
公路对天山马鹿(Cervus elaphus songaricus)种内遗传分化的影响
2014-03-02 11:24周璨林艾萨江图合提阿米拉阿布来提日沙来提吐尔地艾斯卡尔买买提马合木提哈力克
中国草食动物科学 2014年6期
周璨林,艾萨江·图合提,阿米拉·阿布来提,日沙来提·吐尔地,艾斯卡尔·买买提,马合木提·哈力克
(1.新疆大学生命科学与技术学院资源生物研究所,乌鲁木齐 830046;2.新疆医科大学基础医学院,乌鲁木齐 830011)
遗传育种
公路对天山马鹿(Cervus elaphus songaricus)种内遗传分化的影响
周璨林1,2,艾萨江·图合提1,阿米拉·阿布来提1,日沙来提·吐尔地1,艾斯卡尔·买买提1,马合木提·哈力克1
(1.新疆大学生命科学与技术学院资源生物研究所,乌鲁木齐 830046;2.新疆医科大学基础医学院,乌鲁木齐 830011)
为研究公路对大型哺乳动物造成的影响,分别采集新疆省道110、203、303公路两侧的天山马鹿粪便样本,采用5个微卫星位点进行个体识别后利用Arlequin 3.11进行遗传距离、基因多样性、杂合度等测定分析。结果表明:5个微卫星均具有很好的多态性,5地区中松树塘与寒气沟、白石头马鹿的遗传距离最远,松树塘马鹿的基因杂合度、基因型数量均最低;小西沟与白杨沟马鹿之间遗传距离为负值,其基因杂合度、基因型无显著性差异(P>0.05)。说明公路对栖息地的分割状态不同,会导致种群的遗传情况改变。当公路仅深入栖息地没有将栖息地割裂时,公路两旁种群无差异;当栖息地被公路完全隔绝,则该栖息地内动物的基因杂合度、基因型数量降低;使用中的公路对临近种群的隔绝效果强于废弃旧公路。
公路;天山马鹿;遗传分化;栖息地破碎化
道路在现在社会中发挥着重要的作用,但是对于生态系统却起了负面作用,比如污染、噪音、打搅其生活以及破坏家域。更为严重的是公路存在的潜在效应,栖息地破碎会威胁道路两旁的动物生存,加速其灭绝。栖息地被公路或者其他人工建筑分成了许多破碎的小块,这些公路会减少基因流,其结果将会使得种群被分割并使其数量减少。这样的种群会进一步受到近亲交配的威胁,并丧失其遗传多样性。
为解决这些问题,世界各国特别是欧洲、北美、澳大利亚等国纷纷建立了野生动物通道。世界上第一条野生动物通道建立于上世纪50年代美国的佛罗里达州,中国的第一条带有野生动物通道的公路是云南的小勐养高速公路[1]。中国的大多数公路设计没有考虑野生动物通道的建设,公路两旁仅有小型动物可以通过公路的排水沟等渠道穿越公路[2],大型哺乳动物由于需要至少宽7 m高2.4 m以上的专用通道,因而无法安全通过公路。这些大型哺乳动物由于受到诸如体型庞大、活动区域面积大、低种群密度以及种群增长缓慢等内在的生物属性影响,因此对于栖息地的丧失以及破碎化特别敏感[3-4]。
天山马鹿就是这样的一种在生态系统中发挥着重要作用的大型哺乳动物,它是国家二级保护动物,并且被国际自然保护联盟(the international union for the conservation ofnature,IUCN)列为LC级。天山马鹿是马鹿的一个亚种,广泛分布于天山山脉,它不仅受到种群数量小和捕猎的威胁,近亲交配以及栖息地破碎化也是威胁其生存的重要因素。一些研究者已经对于栖息地破碎化的后果进行了研究,比如食性选择、遗传多样性丧失等[5]。但是道路对于家域大小的影响大于其他自然因素。尽管欧洲第一个研究道路对大型动物遗传效应的文章发表于2007年[6],我国最早的对动物遗传效应的文章发表于2006[7],但是至今为止没有关于道路对大型哺乳动物遗传效应的研究。
天山马鹿的栖息地被许多道路以及其他人造障碍阻隔。已有的证据表明,道路对有蹄类动物的阻碍作用会随着道路等级的变化而变化,高速公路的阻碍效力为1,也就是完全阻挡有蹄类动物的迁徙与基因交流。国道和省道的阻碍效力分别是0.71~0.79和0.43~0.59[8]。这些数据都预示着在省道两旁的有蹄类动物的基因交流存在着阻隔效应。但是所有的研究均没有考虑到省道与生境之间的关系。本研究以天山马鹿为例(C.elaphus songaricus)研究不同道路对遗传效应的影响。
1 材料与方法
1.1 研究地点交通情况
天山山脉的哈密地区部分被省道203、303、302将马鹿栖息地分割为3个区域,从东到西依次为寒气沟、松树塘、白石头,如图1所示。3条公路当中,省道203为旧公路,在省道303通车后来往车辆较少,但是野生天山马鹿天性胆小,一般不会到公路水泥等人工硬地,且习惯于与人类保持一定距离,因此依然造成邻近区域间马鹿的迁徙障碍;而省道303为新公路,2003年建成通车后车流量较密集,现已成为哈密向北过天山的主要通路;省道302是哈密与伊吾县之间的唯一通路,往来车辆较多。寒气沟地区由于地处天山山脉的东部,受到省道303、302的阻挡迁徙向西迁徙困难;松树塘地区东西两侧分别是省道203、303;白石头地区地处哈密天山山区西部,与天山山脉主体相连。天山山脉的乌鲁木齐地区被国道216分割,在国道的西侧又被省道109、110分为3部分,其中靠近西部的2个地区分别是白杨沟(NB)和小西沟(NX),两个栖息地被省道109分割,该路主要为旅游及当地牧民交通用,因此并未将山脉分割,如图1所示。
1.2 样品收集与DNA提取
本研究一共有5个采样点,小渠子(NX)和白杨沟(NB)之间的省道并没有将生境完全分割,在南部仍有林地相连;白石头(BST)和松树塘(SST)之间被旧的废弃省道完全分割;松树塘(SST)和寒气沟(HQG)之间被正在使用的省道完全分割。
从2010至2011年,分别在天山山脉的哈密、乌鲁木齐山区收集马鹿粪便样本,采集地点如图1所示,每一个采集地收集的样品数量如表1所示。粪便样品使用纯乙醇保存在-80℃冰箱中。使用日沙来提[9]改进过的CTAB裂解法提取总DNA,提取结果和纯度使用1.2%的凝胶电泳检测。

图1 天山马鹿样品采集地点(实线表示公路S110,S109,G216S203,S303以及S302,虚线表示马鹿分布区)

表1 马鹿粪便样品来源
1.3 微卫星扩增与个体识别
选择马鹿遗传多样性分析中使用的具较高多态性的5个微卫星位点(BM888,BM4208,BM5004,BM6438,BM6506)[10]如表2所示。PCR扩增体系为50 μL,反应体系中含50 ngDNA,每条引物终浓度2 pM,25 μL的2× PrimeSTAR MaxPremix(TaKaRa Bio Inc.,Dalian,China)。扩增条件:98℃预变性1min后30个循环,98℃解链10s,使用表2所示的退火温度退火10 s,72℃延伸5 s,最后72℃30 s延伸,4℃保存。PCR的扩增产物利用聚丙烯凝胶电泳检测,EB显色人工辨认微卫星大小。利用软件Microsoft Excel,寻找数据中相匹配的基因型,进行个体识别。

表2 5个微卫星引物序列
1.4 个体识别
PCR的扩增产物利用聚丙烯凝胶电泳检测,EB显色,利用肉眼观察判断同一个体。使用POPgene 3.2软件计算2个马鹿群体的5个微卫星座位哈代-温伯格平衡值(HW),通过EXCEL microsatellite tool kit和cervus 3.0软件寻找数据中相匹配的基因型。判断不同样品来自于同一个个体的原则:①所有座位上的基因型都相同;②只有1个座位上的1个等位基因存在差异。同时使用性别鉴定作为个体识别的辅助手段。
1.5 通过SRY基因进行性别鉴定
本实验使用2对引物对粪便DNA进行复合扩增:引物 SRY12(5'-CTTCATTGTGTGGTCTCGTG-3'与 5'-CGGGTATTTGTCTCGGTGTA-3')扩增Y染色体SRY区域片段;引物BMC1009作为反应阳性内对照扩增常染色体微卫星位点,以防止由于实验失败而错误判别。引物由上海生工生物工程技术服务有限公司(Sangon)合成,并用灭菌双蒸水配制成浓度为100 μmol/L的储存液备用。反应体系:10×PCR buffer 3 μL,dNTP 3 μL,两对引物上下游各1 μL,模板DNA 2 μL,Taq DNA酶1 μL,最后加入双蒸灭菌水12 μL使总体积为25 μL。反应条件:95℃预变性10min,95℃变性30s,61℃退火45s,72℃延伸1 min,40个循环,最后72℃延伸10 min,4℃保存。
1.6 数据分析
将不同个体的微卫星数据电泳后的片段大小输入Arlequin 3.11软件,数据类型设为microsat,计算遗传结构时将Markov的计算次数设为100 000,检验次数设为10 000,显著性差异定位0.05;计算Hard-Weinberg平衡,使用的Markov的计算次数设为100 000,检验次数设为10 000,最后进行成对计算。使用POPgene 3.2计算Ne。使用FSTAT2软件计算种群之间的Fst值。参照Botstein等的方法计算多态性信息(PIC,polymorphism information content):

式中n代表等位基因数。Pi和Pj分别表示第i和第j个等位基因出现的频率。
使用2种方法来描述空间的遗传变异模式。首先,Mantel测试评估地理距离对种群分化的影响。地理距离采样点的坐标计算基于抽样的近似中心地区。使用R-PACKAGE模块计算地理距离之间的相关性和遗传距离[Fst/(1-Fst)],使用10 000次随机化确定统计学差异关系。
使用Monmonier方法确定遗传边界,即该区域对种群的遗传差异是最高的,这个方法适用于检测突变遗传改变。运行BARRIER version 2.2程序,应用Monmonier的最大差分算法以后,使用Delaunay三角测量法分别对哈密以及乌鲁木齐的种群进行连接。
2 结果与分析
2.1 马鹿种群的遗传多样性
本研究结果显示,449份粪便来自251个个体,其中来自白杨沟的127份粪便属于60个个体;来自小渠子的128份粪便属于51个个体;来自松树塘的41份粪便属于32个个体;来自白石头的87份粪便属于66个个体;来自寒气沟的66份粪便属于42个个体。雌雄比为2.24:1。
使用Arlequin 3.11软件分析5个微卫星位点在各地天山马鹿的遗传距离,两两比较获得Fst结果如表3所示,各地的遗传距离的P值除乌鲁木齐的白杨沟、小渠子之间以外均小于0.05,差异显著。使用Markov链式检验方法,分别计算白杨沟、小渠子、松树塘、白石头、寒气沟5地区5个微卫星位点的Hardy-Weinberg平衡,结果表明5个位点在马鹿种群中的期望杂合度(He)的变化范围为0.69~0.72,平均0.71;杂合度的观测值(Ho)变化范围为0.75~0.91,平均0.83。5个多态位点中多态信息含量0.47~0.75,其中BM4208位点多态信息最多,平均值为0.69。有效等位基因数(Ne)2.23~4.46,Shannon’s系数(I)0.96~1.55(表4)。
选择Arlequin3.11软件计算等位基因差异数(Fst)作为衡量遗传距离的单位,其结果表明,除了乌鲁木齐地区的白杨沟(NB)与小渠子(NX)种群之间的遗传差异为负值外,其余所有的地区之间Fst差异显著,最大的Fst来自于哈密地区的松树塘(SST)和寒气沟(HQG)之间,如表3和图2所示。

表3 各天山马鹿群体间遗传距离的比较
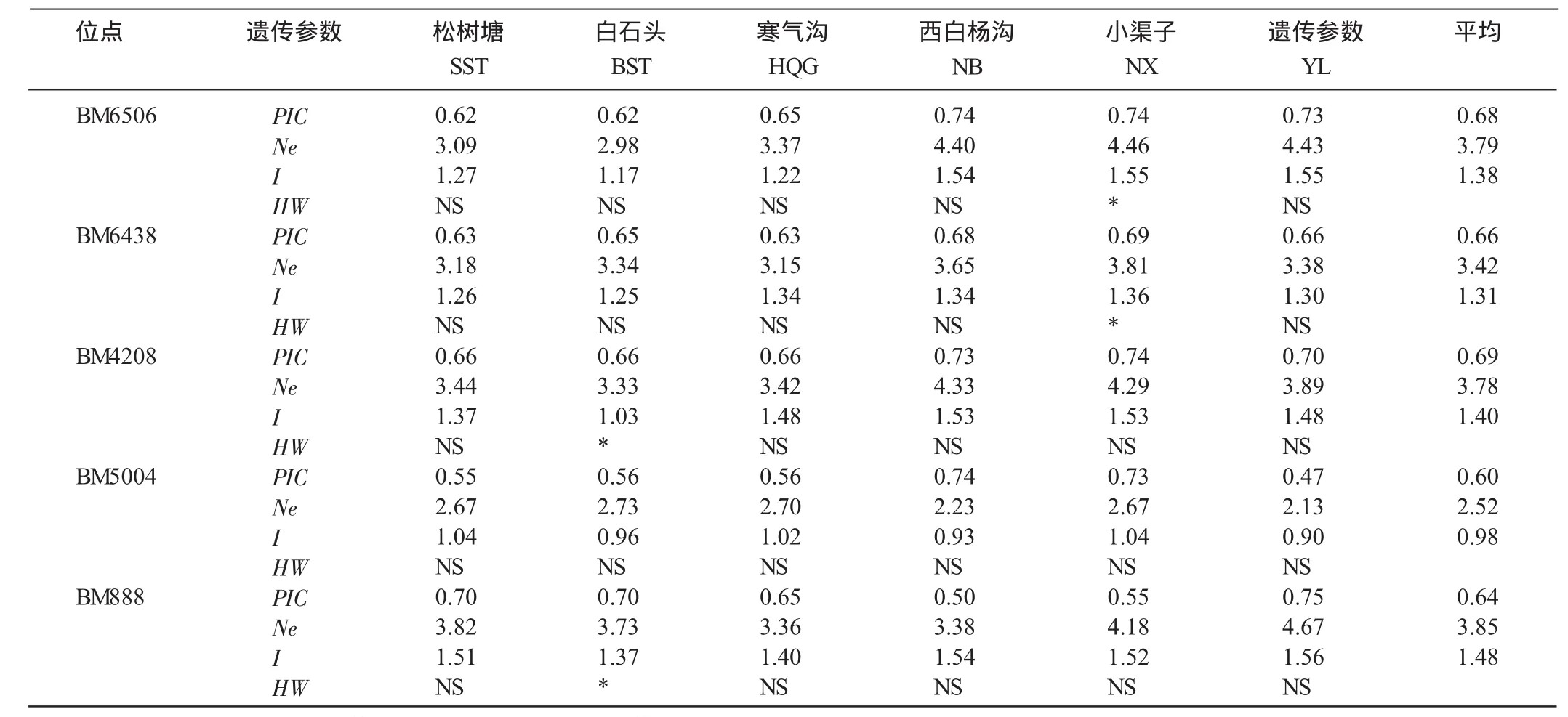
表4 5个微卫星位点在天山马鹿两个种群中的遗传参数

图2 5个微卫星位点在天山马鹿两个种群中的遗传参数
3 讨论
传统的野生动物样品采集方式多为捕捉、猎杀等方式。为减少样品采集时对野生马鹿的伤害,不打扰其野外生活,本研究采用非损伤性采样法,利用新鲜粪便提取表面脱落肠道细胞中的DNA,因此样品存在多次采样的情况。为排除重复取样对本研究的干扰,首先使用5个微卫星引物 BM888、BM4208、BM5004、BM6438、BM6506进行个体识别,判断不同样品来自于同一个个体的原则是:①所有座位上的基因型都相同;②只有一个座位上的一个等位基因存在差异[11]。
天山山脉是新疆的主要山脉之一,被分为东天山与西天山。东天山山脉仅有的两条穿山公路均为省道且都在哈密境内,二者距离较近;西天山山脉的穿山公路均为国道,省道均未穿越天山山脉。本研究所选的天山山脉乌鲁木齐地区以及哈密地区分别属于东天山与西天山,而所分析的公路依据对栖息地的割裂方式以及使用情况,分为如下几类:省道110属于对栖息地的入侵,会对马鹿的生活造成干扰,但不会完全割裂小渠子与白杨沟之间的基因交流,没有将两地生境分开;省道203属于旧公路,虽然穿过天山山脉,完全割裂了松树塘与白石头之间的基因交流,曾经被人类使用,但现在几乎不再使用,因此两地之间的马鹿曾经完全隔离,但是现在两地之间存在相互交流的可能性;省道303为新建公路,其中的南北段穿过天山山脉,完全将寒气沟与松树塘分开,因此本研究几乎涉及了各类影响栖息地类型的公路。
松树塘地区东西两侧分别被两条省道阻隔,虽然和其余两地区紧邻,地理距离均在5 km以内,但是松树塘与其他两地的遗传距离差异极显著,是哈密地区3地间遗传距离的最大值和最小距离。这是由于松树塘地区与寒气沟地区之间是新省道303,该路段地势相对就省道平坦,且海拔低无盘山路段,一年四季车流不断,特别秋季是煤炭、牧草等运输的主要时段,而马鹿的发情期是10月,公路车流使得两地区之间难以相互交配,而省道203则车流稀疏,松树塘地区与白石头地区间的马鹿种群基因交流相对方便。松树塘地区的马鹿基因型平均值31.6 ,杂合度0.76,均为研究区域中的最小值,这是由于该地区难以与邻近地区的马鹿进行基因交流,加之地形较小(约43 km2),种群数量十分有限,因此更易受到遗传漂变的影响丧失遗传多样度,并且由于种群数量少,近亲交配风险增高,导致杂合度降低,基因型丢失。白石头地区与松树塘地区紧邻,与寒气沟地区地理距离较远,从遗传距离看,白石头地区马鹿与松树塘地区马鹿遗传距离较寒气沟地区马鹿之间近。表面上看似乎是遗传距离是地理距离的反映,但是考虑到寒气沟地区的马鹿与邻近的松树塘马鹿遗传距离远,而与地理较远的白石头地区马鹿遗传距离相对较近,那么白石头地区与寒气沟地区之间的基因交流就不是通过松树塘地区作为中转实现的,它们之间还可以通过省道以西地区进行交流。省道302对省道303有分流作用,因此马鹿可能是以302以西地区为中介,在白石头地区与寒气沟地区间偶有交流。从基因型数量来看,白石头地区马鹿是63.6,为所有研究区域中最高值,杂合度的观测值则低于寒气沟地区马鹿,但是二者没有统计学差异。这种基因型数量与高杂合度较丰富的原因,是由于白石头地区西侧和东天山山脉主体相连,白石头地区马鹿可以在东天山的广大区域内自由迁徙造成的。寒气沟地区位于哈密的最东侧,处于整个东天山山脉的末端,虽然整个寒气沟地区内部再无公路将其分割,但是由于总面积远远小于白石头地区及以西地区,因此该地区马鹿杂合度的观测值与白石头地区相当,但是基因型数量依然低于白石头地区。
对于天山山脉乌鲁木齐山区的小渠子、白杨沟两地的遗传分析显示,两者之间的遗传距离为负值,说明两地之间的马鹿实际为同一种群。小渠子、白杨沟两地之间是省道110,但是该公路并未穿过天山山脉,因此两地之间的天山马鹿可以自由往来,遗传分析结果也印证了这种地理结构。
4 结论
如果公路完全分割栖息地,则以马鹿为代表的大型哺乳动物在公路两侧的相互基因交流就会受阻,如果其中一部分栖息地较小则更容易失去遗传多样性,容易灭绝。因此,在未来的道路施工中应当增加对栖息地保护,对已有公路进行改造,以方便基因交流,这样才能更好地保护其遗传多样性,避免种群的灭绝。
[1]Qiu L,Feng Z J.Effects of traffic during daytime and other human activities on the migration of Tibetan Antelope along the Qinghai-Tibet highway,Qinghai-Tibet Plateau[J].Acta Zoologica Sinica,2004,50(4):669-674.
[2]RodriguezA,Crema G,Delibes M.Factors affectingcrossingof red foxes and wildcats through nonwildlife passages across a highspeed railway[J].Ecography,1997,20(3):287-294.
[3]Crooks K R.Relative sensitivities of mammalian carnivores to habitat fragmentation[J].ConservBiol,2002,16:488-502.
[4]CardilloM,Mace G M,Jones K E,et al.Multiple causes of high extinctionriskinlargemammalspecies[J].Science,2005,309:1239-1241.
[5]Ochoa A,J Gasca,G J Ceballos,et al.Spatiotemporal population genetics of the endangered Perote ground squirrel(Xerospermophilus perotensis)in a fragmented landscape[J].Journal of Mammalogy,2012,93(4):1061-1074.
[6]Ralph K,Karin E H,Otto H,et al.Genetic effect of transportation infrastructure on Roe Deer populations(Capreolus capreolus)[J].Journal ofHeredity,2007,98(1):13-22.
[7]周乐,殷宝法,杨生妹,等.青藏公路对高原鼠兔种内遗传分化的影响[J].生态学报,2006,26(11):3572-3777.
[8]Zhou J J.The highway's influence on ungulate in Heilongjiang province[D].Harbin:Northeast ForestryUniversity,2011.
[9]日沙来提·吐尔地,艾斯卡尔·买买提,日孜汗·阿布地艾尼,等.一种从马鹿粪便中提取DNA的改进方法[J].安徽农业科学,2012,40(10):5739-5740,5754.
[10]Mahmut H,Ganzorig S,Onuma M.A preliminary study of the genetic diversity of Xinjiang Tarim red deer(Cervus elaphus yarkandensis)usingthemicrosatelliteDNAmethod[J].JpnJVetRes,2001,49:231-237.
[11]Bellemain E,Swenson J E,Tallmon D,et al.Estimating population size of elusive animals with DNA from hunter-collected feces:four methods for brown bears[J].Conservation Biology,2005,19(1):150-161.
Effects of Road on Genetic Differentiation of Tianshan Red Deer(Cervus elaphus songaricus)
Zhou Canlin1,2,Aysajan Tohti1,Mahmut Halik1,et al
(1.College ofLife Science and Technology,XinjiangUniversity,Urumqi 830046,China;2.Basic Medical College,XinjiangMedical University,Urumqi 830046,China)
To study the effects of highway on large mammals,Tianshan red deer fece samples from the both side of road S110,203,303 were collected.After the individual recognition by5 kinds ofmicrosatellite,the genetic diversity,genetic distance,heterozygositywere determined usingArlequin 3.11.The results showed that the five kinds ofmicrosatellites had good polymorphism,the highwayor road on habitat fragmentation could lead topopulation genetic change.When the road did not isolate the habitat,the populations along highways had no difference;when the habitat was completely isolated by highway,the heterozygosity and the number of genotypes decreased;The isolation effect of highway in use on the population was stronger than that of the road abandoned.
highway;Tianshan red deer;genetic differentiation;habitat fragmentation
S825.2
A
2095-3887(2014)06-0005-05
10.3969/j.issn.2095-3887.2014.06.001
2014-08-03
国家自然科学基金项目(31060152);新疆维吾尔自治区自然科学基金(2010211A02);新疆大学自治区重点学科-动物学科资助
周璨林(1980-),男,副教授,博士。
马合木提·哈力克(1959-),男,维吾尔族,教授,博士,博士生导师,主要从事哺乳动物的保护遗传学及其分子进化研究。
猜你喜欢
中国临床医学影像杂志(2022年5期)2022-07-26
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
南京师范大学学报(工程技术版)(2021年2期)2021-10-20
小学生(看图说画)(2020年6期)2020-06-10
建材发展导向(2019年10期)2019-08-24
东方企业家(2018年10期)2018-10-22
中南林业科技大学学报(2015年6期)2015-12-20
筑路机械与施工机械化(2014年10期)2014-03-01
筑路机械与施工机械化(2014年7期)2014-03-01
筑路机械与施工机械化(2014年2期)2014-03-01
