
樟树光合特性的季节动态
2014-03-06 03:19罗辅燕陈卫英
东北林业大学学报 2014年9期
罗辅燕 陈卫英
(西南野生动植物资源保护重点实验室(西华师范大学),南充,637009)
樟树(Cinnamonum campora)是亚热带常绿阔叶林的优势树种,同时也是集亚热带植被固碳、药用、香料、化工、城市园林绿化等于一体的多用途树种[1],在维持区域生态环境和经济社会发展等方面发挥着重要的作用。
目前,国内关于樟树光合作用的研究主要集中在环境因素对其光合特性的影响等方面[2-17],而有关樟树春、夏、秋、冬不同季节光合参数特性的动态研究相对较少[18]。
樟树每年春季换叶,老叶逐渐脱落,该生物学特性必定引起樟树叶片光合参数特性的改变。因此,有必要对樟树的光合参数特性进行季节动态分析。目前,有少量基于非直角双曲线模型[19]计算城市樟树人工林不同季节光合参数特性的报道[18],而非直角双曲线模型拟合光响应曲线数据所估算出的最大净光合速率远大于真实值[20-24],其估算出的光饱和点远小于真实值[25],且无法拟合和模拟植物在光饱和点以后随光强的增加而降低的净光合速率[20-22]。直角双曲线修正模型[26]和指数改进模型[27]克服了上述缺点,且指数改进模型的拟合效果略好一些[27]。鉴于基于指数改进模型[27]研究樟树光合参数季节动态变化规律鲜有报道,本研究试图分析和比较樟树春、夏、秋、冬季的光合参数特性,从而为准确估算樟树光合作用和固碳能力提供基础数据,同时从生理生态的角度为常绿阔叶林群落演替和生态系统恢复重建提供理论依据。
1 试验地概况
试验地位于四川省南充市西华师范大学校园内(30°48'43.20″~30°49'33″N,106°04'12″~106°04'56″E),属于中亚热带湿润季风气候区,四季分明。气候特征为春早、夏长、秋短、冬暖。年平均气温17.5℃,年日照时间1 200 h,常年无霜期306 d,年降水量1 100 mm。土壤为红黄壤,肥力中等。
2 材料与方法
2.1 光合作用观测
试验材料为7年生的校园绿化樟树幼树,并于2012年4月6—11日(春季)、8月8—12日(夏季)、10月10—14日(秋季)、12月9—13日(冬季)的08:00—18:30 在西华师范大学校园内选择长势较好的樟树幼树1 ~2 株进行光合作用测定。选取幼树上层完全展开的向阳叶片,采用开路式Li-6400光合测定系统(LI-COR,Inc,Lincoln,NE,USA)进行光合作用日动态和光响应曲线测定,并确保各项光合观测均有3 次以上的重复。测光响应曲线时设定的光强由2 000 μmol·m-2·s-1逐渐递减到0。
2.2 光响应曲线模型简介
本研究基于指数改进模型[27]拟合樟树光响应曲线,进而估算出最大净光合速率(Pmax)、暗呼吸速率(Rd)、表观量子效率(AQ,Y)、光饱和点(LS,P)及补偿点(LC,P)等参数,并用SPSS15.0 统计软件对各光合参数进行多重比较分析。指数改进模型[27]的计算公式如下:
净光合速率公式表达为:

式中:α、β、γ、ξ 为系数;PA,R为光照强度。
当PA,R=0 时,暗呼吸速率(Rd)表示如下:

当Pn=0 时,可推导出光补偿点计算公式:
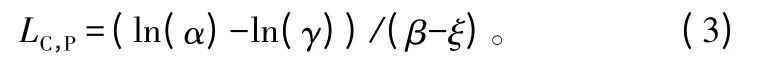
表观量子效率:

光饱和点计算公式:

光饱和点时的净光合速率为最大净光合速率

3 结果与分析
3.1 樟树光合作用日动态
从图1可以看出,樟树春、夏、秋季的净光合速率日动态均呈现“双峰型”曲线,而在冬季呈现“单峰型”曲线。樟树夏季净光合速率在11:00 达到最大,为9.06 μmol·m-2·s-1,随后叶片净光合速率逐渐下降出现“午休”现象,到15:00 点出现第二个高峰,为8.40 μmol·m-2·s-1,之后净光合速率又呈现下降趋势。春季和秋季的日动态规律较为相似,12:00 净光合速率达到最大,分别为6.91、7.23 μmol·m-2·s-1,“午休”现象后在15:00 点光合速率出现第二个高峰,分别为6.50、6.90 μmol·m-2·s-1。在冬季,樟树净光合速率随光照的增加而增加,在12:00 达到最大,为5.60 μmol·m-2·s-1,之后呈逐渐下降趋势。多重比较分析发现,夏季,樟树叶片日最大净光合速率和日均值最大,分别为9.06、7.03 μmol·m-2·s-1;冬季最小(P<0.05),分别为5.60、4.32 μmol·m-2·s-1;春、秋季两者没有显著差异。

图1 樟树净光合速率日动态
3.2 樟树光合参数季节动态变化
基于指数改进模型[27]和光响应曲线数据来估算樟树各季节的光合参数。樟树春、夏、秋、冬季具有较高的光饱和点,变化幅度为1 267.31 ~1 625.70 μmol·m-2·s-1,具有明显的季节变化。其中,夏季光饱和点最大,其次是春、秋季,春季和秋季光饱和点两者没有显著差异,冬季最小。樟树光补偿点也较高,变化范围为48.70 ~63.44 μmol·m-2·s-1,其季节动态变化规律为:夏季光补偿点最大,春、秋、冬季没有显著差异(表1)。
樟树光合能力具有“单峰”或者“双峰”形状的日动态变化,同时也具有季节动态特征。樟树春、夏、秋、冬季的最大净光合速率变化范围分别为5.31~9.19 μmol·m-2·s-1,具有明显的季节动态。其中,夏季樟树最大净光合速率最大,其次是春、秋季,春季和秋季两者没有显著差异,冬季最小。四季中,樟树夏季暗呼吸速率最大,为2.53 μmol·m-2·s-1,春、秋、冬三季没有显著差异,约为1.40 μmol·m-2·s-1。
四季中,樟树春、秋、冬季具有较高的表观光量子利用效率,均在0.060 左右,而夏季表观光量子利用效率相对较低,为0.046。其中,春、秋、冬季表观 光量子利用效率没有显著差异。

表1 樟树春、夏、秋、冬季光合参数
4 结论与讨论
在自然生长环境中,植物经历着环境因子在时空上的日变化和季节变化,从而引起叶片净光合速率的日动态变化和季节变化。有研究指出,樟树夏、冬季均出现“单峰型”曲线[28],春季樟树叶片的净光合速率日变化均呈“单峰”型,未出现光合“午休”现象[8]。黄志宏等[18]得出春、夏、秋、冬季四季的净光合速率日动态呈现出“双峰型”曲线。以上结论和本研究结果有所差异,本研究结果是,樟树春、夏、秋季的净日动态均呈现“双峰型”曲线,而在冬季呈现“单峰型”曲线。“午休”现象是否出现,主要取决于当时的光强、温度及植物水分亏缺等情况,是综合因素共同作用的结果。光强逐渐增加到某一阈值,植物叶片蒸腾速率加剧,净光合速率也缓慢增加至最大值。当光强超过这一阈值之后,长时间的高温、高光和水分亏缺等环境因子胁迫会引起叶片气孔关闭和光合活性的降低,从而产生“午休”现象,这也是植物对不利环境的一种生理生态适应和策略。因此,在实际苗木管理中,应该减少光照、水分、温度等环境因子对林木生长胁迫,在强光、高温、低湿时对苗木进行适当的遮阴、喷灌处理,改善苗木的微环境,减缓或者消除“午休”现象的影响,从而使林木处于较高水平的净光合速率和生长速率。
进行植物光合生理特征研究是揭示不同植物对其生存环境生态适应性机制[29]和高效培育管理的有效途径。一般来说,植物的光饱和点和光补偿点低,其耐阴性强,而阳性植物则具有较高的光饱和点和光补偿点。蒋高明[30]认为阴性植物的光补偿点小于20 μmol·m-2·s-1,光饱和点为500 ~1 000 μmol·m-2·s-1或更低;阳性植物的光补偿点为50~100 μmol·m-2·s-1,光饱和点为1 600~2 000 μmol·m-2·s-1或更高。强阳性树种樟树四季具有较高的光饱和点和光补偿点,且具有明显的季节动态。其中,夏季最大,分别为1 625.70、63.44 μmol·m-2·s-1。然而,有研究[11]基于非直角双曲线模型估算出夏季(7月份)的幼树光饱和点和光补偿点偏低,分别为1 250.00、40.60 μmol·m-2·s-1。通过对该文献中的光响应曲线数据进行反算得出,这主要是由于非直角双曲线模型估算出的光饱和点远小于真实值[25]等缺陷引起的。另有研究[14]基于非直角双曲线模型估算的夏季(8月份)樟树光饱和点和光补偿点更低,仅分别为748.00、4.70 μmol·m-2·s-1,这显然不是阳性树种该有的光适应特性。可见,光合模型的选择及其对光合参数的准确估算,对研究植物光合特性具有极其重要的意义。
植物暗呼吸通过消耗光合产物进行糖酵解释放大量能量,且在0 ~40 ℃时植物的呼吸强度随温度上升而提高。暗呼吸速率的大小表征着植物的生理活动强弱。本研究得出,四季中樟树夏季暗呼吸速率最大,高达2.53 μmol·m-2·s-1。这是由于夏季温度高,暗呼吸所需要的光合底物充足,叶片生理活动中的酶活性强,进而导致夏季樟树暗呼吸最大。
相比亚热带常绿阔叶乡土树种米槠、丝栗栲,樟树具有较高的最大净光合速率,均在5 μmol·m-2·s-1以上。樟树常作为常绿阔叶林恢复和演替初期的先锋树种而存在,这是由于其春、夏、秋、冬季均具有较好的光合能力和生长速率。这就为其在群落演替之初就能较早的取得优势地位并在群落演替中保持其优势地位奠定了基础。
另外,目前不少研究[14,17]基于某一个季节的光合观测数据来描述樟树的光合特性,而樟树光合特性具有较强的季节性。因此,不能简单地用某一时段的光合数据来表征和比较植物光合特性。
[1] 郝日明,魏宏图.紫金山森林植被性质与常绿落叶阔叶混交林重建可能性的探讨[J].植物生态学报,1999,23(2):13-20.
[2] 赵平,曾小平,彭少麟.植被恢复树种在不同实验光环境下叶片气体交换的生态适应特点[J].生态学杂志,2003,22(3):1-8.
[3] 廖凌娟,曹洪麟,吴林芳,等.交通尾气污染对城市绿化乡土树种的影响:气孔、光合生理的响应[J].热带亚热带植物学报,2011,19(5 ):446-452.
[4] 刘志发,申卫军,温达志.南澳岛光裸地和次生林下6 种乡土树幼苗生长与光合特征[J].生态环境学报,2009,18(2):621-628.
[5] 史建君.樟树对14CO2的吸收和积累[J].中国环境科学,2010,30(12):1690-1694.
[6] 周帅,林富平,王玉魁,等.樟树幼苗机械损伤叶片对挥发性有机化合物及叶绿素荧光参数的影响[J].植物生态学报,2012,36(7):671-680.
[7] 刘海星,张德顺,商侃侃,等.不同黄化程度樟树叶片的生理生化特性[J].浙江农学院学报,2009,26(4):479-484.
[8] 李全发,王宝娟.樟树不同叶龄叶片的光合特性研究[J].福建林业科技,2011,38(4):78-80.
[9] 闫文德.樟树生理生态特性及其林分生物产量的研究[D].长沙:中南林学院,2003.
[10] 田大伦,罗勇,项文化,等.樟树幼树光合特性及其对CO2浓度和温度升高的响应[J].林业科学,2004,40(5):88-92.
[11] 刘茜.樟树叶片蒸腾特性及其与生理生态因子关系分析[J].中南林业科技大学学报,2009,29(1):1-5.
[12] 闫明,钟章成.铝胁迫和丛枝菌根真菌对樟树幼苗生长的影响[J].北京林业大学学报,2007,29(4):161-165.
[13] 陈卓梅,陈英旭,杜国坚.二氧化氮对樟树幼苗生长及光合作用的影响[J].应用生态学报,2009,20(7):1650-1656.
[14] 潘文,张卫强,甘先华,等.SO2胁迫对园林植物幼苗光合生理特征的影响[J].中国水土保持科学,2013,11(1):82-87.
[15] 付晓萍,田大伦,闫文德.模拟酸雨对樟树光合日变化的影响[J].中南林学院学报,2006,26(6):38-43.
[16] 田大伦,付晓萍,方晰,等.模拟酸雨对樟树幼苗光合特性的影响[J].林业科学,2007,43(8):29-35.
[17] 郭淑红,薛立,杨振意,等.水淹胁迫对4 种园林绿化树种幼苗气体交换参数的影响[J].华南农业大学学报,2013,34(4):548-552.
[18] 黄志宏,田大伦,闫文德,等.城市樟树人工林冠层光合作用的时空特征[J].中南林业科技大学学报,2011,31(1):38-46.
[19] Kubiske M E,Pregitzer K S.Effects of elevated CO2and light availability on the photosynthetic light response of trees of contrasting shade tolerance[J].Tree Physiology,1996,16(3):351-358.
[20] Evans J G,Jakonbsen I,Ogren E.Photosynthetic light-response curves.2.gradients of light absorption and photosynthetic capacity[J].Planta,1993,189(2):191-200.
[21] Kyei-Boahen S,Lada R,Astatkie T,et al.Photosynthetic response of carrots to varying irradiances[J].Photosynthetica,2003,41(2):301-305.
[22] Yu Qiang,Zhang Yongqiang,Liu Yunfen,et al.Simulation of the stomatal conductance of winter wheat in response to light,temperature and CO2changes[J].Annals of Botany,2004,93(4):435-441.
[23] Leakey A B,Uribelarrea M,Ainsworth E A,et al.Photosynthesis,productivity,and yield of maize are not affected by open-air elevation of CO2,concentration in the absence of drought[J].Plant Physiology,2006,140(2):779-790.
[24] Posada J M,Lechowicz M J,Kitajima K.Optimal photosynthetic use of light by tropical tree crowns achieved by adjustment of individual leaf angles and Nitrogen content[J].Annals of Botany,2009,103(5):795-805.
[25] 高俊,孟平,吴斌,等.杏—丹参林药复合系统中丹参光合和蒸腾特性的研究[J].北京林业大学学报,2006,28(2):64-67.
[26] 叶子飘,于强.冬小麦旗叶光合速率对光强度和CO2浓度的响应[J].扬州大学学报:农业与生命科学版,2008,29(3):33-37.
[27] 陈卫英,陈真勇,罗辅燕,等.光响应曲线的指数改进模型与常用模型比较[J].植物生态学报,2012,36(12):1277-1285.
[28] 黄娟,吴彤,孔国辉,等.油页岩废渣地12 种木本植物光合作用的季节变化[J].植物生态学报,2006,30(4):666-674.
[29] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002:108-113.
[30] 蒋高明.植物生理生态学[M].北京:高等教育出版社,2004:59-62.
猜你喜欢
新作文·小学低年级版(2022年4期)2022-08-30
江苏农业科学(2020年12期)2020-07-20
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
北方音乐(2019年6期)2019-07-16
现代园艺(2018年3期)2018-02-10
新课程·下旬(2017年7期)2017-08-14
西北林学院学报(2015年5期)2015-01-03
新课程·中旬(2014年10期)2014-12-26
天然产物研究与开发(2014年6期)2014-04-27
植物营养与肥料学报(2011年2期)2011-10-26
