化夷
食品科学 2014年15期
王远微,张诚民,索化夷,岳 华,李 键,汤 承,*
(1.西南民族大学生命科学与技术学院,四川 成都 610041;2.西南大学食品科学学院,重庆 400715)
传统发酵牦牛酸奶中马克斯克鲁维酵母菌的分离鉴定及系统发育分析
王远微1,张诚民1,索化夷2,岳 华1,李 键1,汤 承1,*
(1.西南民族大学生命科学与技术学院,四川 成都 610041;2.西南大学食品科学学院,重庆 400715)
对川西北部分牧区的10 份传统发酵牦牛酸奶样品进行酵母菌的分离,通过常规形态特征和26S rRNA基因测序分析鉴定出16 株马克斯克鲁维酵母菌(Kluyveromyces marxianus)。同源性分析显示16 株分离菌与已知马克斯克鲁维酵母的同源性高达99.3%~100%。16 株分离菌中形成明显的两种序列类型,其中10 株分离菌与另外6 株分离菌相比在扩增片段的第537位点上发生碱基缺失、第554位点上碱基由G突变为A、第564位点上碱基由A突变为T。系统进化分析显示两种序列类型的分离株也形成两个独立进化分支。
传统发酵牦牛酸奶;马克斯克鲁维酵母;分离鉴定;系统发育分析
传统发酵牦牛酸奶是由青藏高原地区牧民采用传统方法制作而成,其营养价值很高,蛋白质和脂肪含量分别达到4.91%和6.89%[1-4],且含有大量的挥发性脂肪酸[5],并富有良好的风味。其在维持肠道菌群生态平衡、促进肠道蠕动、促进消化、抗衰老、抗癌、提高人体免疫力等方面有很好的效果[6]。它是牧区非常传统的奶制食品,也是牧民重要的经济收入来源。
牦牛酸奶是由牦牛乳经传统发酵方法制得,其发酵过程受海拔地理环境、气候环境、发酵温度、发酵时间、制作方法、文化及奶源的影响,因此牦牛酸奶中微生物菌群非常复杂。Zhang Heping等[3]报道青海地区牦牛酸奶与其他发酵酸奶相比含有更多的乳酸菌和酵母菌,其中青海西北部的高原牦牛酸奶和青海东部的环湖牦牛酸奶中乳酸菌是优势菌,而青海南部的环湖牦牛酸奶中酵母菌是优势菌。Wu Xiaohe等[4]研究发现:3 种乳酸菌以及5 种酵母菌是西藏不同海拔地区牦牛酸奶中主要发酵菌群,其中发酵乳杆菌是优势菌。Koichi[7]对蒙古不同动物的传统发酵酸奶的中乳酸菌和酵母菌的分离研究中发现,牦牛等动物酸奶中乳酸菌数量明显高于酵母菌数量。从以上研究结果中可以看出牦牛酸奶中的微生物菌群尽管比较复杂,但是都含有乳酸菌和酵母菌,这也符合牦牛酸奶为典型的IV型发酵乳的特征,这种类型的发酵乳中优势微生物除乳酸菌外还有酵母菌[8],只是有的地区的牦牛酸奶的优势菌是乳酸菌,有的是酵母菌,而且优势的乳酸菌和酵母菌的种类也有所不同。
随着对传统发酵乳制品中酵母菌研究的深入,越来越多的酵母菌种属在各种乳制品中被检出,克鲁维酵母就是其中之一。有报道在开菲尔酸奶、蒙古酸马奶、苏丹发酵乳Rob、匈牙利发酵乳、意大利牛乳、水牛乳、山羊乳以及希腊的山羊干酪中都分离到该属菌株[9-11]。马克斯克鲁维酵母是克鲁维酵母属中重要的一个种,该菌株被认为是开菲尔酸奶和苏丹发酵乳Rob中的优势酵母菌[12-13]。国内Bai Mei等[14]也分别从青海地区和西藏地区传统发酵牦牛酸奶中分离出马克斯克鲁维酵母,但对川西北高原牧区传统发酵牦牛酸奶中马克斯克鲁维酵母菌的分离报道比较少。
本实验对川西北地区的红原、若尔盖、康定等地的10 份传统发酵牦牛酸奶样品进行酵母菌的分离鉴定,并对分离菌进行了系统进化分析,以期对川西北高原地区传统发酵牦牛酸奶中蕴含的大量原始微生物菌种资源的保护及研究提供菌株,为该地区传统发酵牦牛酸奶中微生物组成研究以及马克斯克鲁维酵母菌的遗传多样性研究提供参考。
1 材料与方法
1.1 材料与试剂
样本采自川西北地区的四川省阿坝洲红原县(3 份)、若尔盖县(3 份)和甘孜州康定县(4 份),样本置于4 ℃样品箱运到实验室,立即进行分离培养。
酵母膏胨葡萄糖(yeast extract peptone dextrose,yPD)培养基、革兰氏染液 杭州微生物制剂有限公司;Taq DNA聚合酶、聚合酶链式反应(polymerase chain reaction,PCR)试剂、DNA Marker、dNTPs 日本TaKaRa公司;琼脂糖 美国Oxoid公司;细菌基因组DNA提取试剂盒 天根生化科技(北京)有限公司;AXy prepTMDNA Gel extraction kit 美国Axygen公司。
1.2 仪器与设备
MyCyclerTMPCR仪、Powerpac UniversalTM核酸电泳仪、VerSa Doc2000凝胶成像系统 美国Bio-Rad公司;超纯水仪Milli-Q 法国Millipore公司;HR40- ⅡA2 生物安全柜 青岛海尔特种电器有限公司;AR2130/C型精密电子天平 梅特勒- 托利多仪器(上海)有限公司。
1.3 方法
1.3.1 酵母菌的分离与纯化
吸取1 mL牦牛酸奶置于装有9 mL无菌生理盐水的试管中,充分振荡使其混匀。将混合后牦牛酸奶进行梯度稀释,稀释度分别为10-1、10-2、10-3、10-4、10-5、10-6、 10-7。选取10-3、10-4、10-5、10-6、10-75 个稀释度,吸取1 mL 样液采用涂布法均匀涂布于yPD培养基上,将制备好平板置于28 ℃恒温培养箱中培养24~48 h,观察并记录菌落特征,挑取菌落较大,呈白色或乳白色的单个菌落进行革兰氏染色,观察细胞形态,呈圆形或椭圆形、腊肠状且细胞较大者,再将其接种于yPD培养基,于28 ℃恒温培养箱中培养24~48 h,重复几次纯化酵母菌,直至镜检结果为同一细胞形态后,将其接种于yPD液体培养基中,于28 ℃恒温培养24~48 h后,加入30%的灭菌甘油,混匀后置于-80 ℃保存备用。
1.3.2 形态结构观察
将上述纯培养菌株接种于yPD培养基斜面,置28 ℃恒温培养18 h后涂片,经革兰氏染色后镜检菌体形态。
1.3.3 酵母菌的26S rRNA序列测定与系统发育学分析
1.3.3.1 PCR模板DNA的制备
将上述纯化的酵母菌接种于yPD液体培养基中置于28 ℃振荡培养24 h,取培养液按照细菌基因组DNA提取试剂盒的操作说明提取细菌基因组DNA,作为本实验的PCR模板。
1.3.3.2 PCR引物
PCR扩增的目的片段在酵母菌的大亚基26S rRNA的D 1/D 2可变区域内,引物序列N L-1:5’-GCATATCAATAAGCGGAGGAAAAG-3’,NL-4:5’-GGTCCGTGTTTCAAGACGG-3’[15],扩增的目的片段大小在500~600 bp,引物由生工生物工程(上海)股份有限公司合成。
1.3.3.3 PCR方法
PCR反应采用25 μL反应体系:DNA模板1 μL、上下游引物各1 μL(10 pmol)、 4×dNTPs(10 mmol/L)2 μL、Taq酶(5 U/uL)0.125 μL、Mg2+3 μL(10 mol/mL)、10×PCR buffer 2.5 μL,超纯水补足至25 μL。PCR 程序如下:95 ℃预变性5 min,94 ℃ 1 min、52℃ 1 min、72 ℃1.5 min,36 个循环;72 ℃温浴10 min,降至室温后使用2%琼脂糖凝胶电泳检测。
1.3.3.4 目的片段的纯化回收及测序
按照1.3.4.3节的反应体系和反应条件扩增100 μL PCR产物,2%琼脂糖凝胶电泳后按照回收试剂盒操作说明进行目的片段的纯化回收,回收后再2%琼脂糖凝胶电泳检测。将电泳检测后的回收产物送生工生物工程(上海)股份有限公司进行测序。
1.3.3.5 序列分析与系统发育树构建
将上述测序得到的26S rRNA基因序列通过NCBI的BLAST检索系统进行序列同源性分析,选择同源性大于99%的10 个酵母序列的片段,同时调取GenBank数据库中收录的克鲁维菌属的18 个种的32 株酵母的相同区段基因序列,使用ClustalX1.83软件进行多序列匹配比对,通过Mega4.1软件采用Neighbor-Joining法构建分离酵母的同源序列系统进化树和克鲁维酵母属内系统进化树,采用自举分析进行置信度检测,自举数据集为3 000 次。
2 结果与分析
2.1 酵母菌分离菌株的菌落形态及细胞形态特征
从四川红原、若尔盖、康定等地的10 份传统发酵牦牛酸奶样品中分离到的16 个优势菌株,菌落形态呈圆形,表面光滑湿润,颜色呈乳白色,表面凸起、边缘整齐、不透明(图1)。显微镜下细胞染色为蓝紫色,形态为圆、椭圆、卵圆等,无性繁殖为芽殖,一端出芽,细胞形态符合酵母菌特征(图2)。

图1 分离菌的菌落形态Fig.1 Colony morphology of isolates
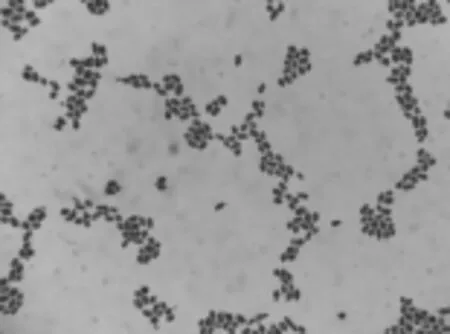
图2 分离菌的细胞形态(1 000×)Fig.2 Cell morphology of isolates (1 000×)
2.2 酵母菌26S rRNA序列PCR扩增结果
按照PCR反应条件进行扩增,经琼脂糖电泳检测结果显示酵母26S rRNA基因PCR扩增产物在约587 bp左右处出现一条明亮的条带,与预期片段大小相符(图3)。

图3 分离菌的26S rRNA基因的PCR扩增结果Fig.3 PCR amplification of 26S rRNA genes from 16 isolates
2.3 酵母菌26S rRNA基因测序同源比对及系统发育分析
经26S rRNA基因序列测定,扩增序列长度为586~588 bp,与Genbank中同源性较高的菌株进行同源性比对。结果显示,分离的16 株酵母菌与Genbank中马克斯克鲁维酵母的同源性最高,相似度为99.3%~100%。序列分析显示其中10 株分离菌与另外6 株分离菌形成明显的两种序列类型,10 株分离菌在扩增片段的第537位点上发生碱基缺失、第554位点上碱基由G突变为A、第564位点上碱基由A突变为T。
(4)某植物夏日晴天中午12:00时叶片的光合速率明显下降,其原因是进入也有细胞的(填“O2”或“CO2”)_______不足。

图4 分离株和同源性较高菌株的26S rRNA基因序列系统发育树Fig.4 Phylogenic tree of Kluyveromyces marxianus and 16 isolates based on 26S rRNA gene sequences

图5 克鲁维菌属26S rRNA基因序列系统发育树Fig.5 Phylogenic tree of Kluyveromyces based on 26S rRNA gene sequences
图4为16株分离菌与Genbank中同源性最高的7 株马克斯克鲁维酵母的序列构建的系统发育进化树。由图4可知,与序列分析显示的两种序列类型的结果相同,16 株酵母分离株也成两个大的分支,其中6 株分离菌与Genbank中的7株马克斯克鲁维酵母聚在一个分支,另外10 株分离菌聚在另一个大的分支上。
图5为16 株分离菌与GenBank数据库中收录的克鲁维菌属的18 个种的32 株酵母菌的相同区段基因序列构建的系统发育进化树。由图5可知,克鲁维菌属的18 个种的菌株分在不同的分支上,16 株分离菌与3 株马克斯克鲁维酵母和3 株乳酸克鲁维酵母分在一个大的分支上,3 株乳酸克鲁维酵母形成一小的分支,6 株分离株与3株马克斯克鲁维酵母形成一个小的分支,另外10 株分离菌单独形成一个小的分支,这也与序列分析显示的两种序列类型的结果相同。
3 讨 论
Kuttzman等[15]测定了大约500 种酵母的大亚基26S rRNA基因的D1/D2可变区的序列,包括假丝酵母和其他有性型及无性型子囊菌酵母几乎所有已知种的模式菌株,发现用这段序列(500~600 bp)可以将绝大部分种区分开,同一种内不同菌株间D2区的核苷酸差异不超过1%。这些序列均已公布在GenBank 等国际核酸序列数据库中,给酵母菌的分子鉴定提供了很大方便。本实验扩增了16 株分离菌的26S rRNA基因的D1/D2可变区进行测序分析,对分离菌进行分子鉴定,结果发现16 株酵母菌与马克斯克鲁维酵母的同源性最高,相似度为99.3%~100%,按照Kuttzman的同一种内不同菌株间D2区的核苷酸差异不超过1%的结论,因此将16 株分离菌鉴定为马克斯克鲁维酵母。本实验为川西北高原地区传统发酵牦牛酸奶中原始微生物菌种资源的保护及研究提供了菌株。
本实验从川西北地区采集的10 份传统发酵牦牛酸奶样品中均分离到了马克斯克鲁维酵母菌,分离率很高,共获得了16 株分离菌,可见马克斯克鲁维酵母在该地区牦牛酸奶中是一种广泛存在的菌株。本实验为该地区传统发酵牦牛酸奶的微生物组成研究提供参考。马克斯克鲁维酵母是克鲁维酵母属的一个成员,因其具有多种生物学作用[16],得到了国内外相关专家广泛的关注和研究。发酵乳制品具有独特的风味是因为在发酵过程中产生的大量的芳香化合物,马克斯克鲁维酵母可以产生芳香化合物,因此在某些乳制品中分离到马克斯克鲁维酵母可能是其具有独特风味的原因。有相关报道在发酵可可的时候添加马克斯克鲁维酵母可以改变可可的风味[17]。马克斯克鲁维酵母还可以减少食品中的乳糖含量,在哺乳动物奶汁里,乳糖的含量达99.8%以上,但是除了新生儿之外,成人体内的乳糖酶的活性都很低,“乳糖不耐受”人群体内的乳糖酶活力尤其低,因此成人摄取的乳制品中的乳糖含量过多不但不能成为营养物质,还可以引起高血糖和肥胖等症状。马克斯克鲁维酵母具有减少食品中的乳糖含量的作用,对乳制品的营养组成和人类的健康都有很大的意义。而且马克斯克鲁维酵母通过发酵分解乳糖,可以产生多种羰基化合物和挥发性脂肪酸,这些芳香物质也是形成乳制品独特的风味的主要原因之一。马克斯克鲁维酵母在该地区牦牛酸奶中分离率很高,因此对该地区牦牛酸奶的风味和营养价值研究具有重要的参考价值。
26S rRNA基因是核糖体RNA基因中的一员,具有进化速率慢、序列保守性高的特点,但同时由于各种原因其基因序列中又具有一定的突变,而这些突变的序列具有种属的差异,使其作为一种良好的生物分类鉴定材料而受到广泛的重视,被作为种属的鉴定以及分子分型的工具[18-19]。本实验中对分离酵母的系统发育分析显示16 株分离菌其中10 株分离菌与另外6 株分离菌形成明显的两种序列类型,10 株分离菌在扩增片段的第537位点上发生碱基缺失、第554位点上碱基由G突变为A、第564位点上碱基由A突变为T,构建的进化树中两种序列类型的分离菌也形成两个独立分支,说明马克斯克鲁维酵母在遗传进化过程中具有一定多样性,为马克斯克鲁维酵母菌的遗传多样性研究提供参考。发生变异的3 个碱基位点可能是在一定的条件下该种酵母产生的特异突变,推测其可能是这10 个分离株的单核苷酸多态性位点,而作为分子分型的遗传标记。形成独立分支的10 株分离菌出现以上碱基的变异,可能是由于在四川红原、若尔盖、康定等高原地区的高海拔环境条件下经过漫长的选择进化造成的,也可能是在牦牛酸奶低温发酵条件下长期的选择进化引起的。发生3 个碱基位点变异有可能引起10 株马克斯克鲁维酵母分离菌表型特性的变异,例如发酵特性、耐热、耐酸等特性的变化。如果该10 个分离株发生表型变异而具有特殊的表型特征,那这3 个SNP位点则可能作为具有该特殊表型的马克斯克鲁维酵的遗传标记,用于该类马克斯克鲁维酵的筛选和鉴定。以上的推测还需要进一步实验用更多的分离菌株进行验证,但该结果为马克斯克鲁维酵母的遗传多样性研究、分子分型研究以及遗传标记研究提供一定的参考。
从川西北地区的红原、若尔盖、康定等地的10 份传统发酵牦牛酸奶样品中分离鉴定出16 株马克斯克鲁维酵母。序列分析显示16 株马克斯克鲁维酵母分离株形成两种不同的序列类型,系统进化分析分离株也形成两个独立进化分支。本实验为川西北高原地区传统发酵牦牛酸奶中蕴含的大量原始微生物菌种资源的保护及研究提供菌株,为该地区传统发酵牦牛酸奶中微生物组成研究以及马克斯克鲁维酵母的遗传多样性研究提供参考。
[1] 吴春生, 李键, 骞宇, 等. 牦牛乳及牦牛酸乳营养价值的研究现状[J].乳业科学与技术, 2012, 35(3): 43-46.
[2] 张国强, 马长中, 李亚辉, 等. 西藏发酵牦牛乳化学成分和微生物组成分析[J]. 食品研究与开发, 2011, 30(2): 4-7.
[3] ZHANG Heping, XU Jie, WANG Junguo, et al. A survey on chemical and microbiologicalcomposition of kurut, naturally fermented yak milk from Qinghai in China[J]. Food Control, 2008, 19(6): 578-586.
[4] WU Xiaohe, LUO Zhang , YU Li, et al. A survey on composition and micro biota of freshand fermented yak milk at different Tibetan altitudes [J]. Dairy Science and Technology, 2009, 89(2): 201-209.
[5] 苟钰姣, 丁路明, 王玉鹏. 牦牛乳及乳制品、犏牛和黑白花奶牛乳的脂肪酸组成分析[J]. 草业科学, 2013, 30(2): 274-280.
[6] 梁春年. 牦牛乳的特性及利用现状[C]//第四届中国牛业发展大会论文集. 北京: 中国畜牧业协会, 2009: 345-348.
[7] KOICHI W. Diversity of lactic bacteria and yeasts in Airag and Tarag, traditional fermented milk products of Mongolia[J]. World Journal of Microbiology and Biotechnology, 2008, 24(8): 1313-1325.
[8] 吴春生, 舒畅, 李键, 等. 牦牛酸乳中乳酸菌的研究进展及前景展望[J].食品工业, 2012, 33(9): 129-133.
[9] VASDINyERI R, DEAK T. Characterization of yeast isolates originating from Hungarian dairy products using traditional and molecular identif i cation techniques[J]. International Journal of Food Microbiology, 2003, 86(1/2): 123-130.
[10] SIRNOVA E, BESHKOVA D, ANGELOV A, et al. Lactic acid bacteria and yeasts in kefir grains and Kefir made from them[J]. Journal of Industrial Microbiology and Biotechnology, 2002, 28(1): 1-6.
[11] SHUANG Q, BURENTEGUSI, BAI y, et al. Microf l ora in traditional starter cultures for fermented milk, hurunge, from Inner Mongolia, China[J]. Animal Science Journal, 2006, 77(2): 235-241.
[12] ABDELGADIR W S, HAMAD S H, MOLLER P L, et al. Characterisation of the dominant microbiota of Sudanese fermented milk Rob[J]. International Dairy Journal, 2001, 11(1/2): 63-70.
[13] LATORRE G L, CASTILLO A L, POLAINA J. Taxonomical classif i cation of yeasts isolated from Kef i r based on the sequence of their ribosomal RNA genes[J]. World Journal of Microbiology and Biotechnology, 2007, 23(6): 785-791.
[14] BAI Mei, QING Manjun, GUO Zhuang, et al. Occurrence and dominance of yeast species in naturally fermented milk from the Tibetan Plateau of China[J]. Canadian Journal of Microbiology, 2010, 56(9): 707-714.
[15] KURTZMAN C K, ROBNETT C J. Identification and phylogeny of ascomycetous yeasts fromanalysis of nuclear large subunit (26S) ribosomal DNA partial sequences[J]. Antonie van Leeuwenhoek, 1998, 73(4): 331- 371.
[16] GUSTAVO G F, ELMAR H, CHRISTOPH W, et al. The yeast Kluyveromyces marxianus and its biotechnological potential[J]. Applied Microbiology and Biotechnology, 2008, 79(3): 339-354.
[17] CRAFACK M, MIKKELSEN M B, SAERENS S, et al. Influencing cocoa flavour using Pichia kluyveri and Kluyveromyces marxianus in a defined mixed starter culture for cocoa fermentation[J]. International Journal of Food Microbiology, 2013, 167(1): 103.
[18] GARNER C D, STARR J K, MCDONOUGH P L. Molecular identification of veterinary yeast isolates by use of sequence-based analysis of the D1/D2 region of the large ribosomal subunit[J]. Journal of Clinical Microbiology, 2010, 48(6): 2140-2146.
[19] BALEIRAS-COUTO M M, GOMES A S , CASAL M, et al. Survey of yeast diversity during wine bottling processes using restriction analysis of 26S ribosomal DNA (rDNA)[J]. Australian Journal of Grape and Wine Research, 2012, 18(1): 39-42.
Isolation, Identification and Phylogenetic Analysis of Kluyveromyces marxianus Strains from Traditional Fermented Yak Yoghurt
WANG yuan-wei1, ZHANG Cheng-min1, SUO Hua-yi2, yUE Hua1, LI Jian1, TANG Cheng1,*
(1. College of Life Science and Technology, Southwest University for Nationalities, Chengdu 610041, China; 2. College of Food Science, Southwest University, Chongqing 400715, China)
Based on the conventional morphological and 26S rRNA gene sequencing analysis, 16 stains of Kluyveromyces marxianus were isolated and identified from 10 samples of traditional fermented yak yogurt in the northwest of Sichuan province. The 16 isolates had 99.3%-100% nucleotide sequence homology with that of K. marxianus available in GenBank. Two quite different sequence types were observed in 16 isolates; one had 10 isolates, the other contained 6 isolates. Compared with the 6 isolates, the 10 isolates had a base deletion at position 537, a point mutation G-A at position 554, and a point mutation A-T at position 564 in the amplified fragments. In the phylogenetic tree, two different sequence types of K. marxianus isolates fell into two distinct groups. Therefore, these results can provide useful information for understanding the microbial composition of traditional fermented yak yogurt and genetic diversity of K. marxianus.
traditional fermented yak yoghurt; Kluyveromyces marxianus; isolation and identification; phylogenetic analysis
TS201.3;TS252.54
A
1002-6630(2014)15-0216-05
10.7506/spkx1002-6630-201415044
2013-08-22
王远微(1982—),男,助教,硕士,研究方向为分子生物学。E-mail:18782269822@139.com
*通信作者:汤承(1965—),男,教授,博士,研究方向为分子生物学。E-mail:tangcheng101@163.com
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
军事文摘·科学少年(2021年1期)2021-02-04
教学考试(高考生物)(2020年6期)2020-11-23
作文小学中年级(2020年6期)2020-07-24
作文小学中年级(2020年4期)2020-06-11
作文小学中年级(2020年2期)2020-04-26
食品与生物技术学报(2020年8期)2020-01-06
科学24小时(2019年5期)2019-06-11
当代水产(2019年3期)2019-05-14
发明与创新(2019年9期)2019-03-26
