
基于nLSU 序列探讨丝盖伞属及相关属的系统学关系1)
2014-03-06 03:19:42范宇光图力古尔
东北林业大学学报 2014年3期
关键词:分支
范宇光 图力古尔
(吉林农业大学,长春,130118)
丝盖伞属(Ⅰnocybe (Fr.)Fr.)最早由Fries 于1821年在他的《真菌系统》中采用,记录了现在仍有效的16 种丝盖伞,丝盖伞属当时作为广义的蘑菇属(Agaricus)下的一个“族”[1]。1863年,Fries 在他的《层菌纲专论》中将其提升为属级水平[2]。之后的很长时间,丝盖伞属被认为隶属于丝膜菌科(Cortinariacea)并占据一个独立的族[3],[4]596-677。但近期的分子系统学分析发现丝膜菌科并不是一个单系群[5-6],之后又有研究表明丝盖伞与靴耳科(Crepidotaceae)的种类在系统发育上关系更为接近[7]。关于丝盖伞科的概念及范畴,目前存在不同的观点。Kirk 等[8]认为丝盖伞科包括丝盖伞属在内的13 个属,而美国学者Matheny 却认为丝盖伞科包括丝盖伞属及近期成立的Auritella和Tubariomyces[9-11]。本研究通过nLSU 序列对中国丝盖伞属的系统位置及丝盖伞科的范畴进行讨论。
1 材料与方法
1.1 研究材料
研究中的自测序列来自中国产丝盖伞标本,其他序列从GenBank 中下载。为充分揭示丝盖伞属的系统学位置以及与相关类群间的系统发育关系,选取了丝盖伞科内假脐菇属(Tubaria)、侧火菇属(Pleurofalmmula)、Flammulaster、暗皮伞属(Phaeomarasmius)、靴耳属(Crepidotus)、绒盖菇属(Simocybe)作为参比类群[8]。此外,形态特征或生态特性较为接近的丝膜菌属(Cortinarius)、滑锈伞(Hebeloma)、脆锈伞属(Naucoria)以及球盖菇科的球盖菇属(Stropharia)、鳞伞属(Pholiota)、垂幕菇属(Hypholoma)、田头菇属(Agrocybe)、盔孢菌属(Galerina)、裸盖菇属(Psilocybe)一并列入参比类群。而丝盖伞属的取样则根据广义丝盖伞属的概念,将Matheny 认为的丝盖伞科(Inocybaceae)中新近描述的Auritella、Turariomyces 及其它几个分支均纳入其中。外类群为肉色香蘑(Lepista irina)。研究材料及序列信息详见表1。
1.2 研究方法
基因组DNA 的提取采用改进的CTAB 法,PCR扩增引物为LR0R(ACCCGCTGAACTTAAGC)和LR7(TACTACCACCAAGATCT)[12]。PCR 扩增体系采用25.00 μL体系,具体为:PCRbuffer(+ Mg2+)2.50 μL,dNTP 2.00 μL,上游引物1.00 μL,下游引物1.00 μL,Taq 酶0.12 μL,用水补齐至25.00 μL。PCR 扩增条件为:预变性,95 ℃1 min;35 个循环,95℃30 s,52 ℃1 min,72 ℃1 min;最后72 ℃延伸8 min。测序委托上海生工生物科技公司完成,所用测序仪器为3730xl DNA Analyzer,测序试剂为BigDye terminator v3.1。
序列峰图的查看采用BioEdit 软件,正反向序列的拼接采用DNAMAN 软件,序列的对齐采用ClustalX 1.81 软件,最后用BioEdit 软件对同源序列进行必要的编辑。最后用Mrbayes V3.1.2 软件进行系统发育树的重建。

表1 用于分析的自测序列和下载序列
2 结果与分析
在基于LSU 序列重建的丝盖伞属与相关属的系统发育树中(图1),丝盖伞属、Tubariomyces、Auritella 聚在一枝并得到很好地的支持(100%),这个分支即为美国学者Matheny 认为的丝盖伞科[9-10]。靴耳属、绒盖菇属和侧火菇属聚在一枝(100%),这个分支即为目前认为的靴耳科,它与丝盖伞科形成姊妹群,包含了靴耳属、绒盖菇属和侧火菇属。暗皮伞属、假脐菇属和丝膜菌属的成员各自占据分支并且各分枝均获得了100%的支持。盔孢菌属的分支中包含有一份裸盖菇属的材料,此分支的支持率为92%。鳞伞属、垂幕菇属和田头菇属的成员聚为一支,支持率达90%,此分支即为目前认为的球盖菇科。滑锈伞属与脆锈伞属聚为一支,但并没有得到很好的支持。
3 讨论
关于丝盖伞属的系统位置,传统的分类系统中一致认为丝盖伞属与丝膜菌属和滑锈伞属关系密切,并由这三个属构成丝膜菌科,它们分为占据科下独立的族[3],[4]596-677;此三属均为外生菌根菌,且它们在宏观特征上较为接近。然而,在本研究中发现丝盖伞属、丝膜菌属和滑锈伞属在系统树中并没有体现出紧密的联系,它们占据各自的分支,这与相关研究成果基本一致[5-6]。其中丝盖伞属及Tubariomyces、Auritella 的种类聚为一个强烈支持的分支(100%),而靴耳属、绒盖菇属和侧火菇属聚为一个强烈支持的分支(100%),此分支与丝盖伞的分支形成姊妹群关系。此结果暗示着木生的靴耳属、绒盖菇属和侧火菇属在系统发育上与丝盖伞的成员表现较为接近,尽管它们的生态特性和形态特征均存在较大的差别。同为外生菌根菌丝膜菌属成员的系统树中聚为一个分支,并且获得了较好的支持(100%);滑锈伞属与脆锈伞属虽然聚到一支,但并没有得到很好地支持,此二属与丝盖伞属的关系也相对较远。Matheny 等[7]应用5 个基因片段联合分析表明Hymenogastraceae 的大分支下包括盔孢菌属、裸盖菇属、暗金钱菌属、滑锈伞属、Alnicola、Anamika、Flammula,其中前三个属聚为一小支,后四个属聚为一小支。这样的结构在本研究的系统树中并没有得到明确的体现,这可能与本研究的取样策略或分析方法有关,在此不再深入谈论。无论如何,丝盖伞科的成员在系统树中形成一个明显的单系群,并得到了强烈的支持,这样的结果与早先的观点一致[10],形态与生态特性与丝盖伞相差较大的类群如靴耳属、绒盖菇属和侧火菇属共聚一支并成为丝盖伞科(Inocybaceae sensu Matheny)的姊妹群,形态或生态特性与丝盖伞相接近的类群如滑锈伞属、丝膜菌属等类群反而与丝盖伞的系统发育关系较远。最近,基于rDNA-ITS 序列的球盖菇科系统发育关系分析表明滑锈伞属与库恩菇属(Kuehneromyces)关系密切,且建议将侧火菇属与淡黄鳞伞亚属的部分种合并于丝盖伞科下或单独成立科[13],即本研究中(图1)的靴耳科。
关于丝盖伞科的范畴,不同学者持多种观点[8,10]。根据本研究结果,丝盖伞科在系统树中可以有3 种划分方法。第一种划分所包含的范围最小(图1中的黑色圆形节点),即由丝盖伞属及Tubariomyces、Auritella 构成的分支(100%),此分支中的成员均为外生菌根菌,这样的划分与美国学者Matheny 的观点[10]相一致。第二种划分的范围最大(图1中黑色方形节点),包含了丝盖伞属、Tubariomyces、Auritella、靴耳属、绒盖菇属、侧火菇属、暗皮伞属和假脐菇属等。此分支的成员包括了外生菌根菌和木生菌,子实体形态特征多样化,这样的划分方式与Kirk 等[8]的观点基本一致。第三种划份包含关系密切的丝盖伞分支和靴耳科分支,这样的划分使得丝盖伞科的范畴介于第一种和第二种之间,但科内成员的形态特征及生态特性仍缺乏一致性。以上3种划分所得到的丝盖伞科范畴存在明显的区别,但无论哪种划分方法都较丝盖伞科的最初概念明显扩大。Jülich[14]建立丝盖伞科时虽然包括了2 个属,即Astrosporina 和Ⅰnocybe,事实上当时的丝盖伞科概念与一般认为的丝盖伞属的范畴并无差异。因此,丝盖伞科这一单元在其建立之后并没有得到广泛的认可,直到最近的分子系统学研究开展后才被重新启用[10]。其中,第一种划分中仅保留了除丝盖伞属外的Tubariomyces 和Auritella,无论从系统发育角度还是从形态特征或生态特性角度均较好地保证了丝盖伞科成员的单系性,使得科的界定具有较强的可操作性。但值得注意的是,这样的划分使得科下属级框架及各属的界定问题成为一个新的研究课题。
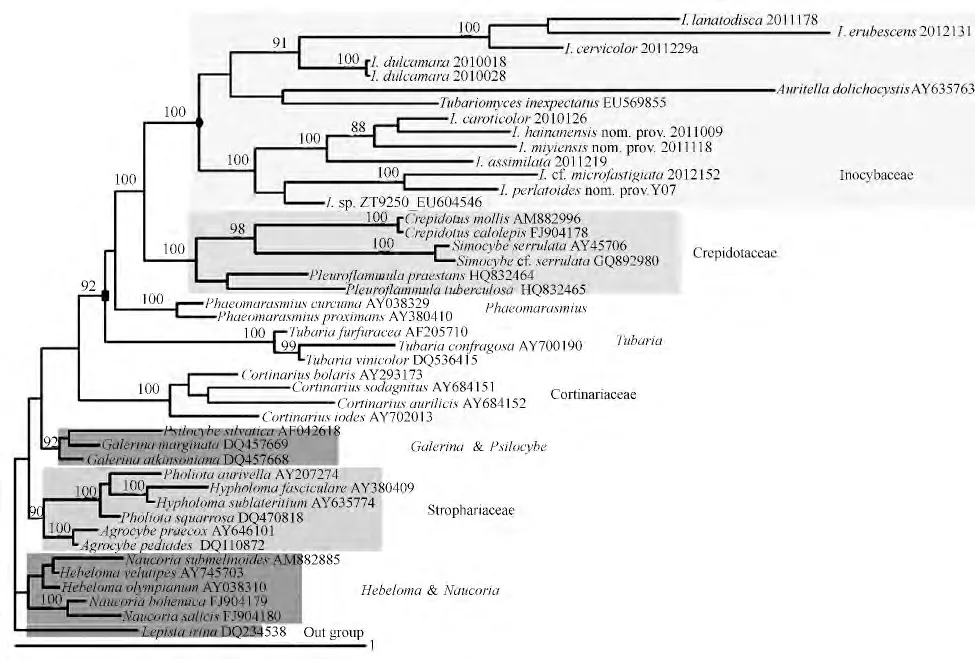
图1 基于nLSU 序列、贝叶斯法重建的丝盖伞属及相关类群系统发育树(后验概率>90%显示在支上)
[1] Fries E.Systema mycologicum:Vol I[M].Lundae:Ex Officina Berlingiana,1821.
[2] Fries E.Monographia hymenomycetum sueciae:Vol II[M].Uppsala:C A LeZer,1863.
[3] Kühner R.Les Hyménomycetes agaricoïdes (Agaricales,Tricholomatales,Pluteales,Russulales:étude générale et classification)[M]//Numéro spécial du Bulletin de la Société Linnéenne de Lyon.Lyon:Société Linnéenne de Lyon,1980.
[4] Singer R.The Agaricales in modern taxonomy[M].4th ed.Koenigstein:Koeltz Scientific Books,1986.
[5] Moncalvo J M,Lutzoni F M,Rehner S A,et al.Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences[J].Systematic Biology,2000,49(2):278-305.
[6] Moncalvo J M,Vilgalys R,Redhead S A,et al.One hundred and seventeen clades of euagarics[J].Molecular Phylogenetics and Evolution,2002,23(3):357-400.
[7] Matheny P B,Curtis J M,Hofstetter V,et al.Major clades of Agaricales:a multilocus phylogenetic overview[J].Mycologia,2006,98(6):982-995.
[8] Kirk P M,Cannon P F,Minter D W,et al.Dictionary of the fungi[M].10th ed.Wallingford:CAB International,2008.
[9] Matheny P B,Bougher N L.The new genus Auritella from Africa and Australia (Inocybaceae,Agaricales):molecular systematics,taxonomy and historical biogeography[J].Mycological Progress,2006,5(1):2-17.
[10] Matheny P B.A phylogenetic classification of the Inocybaceae[J].McIlvainea,2009,18(1):11-20.
[11] Alvarado P,Manjón J L,Matheny P B,et al.Tubariomyces,a new genus of Inocybaceae from the Mediterranean region[J].Mycologia,2010,102(6):1389-1397.
[12] Vilgalys R,Hester M.Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species[J].J Bacteriol,1990,172(8):4238-4246.
[13] 田恩静,图力古尔.基于rDNA-ITS 序列的中国球盖菇科分子系统学[J].东北林业大学学报,2013,41(4):50-54.
[14] Jülich W.Higher taxa of basidiomycetes.Bibliotheca Mycologica[M].Vaduz:J Cramer,1982:85.
猜你喜欢
黑龙江大学自然科学学报(2022年1期)2022-03-29 00:57:32
数学物理学报(2021年4期)2021-08-30 08:27:44
数学物理学报(2020年3期)2020-07-27 01:19:50
学生天地(2019年28期)2019-08-25 08:50:54
经济技术协作信息(2018年28期)2018-11-22 05:27:12
数学物理学报(2018年1期)2018-03-26 08:16:36
数学物理学报(2017年5期)2017-11-23 07:51:10
数学年刊A辑(中文版)(2017年3期)2017-10-30 01:45:48
Chinese Journal of Cancer Research(2017年2期)2017-07-18 11:01:15
数学物理学报(2015年2期)2015-02-28 16:06:43
