
云杉叶栖真菌的群落结构与多样性1)
2014-03-06 03:19:40许秀兰黄晓丽刘应高
东北林业大学学报 2014年3期
许秀兰 张 翅 黄晓丽 张 雪 刘应高
(四川农业大学,雅安,625014) (成都温江花卉园林局) (四川农业大学)
树栖真菌泛指生活在林木上的所有真菌,是森林微生物系统的重要组成部分[1],其中内生真菌[2]被作为具有潜在利用价值的真菌资源得到广泛的重视[3-7]。截止目前,这一类丰富的真菌资源仍未得到充分的研究和利用。四川西部康定地区的狭小区域是云杉(Picea)的现代分布中心[8],该区域主要云杉树种有:粗枝云杉(P.asperata)、川西云杉(P.likianensis var.balfouriana)、麦吊云杉(P.brachytyla)、丽江云杉(P.likianensis)、紫果云杉(P.purpurea)、白皮云杉(P.aurantiaca)、鳞皮云杉(P.retroflexa)等。目前报道的云杉叶部病害最为常见[9-13]。近年来,在四川二郎山地区也发现存在严重的云杉针叶枯死脱落的现象,严重影响云杉林的生长。本研究以云杉人工林中的云杉针叶为研究对象,对云杉内生真菌的群落组成、结构和多样性进行研究分析,探索该地区云杉发病的原因,也可为后期筛选有效的生物拮抗菌奠定一定基础。
1 研究方法
1.1 样品采集
云杉针叶样品采自二郎山林场的幼龄云杉林,采样时间自9月份到次年7月份。每2月采集一次样品,共计6 次。同时以海拔100 m 的差值设置5个样点:2 480、2 580、2 680、2 780、2 880 m。并按树冠的上、中、下进行取样。
1.2 真菌的分离与纯化
将云杉针叶切成5.0 mm 小段。处理Ⅰ:表面消毒,用70%的酒精处理30 s,0.1%升汞溶液表面消毒3 min,最后无菌水漂洗3 次,移至PDA 培养基。处理Ⅱ:表面不消毒,无菌水漂洗3 次后移至PDA 培养基。每个样接种6 皿,每皿5 小段。置于人工气候培养箱23 ℃恒温培养,记录菌落形态,采用尖端菌丝挑取法,在无菌条件下挑取形态不同、独立的菌落转入PDA 上纯培养。同时统计分离率[14-16]。
1.3 真菌的形态分类鉴定
在PDA 培养基上所得的纯培养,通过水载片,在显微镜下观察其产孢结构及孢子性状,并结合菌落特征参考相关资料数据对各产孢菌株进行鉴定。采用不同培养基的方法对未产孢菌株进行产孢诱导[17]。
1.4 分子辅助分类鉴定
1.4.1 真菌总DNA 的提取
将PDA 培养基上培养的菌株用打孔法接种于PDB 培养液中,25 ℃振荡培养5 ~7 d,无菌纱布过滤收集菌丝体,再用无菌滤纸将菌丝吸干,-20 ℃冷冻备用。采用改良二次沉淀法提取总DNA:将冷冻变硬的菌丝体转移到已冷冻的研钵中,分别加入液氮2 ~3 次,把冷冻的菌丝体研磨成细粉末;迅速将粉末转移到20 mL 离心管中,加入10 mL 缓冲液A(SDS-EB,pH=8.0),密封后在旋涡振荡器上混匀并静置至室温;将混合液置于65 ~68 ℃水浴20 min,室温10 000 r/min 离心15 min;取上清液移至另一离心管,加入0.5 mL缓冲液B(8 MKOAc,pH =4.2),混匀后冰浴45 min,于4 ℃下10 000 r/min 离心20 min;取上清液至另一试管,加入等体积的异丙醇,小心颠倒混合均匀,可以观察到沉淀出现;将离心管静置5 min,倒掉上清液,将沉淀转移到1.5 mL的离心管中,用70%乙醇清洗1 次,无水乙醇清洗2次,然后让沉淀自然干燥;将沉淀溶解于50 μL 超纯水,混匀后于冰箱中-20 ℃保存备用。
1.4.2 rRNA 基因ITS 区的PCR 扩增及测序
采用真菌通用引物ITS1 和ITS4 进行扩增,ITS1(5'TCCGTAGGTGAACCTGCGG- 3'),ITS4(5'TCCTCCGCTTATTGATATGC-3')。反应体系为50.0 μL,DNA 模板0.5 μL,dNTP mixture1.0 μL,ITS1 1.0 μL,ITS4 1.0 μL,Taq plus 酶0.5 μL,10 ×Buffer 5.0 μL,MgCl22.0 μL,超纯水39.0 μL。扩增条件:95 ℃预变性5 min,30 个循环扩增(95 ℃1 min,50 ℃1 min,72 ℃1 min),然后72 ℃延伸5 min,最后于4 ℃保存。产物用0.8%琼脂糖电泳进行检测。送至上海生工生物有限公司进行DNA 序列测定。
1.5 数据处理
以生长在不同海拔、不同树体高度的云杉针叶为云杉叶栖真菌群落的栖息地结合采集样本的不同季节,进行多样性与均匀度比较。测度方法选用Simpson 的多样性指数(λ,1949)、Shannon-Wiener的多样性指数(H',1949)、Pieloum 的均匀度指数(E1,1977)、Sheldon 的均匀度指数(E2,1969)和Heip 的均匀度指数(E3,1974),计算公式为:
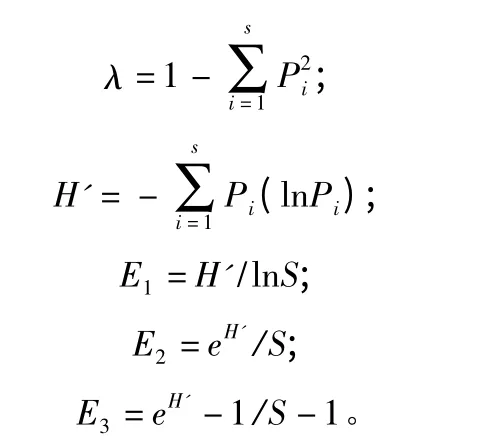
式中:S 为某部位树栖真菌群落中的真菌种类;Pi为该部位树栖真菌群落中第i 种的检出率或称出现频率;H'为该部位树栖真菌群落中S 个种的已知分离率总和;i=1,2,3,…,S[18]。
2 结果与分析
2.1 不同冠层、叶龄的云杉叶栖真菌的种群组成
经分离鉴定,自云杉树冠上层针叶分离得到16个属的19 种真菌。其中,表面消毒处理(Ⅰ)的针叶上有11 种,以Lophodermium sp.1、Pestalotiopsis sp.1、Phomopsis 为优势种群;不消毒处理(Ⅱ)的针叶上检出真菌16 种,其中Pestalotiopsis sp.1、Aspergillus、Penicillium sp.2 的检出率相对较高,为优势种。不同龄级真菌的种类差异较大,其中Pestalotiopsis sp.,Aspergillus sp.在各龄级针叶中都有分布。详见表1。

表1 云杉树冠上层针叶叶栖真菌检出率
中冠层针叶组织分离结果见表2。自云杉树冠中层的针叶上分离得到17 种真菌,分属于14 个属。表面消毒处理的针叶上获得真菌9 种,Preussia sp.2的检出率最高;不消毒处理的针叶上检出真菌12种,其中Epicoccum nigrum 和Penicillium sp.1 的检出率相对较高。叶龄不同,真菌的种类和数量都有差异。在各龄级都有分布的真菌种类有Penicillium sp,Epicoccum nigrum sp.,且数量较多。

表2 云杉树冠中层针叶叶栖真菌检出率
下冠层组织分离结果见表3。共分离得到26种真菌分别归属于20 个属。经表面消毒处理的针叶上获得真菌14 种,以Pestalotiopsis sp.2 为优势种群;不消毒处理的针叶上检出真菌20 种,其中Epicoccum nigrum、Xylaria sp.的检出率相对较高。各龄级针叶真菌种类差异大,Preussia sp.,Epicoccum nigrum 为各龄级针叶共有的真菌。
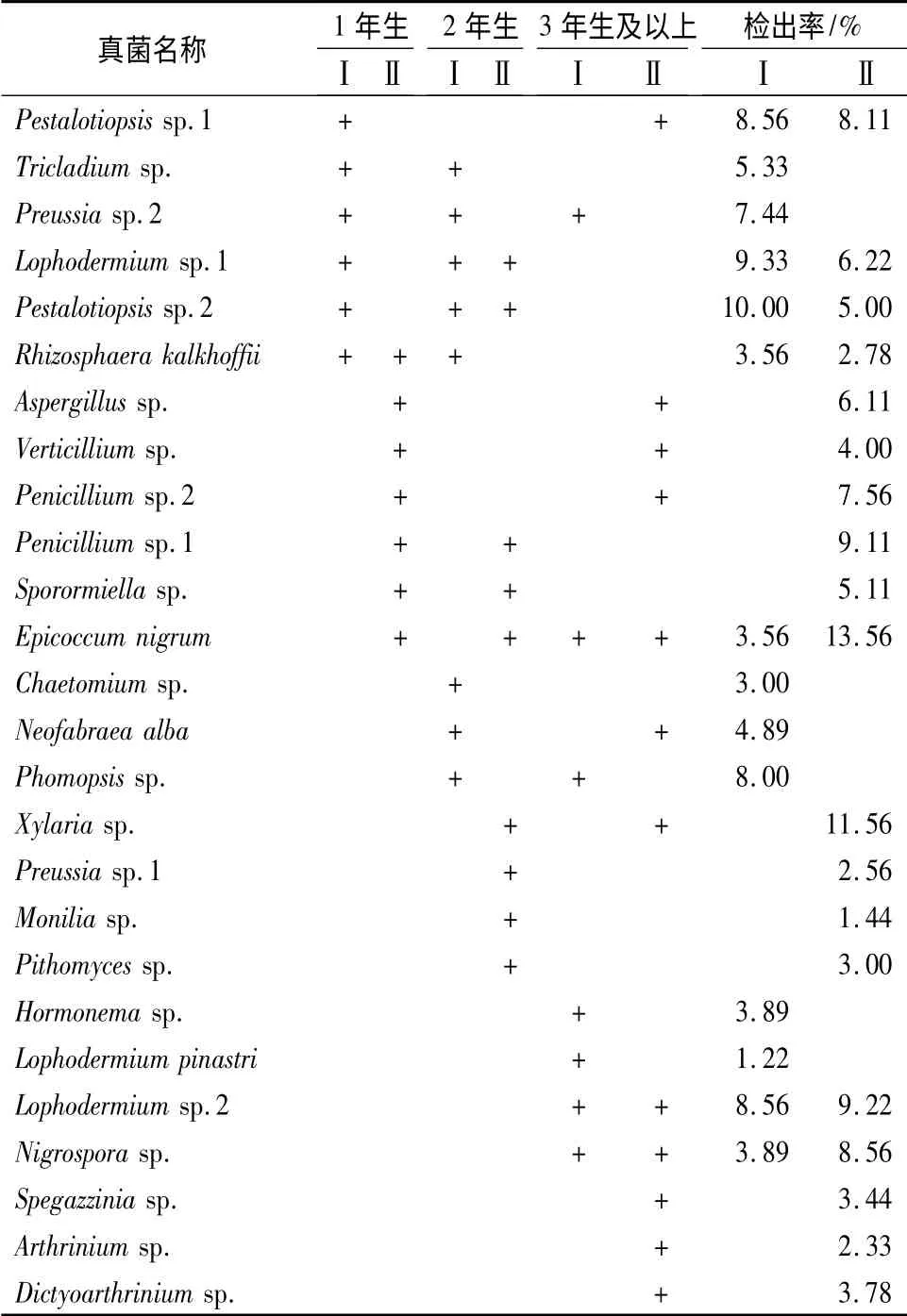
表3 云杉树冠下层针叶叶栖真菌检出率
2.2 不同海拔云杉各冠层叶栖真菌的多样性
从表4可以看出:云杉叶栖真菌的多样性在同海拔处随着冠层改变而有所不同。λ 指数反映出,叶栖真菌在海拔2 480、2 580 m 的云杉上、下冠层的多样性一致,且都高于中冠层。而H'指数所示结果稍有差异,下冠层的多样性最高,上冠层次之,同样是中冠层最低。其他海拔的多样性和均匀度指数,都反映出一致的规律:下冠层的多样性和均匀度最高,中冠层最低,上冠层居于二者之间。
2.3 不同月份云杉各冠层叶栖真菌的多样性
将调查期间6 次取样的每两个月数据进行分析,云杉叶栖真菌多样性分析结果见表5。云杉上、中、下冠层总的叶栖真菌λ 指数和H'指数均在7月份和9月份显示有最高的多样性且均匀度也最高,而相对的11月份和1月份表现出最低的多样性以及均匀度。在上、中、下各冠层单独进行分析时,7月份和9月份的真菌多样性和均匀度也是最高,11月份和1月份最低。
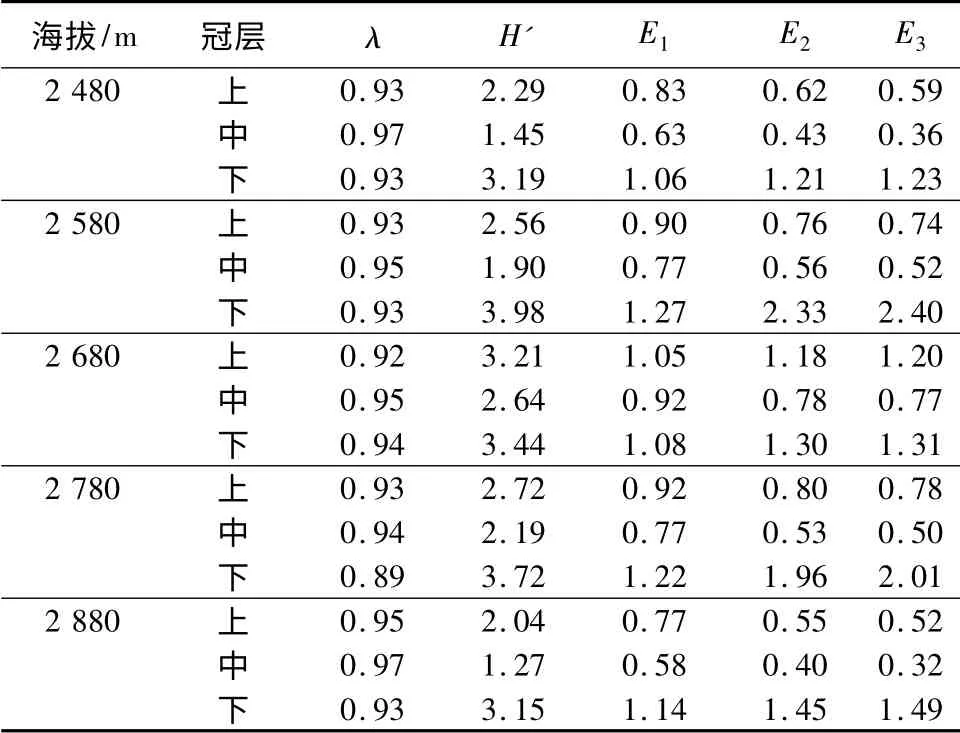
表4 不同海拔云杉叶栖真菌多样性

表5 不同月份云杉叶栖真菌群落多样性
3 结论与讨论
采用2 种针叶表面处理,即消毒组和未消毒组对云杉叶栖真菌进行了组织分离及鉴定,较内生真菌的研究来说更广泛地分析了云杉针叶上真菌的组成和结构,得到的数据也更全面。对云杉叶栖真菌的种群组成分析表明,叶栖真菌种类在不同冠层上的分布以下冠层最多,上冠层次之,而中冠层最少。这一结果与周秀华等[19]略有所不同,相同的是中冠层的真菌种类都是最少的,但和红松叶栖真菌的组成[20]不同,这可能是树种和立地条件等的不同而导致的差异。其次,上冠层和下冠层叶栖真菌的种群数量随着树龄的增长而增长,而中冠层真菌种群的数量在树龄上虽然差异不大,但3年生及以上的叶栖真菌种群数略高于更小叶龄的针叶。这一规律和樟子松针叶上的叶栖真菌随叶龄的增加而增加的结果相一致,且表面消毒后的真菌也明显少于叶表栖真菌[19,21-22]。此外,不同叶龄针叶的真菌数量变化在红松叶栖真菌群落结构组成分析上也有相同结论[20]。
室外调查时发现,每个样点的发病云杉都是树体的中下冠层最为严重。宋瑞清等[23]以红松不同栖息地的树栖真菌分离率作为种群个体参数,采用多样性指数和均匀性指数对树栖真菌进行多样性比较,树栖真菌多样性和均匀度越高,病害发生的可能性越低,呈负相关。对云杉叶栖真菌多样性的分析表明,中冠层的多样性和均匀度都是最低的,下冠层虽然真菌群落的多样性和均匀度指标都较高,但却发病严重。这主要是受病针叶掉落在云杉林下,故下层的针叶更容易接受到病原菌孢子的侵染并定殖而易引起病害的发生。贾云等[24]对红松落针病研究时也提出,病叶落地后产生的子囊孢子向上飞散形成密度递减的梯度。研究表明,7月份和9月份的真菌多样性和均匀度最高,但云杉林在每年的6月份到9月份发病严重,这得考虑到林木病害在森林生态中的分布还受多种环境因素的影响[25]。因此,真菌群落的多样性与均匀度反映了群落的稳定性,也在一定程度上与病害现象相关,有助于对病害的发生进行分析。但这种相关度的高低,是否和病害的发生有必然联系还需要作大量的实践工作加以分析判断。
对云杉叶栖真菌群落的研究表明,散斑壳属(Lophodermium)和拟盘多毛孢属(Pestalotiopsis)在所有叶栖真菌中检出率相对较高,而这两属真菌中的有些种是松落针病和赤枯病的病原菌[26-28]。二郎山林场的树种分布以松树和云杉所占比例最大,并且已有这两种松树病害的发生,故在对松树病害防治的同时,应加强对云杉的施药防病,防止同属植物的病原菌侵染,减少病原菌的繁殖场所。近年来,二郎山地区的云杉幼林出现了严重的针叶枯死脱落症状,以上两属的真菌在病害发生的过程中是否参与致病,还有待研究。同时,在对叶栖真菌的分离中,检出的菌株中有引起云杉叶疫病的根球壳孢菌(Rhizosphaera kalkhoffia),该菌1990年在我国首次报道[13],此后便很少有报道,而国外在早年报道较多[12,29-32]。目前国内报道更多的是由云杉散斑壳菌(Lophodermium picea )引起的云杉落针病[9-10,33-34],但在室外发病针叶上并未发现云杉散斑壳的子囊果。因此,引起二郎山云杉幼林受害的病原菌亟需进一步研究确定。
[1] 黄永清,邵力平,马俊莹.树栖真菌群落多样性研究(Ⅰ):红松芽、叶及短枝栖真菌群落组成分析[J].菌物系统,1997,16(3):182-188.
[2] 郭良栋.内生真菌研究进展[J].菌物系统,2001,20(1):148-152.
[3] Ting A S Y,Meon S,Kadir J,et al.Endophytic microorganisms as potential growth promoters of banana[J].BioControl,2008,53(3):541-553.
[4] Yates I E,Widstrom N W,Bacon C W,et al.Field performance of maize grown from Fusarium verticillioides-inoculated seed[J].Mycopathologia,2005,159(1):65-73.
[5] Morita S,Azuma M,Aoba T,et al.Induced systemic resistance of Chinese cabbage to bacterial leaf spot and Alternaria leaf spot by the root endophytic fungus,Heteroconium chaetospira[J].Journal of General Plant Pathology,2003,69(1):71-75.
[6] Lehtonen P T,Helander M,Siddiqui S A,et al.Endophytic fungus decreases plant virus infections in meadow ryegrass (Lolium pratense)[J].Biol Lett,2006,2(4):620-623.
[7] Sikora R A,Pocasangre L,Felde A,et al.Mutualistic endophytic fungi and in-planta suppressiveness to plant parasitic nematodes[J].Biological Control,2008,46(1):15-23.
[8] 刘增力,方精云,朴世龙.中国冷杉、云杉和落叶松属植物的地理分布[J].地理学报,2002,57(5):577-586.
[9] 孙国忠,王志平,杨庆和,等.云杉落针病防治的研究[J].四川林业科技,1998,19(1):35-37.
[10] 邱书志,蔡斌.云杉落针病防治指标的研究[J].植物保护,2005,31(6):58-62.
[11] 赵震宇.天山林区的三种云杉锈病[J].新疆农业科学,1958(5):190-192.
[12] Livsey S,Barklund P.Lophodermium piceae and Rhizosphaera kalkhoffii in fallen needles of Norway spruce (Picea abies)[J].Eurppean Journal of Forest Pathology,1992,22(4):204-216.
[13] 任玮.云杉叶疫病的研究[J].西南林学院学报,1990,10(2):176-179.
[14] 孙广宇,宗兆锋.植物病理学实验技术[M].北京:中国农业出版社,2002.
[15] 方中达.植病研究方法[M].北京:中国农业出版社,1998.
[16] 杨文博.微生物学实验技术[M].北京:化学工业出版社,2004.
[17] 郭春秋,罗永兰,张志元.几种真菌的诱导产孢试验[J].海南大学学报:自然科学版,2003,21(1):74-77.
[18] 袁秀英,宁瑞些,韩艳洁,等.乌兰布和沙漠固沙树木真菌群落多样性研究[J].内蒙古农业大学学报,2002,23(4):5-8.
[19] 周秀华,崔磊,宋瑞清.樟子松叶栖真菌的群落结构[J].东北林业大学学报,2006,34(2):30-31,56.
[20] 宋瑞清,黄永青.红松叶栖真菌群落结构(Ⅱ)[J].东北林业大学学报,2001,29(1):103-107.
[21] 鞠洪波,吴静.樟子松冬季叶栖真菌群落研究[J].林业科技,2005,30(3):26-28.
[22] 杨传波,黄敬林,李宝年,等.樟子松树栖真菌群落物种多样性及其与病害的关系[J].东北林业大学学报,2004,32(5):94-96.
[23] 宋瑞清,黄永青.红松树栖真菌群落中物种多样性与病害关系(Ⅵ)[J].东北林业大学学报,2001,29(1):123-125.
[24] 贾云,齐乐贤,原戈.红松落针病发生的生态条件分析[J].林业科学,1991,27(1):41-49.
[25] 束庆龙,曹志华,张鑫.树木健康与环境因素的关系分析[J].安徽林业科技,2011,37(1):42-44.
[26] 宋玉双,何秉章,王福生.我国松落针病研究的新进展[J].森林病虫通讯,1994(2):42-46.
[27] 刘应高,潘欣,庄启国,等.四川松树6 种散斑壳菌记述[J].中国森林病虫,2004,23(4):9-12.
[28] 罗孟军,朱天辉.枯斑盘多毛孢菌生物学特性的研究[J].四川林业科技,2001,22(3):15-18.
[29] Diamandis S.“Top-dying”of Norway spruce,Picea abies (L.)Karst.with special reference to Rhizosphaera kalkhoffii Bubák(I):Development of foliar symptoms[J].European Journal of Forest Pathology,1978,8(5/6):337-356.
[30] Diamandis S.“Top-dying”of Norway spruce,Picea abies (L.)Karst.with special reference to Rhizosphaera kalkhoffii Buhák(II):Status of R.kalkhoffii in“top-dying”of Norway spruce[J].European Journal of Forest Pathology,1978,8(5/6):357-361.
[31] Juzwik J.Morphology,cultural-characteristics,and pathogenicity of Rhizosphaera kalkhoffii on Picea spp.in northern minnesota and Wisconsin[J].Plant Disease,1993,77(6):630-634.
[32] Kumi J,Lang K J.The susceptibility of various spruce species to Rhiosphaera kalkhoffii and some cultural characteristics of the fungus in vitro[J].Eurppean Journal of Forest Pathology,1979,9(1):35-46.
[33] 曹秀文.云杉散斑壳菌生物学特性的初步研究[J].甘肃农业大学学报,2000,35(2):176-180.
[34] 孙国忠,杨庆和,赵谦,等.云杉落针病流行规律的研究[J].四川林业科技,1998,19(1):17-22.
猜你喜欢
中国农业信息(2022年1期)2022-05-25 13:31:46
农业机械学报(2021年11期)2021-12-07 05:36:44
大气科学(2021年1期)2021-04-16 07:34:18
现代农村科技(2020年7期)2020-12-19 03:12:08
林业科学(2020年10期)2020-11-30 00:36:56
作文周刊·小学四年级版(2019年8期)2019-04-23 01:04:16
启迪与智慧·教育版(2018年12期)2018-02-26 12:24:06
山西林业科技(2017年4期)2018-01-23 05:45:05
农业环境科学学报(2017年2期)2017-03-20 14:57:37
甘肃林业(2016年2期)2016-11-07 08:56:12
