
厌恶情绪加工特点的事件相关电位研究*
2014-02-06 02:27:01张丹丹柳昀哲罗跃嘉
心理学报 2014年11期
金 熠 张丹丹 柳昀哲 罗跃嘉
(1北京师范大学 认知神经科学与学习国家重点实验室, 北京 100875)(2深圳大学 情绪与社会神经科学研究所, 深圳 518060)
1 引言
厌恶(disgust)是一种由不愉快的、反感的刺激引起的负性情绪体验(Rozin, Haidt, & McCauley,2000), 是人类基本情绪之一。厌恶原型来自口腔对食物的拒绝(Angyal, 1941; Rozin & Fallon, 1987)。由于引起厌恶情绪的对象(如排泄物、蟑螂等)通常具有污染性或传播疾病的可能, 因而厌恶的主观体验为反感并常伴随呕吐反应(Rozin & Fallon, 1987)及躲避行为(Woody & Teachman, 2000), 这些反应具有保护有机体免受疾病和污染的适应意义(Miller, 2004)。经典厌恶表情肌肉活动包括鼻子皱缩和嘴角朝下(Davey, 2004; Ekman, Levenson, &Friesen, 1983), 代表感觉器官对外界刺激输入的抑制(如皱缩的鼻子使得鼻子容积变小、气流速度减慢) (Susskind et al., 2008)。生理方面, 厌恶情绪下个体心率减慢(Ekman et al., 1983; Stark, Walter,Schienle, & Vaitl, 2005)。
脑损伤研究表明, 前部脑岛(Calder, Keane,Manes, Antoun, & Young, 2000)和基底节(Hayes,Stevenson, & Coltheart, 2009; Sprengelmeyer, Rausch,Eysel, & Przuntek, 1998; Suzuki, Hoshino, Shigemasu,& Kawamura, 2006)受损的病人不仅在识别厌恶表情上存在困难, 其表达厌恶情绪的能力也受到抑制,提示这两个脑区可能与厌恶加工有着最为密切的联系。对正常人的脑成像研究也发现观看厌恶图片时个体前部脑岛显著激活(Phillips et al., 1997;Wicker et al., 2003), 与脑损伤研究结果相辅相成。事件相关电位技术(event-related potential, ERP)则从时间角度探讨厌恶加工的神经机制。一些研究认为最早反应厌恶情绪效应的是中线上P2成分:无论是加工厌恶情绪图(Carretié, Ruiz-Padial, López-Martín, & Albert, 2011; Wheaton et al., 2013)还是加工厌恶表情(Sarlo & Munafò, 2010)时都能检测到P2波幅增大, 说明加工负性情绪刺激时皮层活动增强, 与传统情绪加工理论一致(Ito, Larsen, Smith,& Cacioppo, 1998; Smith, Cacioppo, Larsen, &Chartrand, 2003)。而近年来有些学者却认为厌恶拥有和恐惧等负性情绪不同的加工机制:厌恶刺激在情绪加工早期先引起皮层的抑制活动, 随后才增大皮层活动以进行精细加工; 他们的研究亦发现最早体现厌恶情绪效应的为视觉区P1成分(96 ms)(Krusemark & Li, 2011; 2013), 这一成分比同时期的其他研究结果暗示的时间更早。在本研究中, 我们也将使用事件相关电位技术检验被试在观看厌恶图片时的脑电活动变化。根据前人研究, 我们假设厌恶情绪效应产生较早, 大约在刺激后100 ms左右被检测到, 呈抑制趋势; 晚期精细加工情绪刺激时则能观察到脑电活动增强。
Halgren和Marinkovic (1995)在负性情绪加工模型中将情绪加工分为两阶段:第一阶段为“朝向”(orienting), 此阶段独立于意识进行; 第二阶段为“事件整合” (event integration), 需要意识参与, 为深度加工情绪刺激。后来Liddell等人用后掩蔽范式研究恐惧脸阈上/阈下加工特点时验证了该理论中描述的两个加工阶段的存在:在他们的研究中N2和早期P3在阈下加工情绪刺激时显著增强, 而N4和晚期P3则在阈上加工时增强(Liddell,Williams, Rathjen, Shevrin, & Gordon, 2004)。不仅如此, LeDoux等在动物研究基础上亦提出相似的双通道模型用以解释恐惧加工机制。他们认为杏仁核在恐惧情绪形成中具有重要作用; 以杏仁核为中心存在两条情绪加工通路:一条称为“低通路” (low road), 由下丘脑直接传递信息到杏仁核, 起快速、自动化加工情绪刺激的作用; 另一条称为“高通路”(high road), 存在于丘脑‒海马‒杏仁核网络中, 起细致评估情绪刺激并调节行为反应的作用(LeDoux,1998)。根据这一模型, 情绪刺激加工应该是自动化加工和控制加工的整合活动。我们研究中将考察厌恶加工是否存在上述两类加工进程, 并与LeDoux及Liddell等人的模型进行比较。
威胁类情绪刺激能自动捕获注意(Charash &McKay, 2002; Vogt, Lozo, Koster, & De Houwer,2011)、引起相应的自动化加工为当前普遍接受的研究结果。但情绪自动化加工是否必须需要消耗注意资源则成为近年来情绪研究领域颇受争议的话题。传统观点认为, 对负性情绪刺激的加工是自下而上、不需要消耗注意资源就能自动完成的(Vuilleumier, Armony, Driver, & Dolan, 2001)。而随后的研究结果则极大挑战了这一论点, 有些研究认为加工威胁类情绪刺激仍然需要消耗注意资源(Pessoa, 2005)。比如在核磁研究中, 当被试的注意资源被高认知负荷的任务占用时(同时呈现威胁刺激(恐惧脸)和目标刺激)并不能检测到杏仁核活动的显著增强(Pessoa, McKenna, Gutierrez, &Ungerleider, 2002)。又比如在事件相关电位研究中,当被试的注意活动被分配到外显任务时, 尽管同时呈现情绪脸, 其诱发的ERP与中性条件下并无显著差异(Eimer, Holmes, & McGlone, 2003; Holmes,Vuilleumier, & Eimer, 2003)。基于此, 我们假设自动化加工厌恶刺激需要消耗注意资源, 这一消耗可以从被试任务成绩变差中被观察到。
由于视觉搜索任务可以较好的反映个体注意资源分配情况(当情绪刺激和搜索任务同时呈现,如果个体任务表现变差, 说明加工情绪刺激消耗了注意资源; 如果任务表现没有变化, 则说明同时呈现的情绪刺激没有消耗注意资源), 因此被用作为本研究的实验任务(Krusemark & Li, 2011)。如果情绪刺激和任务时同时出现而任务表现因此受到干扰, 说明情绪刺激获得自动化加工, 并且消耗了注意资源。如果情绪刺激先于任务出现, 而随后的任务成绩受干扰, 说明任务开始时自动化加工情绪刺激尚未结束; 如果情绪刺激先于任务出现, 而任务成绩不受干扰, 则说明任务开始前自动化加工已完成, 此时加工情绪刺激需要自上而下控制进行。
2 研究方法
2.1 被试
25名北京地区大学生或研究生(13名女生, 平均年龄23.9岁)参与了此次研究。所有被试均为右利手, 视力正常或者矫正正常。实验结束后获得适当的报酬。
2.2 实验材料
刺激材料为100张情绪图片:50张厌恶图片,50张中性图片, 每张图片呈现4次。100张情绪图中29张来自国际情绪图片系统(IAPS) (Lang,Bradley, & Cuthbert, 2008), 17张来自中国情绪图片系统(CAPS) (白露, 马慧, 黄宇霞, 罗跃嘉, 2005),余下46张来自互联网。中性图片内容为书本、杯子、蘑菇等日常用品, 厌恶图片内容为尸体、蛆虫、排泄物等常见厌恶情绪引发物。另外招募22名被试参与对图片内容效价评定(0~9打分:0完全不恶心–9非常恶心):两类图片效价差异显著,F
(1,21) =70.23,p
< 0.001; 厌恶图片分数[M
(SD
): 5.43(2.20),下文同格式]显著高于中性图片[1.43(0.99)]。图片尺寸均为330×330象素。为保证图片情绪效度, 实验采用彩色图片作为刺激材料。视空间搜索任务图片采用Matlab软件编写制作。一共200张不同的条形布局图, 每张包含7个竖直条形和1个水平条形(搜索目标) (如图1), 条形均为紫色。每张条形布局图在实验中重复两次。目标条形(水平条形)位置在4个象限中随机分布(各以25%的概率出现)。

图1 实验流程示意图
2.3 实验设计
实验为2×4被试内设计。自变量一为图片情绪类型, 有中性/厌恶两个水平。自变量二为情绪图片单独呈现时间, 分为0/300/600/900 ms四个水平。0 ms表示情绪图和搜索任务同时出现, 考察对厌恶刺激的自动化加工。300 ms呈现条件表示情绪图先呈现300 ms再在情绪图片上叠加呈现搜索条形布局图, 此时被试在任务开始前已进行了一段时间的图片内容加工, 但未精细编码。在随后的视觉搜索任务中, 被试任务成绩如果受到情绪图效价影响,说明刺激后300 ms时自动化加工仍存在, 如果不受情绪图效价影响, 则说明自动化加工已消失。600/900 ms呈现条件与300 ms呈现条件一样, 被试在开始视觉搜索任务之前会先进行600/900 ms的情绪图加工, 一般认为500 ms以后的脑电成分主要反映自上而下加工进程, 与情绪刺激的评估有关。根据被试行为成绩可判断600 ms后的自主加工进程中是否含有自动化加工成分。
按照自变量二的4个水平将实验分为4个block, block顺序在被试内随机。每个block包含50张中性图和50张负性图, 顺序随机, 图片内容不重复。每个block结束时给出正确率和平均反应时作为反馈。Block之间允许被试自主控制休息时间。
2.4 实验流程
实验在安静的ERP实验室中进行。刺激材料用e-prime 1.0控制呈现, 记录被试行为数据(任务反应时、正确率)。脑电及眼电数据由64导NeuroScan 4.5记录。实验中, 被试距离屏幕约为1米, 实验材料均呈现于屏幕正中。实验流程如图1所示:注视点后呈现情绪图片(0/300/600/900 ms, 视角:6.2°×6.2°), 随后图片上叠加搜索条形布局图(500 ms), 被试需要尽快对水平条形(目标)所在的位置进行按键反应:左上按“U”键, 右上按“I”键, 左下按“J”键, 右下按“K”键, 均用右手按键。按键在键盘上的布局与其所代表的象限位置相对应。条形布局呈现500 ms后消失, 800~1200 ms试次间隔后进入下一试次。
实验总时长20~25 min。正式实验前被试至少进行20试次练习(额外选取10张中性图), 以保证其完全熟悉任务要求和反应键。
2.5 脑电数据预处理和分析
EEG使用64导NeuroScan 4.5系统在250 Hz采样率下记录, 在线滤波参数为0.05~100 Hz。用四枚电极记录眼电:两枚贴于左右外眼角距离眼睛1 cm处用以记录水平眼电, 两枚贴于左眼上下距离眼睛1cm处用以记录垂直眼电。EEG信号记录时以左侧乳突作为参考, 离线后转双耳连线参考。离线滤波参数为0.05~30 Hz。选取图片呈现前200 ms作为ERP数据分段的起点, 单试次数据分析终止时间为“呈现条件+1200 ms” (比如, 图片呈现条件为300 ms, 则窗口结束时间为图片呈现后1500 ms)。排除EEG电压超过±100 μV (相对于200 ms的基线)的试次, 最后参与分析的试次总计8546个。眼电伪迹采用NeuroScan 4.5分析软件去除。
计算300/600/900 ms呈现条件下厌恶/中性图片平均波幅:从图片呈现后50 ms到图片后300 ms,每50 ms为一个时间区间, 共5个时间区间。计算600/900 ms呈现条件下的厌恶/中性图片平均波幅:从图片呈现后300 ms到图片后600 ms, 每50 ms为一个时间区间, 共6个时间区间。计算900 ms呈现条件下的厌恶/中性图片平均波幅:从图片呈现后600 ms后到图片后900 ms, 每50 ms为一个时间区间, 共6个时间区间。对上述17个时间区间逐一进行情绪效应检测。由于情绪刺激呈现早期即引起初级视觉区活动(Krusemark & Li, 2011), 故选用OZ作为第一个参与分析的电极点; 并且由于反映晚期精细加工的LPP在后部中央位置最为显著(Hajcak, Moser, & Simons, 2006), 故选用CPZ作为第二个参与分析的电极点。
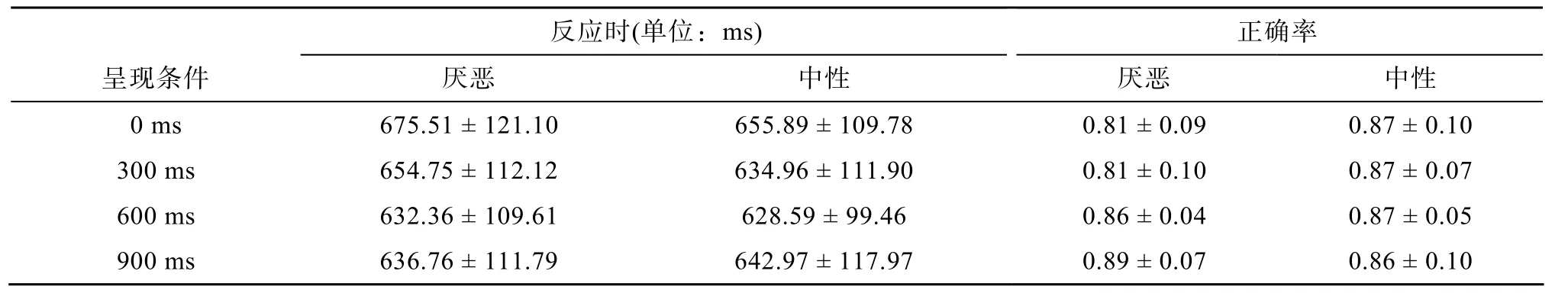
表1 反应时和正确率描述统计量

图2 反应时情绪类型和呈现条件交互作用

图3 正确率情绪类型和呈现条件交互作用
2.6 行为数据统计分析
统计分析使用SPSS 17.0完成。描述性数据以平均数 ± 标准差(M
±SD
)的方式呈现, 显著性水平设定为0.05。对行为结果进行用2×4重复测量方差分析, 考察因素为情绪类型(厌恶/中性)和情绪图片呈现条件(0/300/600/900 ms), 均为被试内因素;因变量为视觉搜索任务反应时(RT)及正确率(ACC)。使用Greenhouse-Geisser矫正ANOVA分析结果。显著的交互作用采用简单效应模型继续分析。3 结果
3.1 行为结果
反应时和正确率描述统计量如表1所示。
反应时两因素ANOVA分析结果:情绪类型主效应显著,F
(1,24) = 10.62,p
= 0.003; 呈现条件主效应不显著,F
(3,72) = 2.33,p
= 0.081; 情绪类型×呈现条件交互作用显著,F
(3,72) = 4.141,p
= 0.026(如图2)。简单效应分析结果:呈现条件0/300 ms时, 情绪类型简单效应显著(0 ms:F
(1,24) = 7.58,p
= 0.011; 300 ms:F
(1,24) = 16.36,p
< 0.001), 厌恶条件下任务反应时显著高于中性条件; 呈现条件600/900 ms时, 情绪类型简单效应不显著(600 ms:F
(1,24) = 0.45,p
= 0.508; 900 ms:F
(1,24) = 0.88,p
= 0.358); 厌恶条件下, 呈现条件简单效应显著(F
(3,72) = 3.52,p
= 0.019):0 ms呈现条件下被试反应时显著高于600 ms (p
= 0.003)/900 ms (p
= 0.038)下的反应时, 其他两两比较均不显著; 中性条件下,呈现条件简单效应不显著,F
(3,72) = 1.38,p
=0.254。正确率两因素ANOVA分析结果:情绪类型主效应显著,F
(1,24) = 18.02,p
< 0.001; 呈现条件主效应显著,F
(3,72) = 3.397,p
= 0.043; 情绪类型×呈现条件交互作用显著,F
(3,72) = 11.255,p
< 0.001(如图3)。简单效应检验发现呈现条件为0 ms[F
(1,24)= 22.15,p
< 0.001]、300 ms [F
(1,24) = 21.21,p
<0.001]、900 ms [F
(1,24) = 6.05,p
= 0.022]时, 情绪类型简单效应显著:0/300 ms下, 中性条件搜索任务正确率显著高于厌恶条件的正确率; 900 ms下,厌恶条件的正确率显著高于中性条件的正确率。厌恶情绪下, 呈现条件简单效应显著[F
(1,24) = 14.74,p
< 0.001]:呈现条件为0/300 ms时的正确率显著低于600/900 ms时的正确率(p
s < 0.006); 中性情绪下, 呈现条件简单效应不显著,F
(3,72) = 0.28,p
=0.840。
图4 不同呈现条件下OZ/CPZ点ERP总平均波形
3.2 脑电结果
不同呈现条件下OZ/CPZ点总平均ERP波形如图4。
厌恶情绪加工进程:厌恶/中性图呈现后50~900 ms每50 ms为一个时间区间, 计算各区间各情绪条件下平均波幅并检验情绪效应。平均波幅和效应检验结果呈现如表2。
4 讨论
本研究的第一个目的是利用事件相关电位技术的时间精度优势探究厌恶图片加工特点。我们发现, 视觉区脑电活动在100 ms产生情绪效应, 反映注意机制对威胁类信息的快速警觉, 对有机体生存有重要意义(Lang, Bradley, & Cuthbert, 1997)。视觉区早期情绪效应持续到150 ms左右, 呈显著的正向波形, 而厌恶对应的波形相对于中性基线呈抑制趋势。和OZ一样, CPZ上情绪效应最早也出现于100 ms, 持续至200 ms, 为负波形式, 并同样呈现负性情绪抑制趋势。OZ和CPZ上观察到的结果说明在情绪加工早期厌恶诱发了皮层抑制活动, 个体通过抑制感觉信息输入来拒绝与有害刺激接触。厌恶的这一皮层加工特点与其生理反应和行为表现相对应, 支持了厌恶“感觉拒绝” (sensory rejection)的适应功能, 从而帮助有机体同潜在威胁保持距离(Rozin & Fallon, 1987; Susskind et al., 2008), 众多围绕厌恶进行的生理、行为、脑机制的研究亦论证了该观点(Ekman et al., 1983; Krusemark & Li, 2011;2013; Susskind et al., 2008)。CPZ在250 ms开始第二次出现情绪效应, 该效应在随后的情绪刺激呈现过程中持续存在。观察这一阶段的波形(如图4), 主要包含一个负成分N2和随后的晚期正成分LPP。OZ第二次情绪效应从350 ms开始, 持续到550 ms结束, 主要也是晚期正成分LPP。LPP反应了对刺激自上而下的有意识注意(Hajcak et al., 2006), 并且被认为与刺激显著性及对刺激意义的精细加工有关(Olofsson, Nordin, Sequeira, & Polich, 2008;Schupp, Flaisch, Stockburger, & Junghöfer, 2006)。一般认为LPP在顶叶中后部达到最大, 本研究中对比OZ和CPZ上同时出现的LPP情绪效应, 也得到相似的结论。LPP增大反映对情绪内容注意增强(Schupp, Junghofer, Weike, & Hamm, 2003), 说明厌恶图片包含的负性内容引起被试投入更大的注意资源进行精细加工(Ito et al., 1998; Smith et al.,2003)。由此, 我们的研究结果支持了厌恶加工具有独特神经机制的观点, 其独特性表现为皮层活动的先抑制后增强, 而这一机制与其进化而来的适应性功能紧密相关。
本研究的第二个目的是探究情绪自动化加工和控制加工的两阶段模型是否同样适用于描述厌恶加工进程, 并检验情绪自动化加工阶段中注意资源的消耗情况。我们发现, 负性情绪图阈下加工(呈现条件0 ms)时被试的任务表现最差, 体现为反应时最长和正确率最低, 而阈下加工导致任务变差仅出现在厌恶条件下, 中性条件中被试表现未受呈现时间影响。由于0 ms呈现条件下情绪刺激作为干扰刺激与视觉搜索任务同时出现, 因此此时对情绪刺激的加工属于自动化加工, 而我们数据则表明这一加工活动消耗了注意资源, 极大支持了Pessoa等人的观点(Pessoa, 2005)。300 ms呈现条件下, 被试的任务成绩仍有情绪效应, 说明任务开始时对厌恶的自动化加工仍未结束。对比600 ms呈现条件下被试行为成绩不受情绪刺激干扰的结果, 我们认为自动化加工结束于图片后300~600 ms之间。Santos等人(2008)检测对厌恶表情的加工特点时发现, 480 ms以后的脑电成分只在被试注意脸时才有情绪效应, 如果被试注意的是房子, 那么与之同时呈现的情绪脸(干扰刺激)在480 ms以后并不能得到自动加工(Santos, Iglesias, Olivares, & Young, 2008)。我们的研究结果某种程度上和Santos等人的研究结果相吻合。另一方面, 我们注意到在加工了300 ms的情绪刺激后, 被试行为成绩虽仍受影响, 但相比0 ms呈现条件下有提高, 说明刺激后300 ms时不但存在自动化加工, 自上而下的控制加工也并行存在。以往研究在探讨情绪加工两阶段模型时并未强调两类加工并存的情况, 而我们的数据则支持了这一点。

表2 各时间区间内平均波幅(单位:μV)和情绪效应显著性检验结果
600/900 ms呈现条件考察了对情绪图的精细编码。一般认为事件相关电位的晚期正成分(LPP)标志自上而下控制加工, 与图片的唤醒度有关, 反映对情绪刺激的复杂评估和精细编码(Olofsson et al., 2008)。我们的研究结果表明, 在精细编码阶段,厌恶引起ERP波幅显著大于中性条件的波幅, 提示此阶段对负性情绪刺激的编码和加工更为复杂。然而又因为对应的反应时没有呈现显著情绪效应,说明在300~600 ms之间的某一时刻后对情绪图的加工已经变为完全的自上而下控制加工, 在这一时刻后开始视搜索任务被试可以迅速转移注意力到任务上而不受先前加工的情绪刺激的干扰。有趣的是, 在精细加工了900 ms的负性图片之后, 被试任务正确率甚至提高了, 这可能是因为负性情绪刺激通常能引起被试更高的唤醒度(Britton, Taylor,Sudheimer, & Liberzon, 2006; Lang et al., 1997;Schupp et al., 2004), 而唤醒度提高后被试能够集中更多注意资源, 从而有利于任务的完成。当然这种情况出现前提是负性情绪刺激已经经过足够长时间的精细编码、结束了自动化加工而进入完全的控制加工阶段。无论是LeDoux等人的双通道加工模型(LeDoux, 1998)还是Liddell等人的两阶段加工模型(Liddell et al., 2004)都是建立在对恐惧情绪研究基础上的, 本研究则证明了上述两阶段模型对厌恶加工同样适用。不仅如此, 我们的研究还确认了过渡阶段的存在:自动化加工消失转变为完全控制加工之前存在两种加工共同作用的阶段, 某种程度上是对前人理论的补充。
总结来说本研究发现了厌恶情绪加工的独特机制:先抑制感觉刺激输入, 随后增大皮层活动深度编码情绪刺激。负性情绪加工包含两阶段, 首先是自动化加工, 此时需要消耗注意资源; 300~600 ms之间某个时间点是自动化加工的结束时间, 此后为完全的自上而下控制加工。自动化加工结束之前存在两类加工并存的情况。由于时间仓促, 我们研究中亦存在不足之处, 有些学者认为个体焦虑特质会影响其对负性刺激的反应(Li, Zinbarg, &Paller, 2007), 而我们认为参与实验的被试均为正常情绪状态故未考察其特质焦虑水平可能对研究结果的影响; 另外我们选用实验材料时根据实验目的主要控制了图片的效价, 没有收集数据评估图片复杂度、亮度等物理属性可能对实验结果产生的影响, 这些是本研究的疏漏, 将一并在今后实验中改进。
Angyal, A. (1941). Disgust and related aversions.The Journalof Abnormal and Social Psychology, 36
(3), 393–412.Britton, J. C., Taylor, S. F., Sudheimer, K. D., & Liberzon, I.(2006). Facial expressions and complex iaps pictures:Common and differential networks.NeuroImage, 31
(2),906–919.Bai, L., Ma, H., Huang, Y. X., & Luo, Y. J. (2005). The development of native Chinese affective picture system—a pretest in 46 college students.Chinese Mental Health Journal, 19
(11), 719–722.[白露, 马慧, 黄宇霞, 罗跃嘉. (2005). 中国情绪图片系统的编制--在46名中国大学生中的试用.中国心理卫生杂志, 19
(11), 719-722.]Calder, A. J., Keane, J., Manes, F., Antoun, N., & Young, A. W.(2000). Impaired recognition and experience of disgust following brain injury.Nature Neuroscience, 3
(11),1077–1078.Carretié, L., Ruiz-Padial, E., López-Martín, S., & Albert, J.(2011). Decomposing unpleasantness: Differential exogenous attention to disgusting and fearful stimuli.Biological Psychology, 86
(3), 247–253.Charash, M., & McKay, D. (2002). Attention bias for disgust.Journal of Anxiety Disorders, 16
(5), 529–541.Davey, G. C. L. (2004). Disgust. In V. S. Ramachandran (Ed.),Encyclopedia of human behavior
(pp. 702–709). San Diego,CA: SanDiego Press.Eimer, M., Holmes, A., & McGlone, F. P. (2003). The role of spatial attention in the processing of facial expression: An erp study of rapid brain responses to six basic emotions.Cognitive, Affective & Behavioral Neuroscience, 3
(2),97–110.Ekman, P., Levenson, R. W., & Friesen, W. V. (1983).Autonomic nervous system activity distinguishes among emotions.Science, 221
(4616), 1208–1210.Hajcak, G., Moser, J. S., & Simons, R. F. (2006). Attending to affect: Appraisal strategies modulate the electrocortical response to arousing pictures.Emotion, 6
(3), 517–522.Halgren, E., & Marinkovic, K. (1995). Neurophysiological networks integrating human emotions. In M. S. Gazzaniga(Ed.), The cognitive neurosciences (pp. 1137–1151).Cambridge, MA: MIT Press.
Hayes, C. J., Stevenson, R. J., & Coltheart, M. (2009).Production of spontaneous and posed facial expressions in patients with huntington's disease: Impaired communication of disgust.Cognition & Emotion, 23
(1),118–134.Holmes, A., Vuilleumier, P., & Eimer, M. (2003). The processing of emotional facial expression is gated by spatial attention: Evidence from event-related brain potentials.Cognitive Brain Research, 16
(2), 174–184.Ito, T. A., Larsen, J. T., Smith, N. K., & Cacioppo, J. T. (1998).Negative information weighs more heavily on the brain:The negativity bias in evaluative categorizations.Journal of Personality and Social Psychology, 75
(4), 887–900.Krusemark, E. A., & Li, W. (2011). Do all threats work the same way? Divergent effects of fear and disgust on sensory perception and attention.The Journal of Neuroscience,31
(9), 3429 –3434Krusemark, E. A., & Li, W. (2013). From early sensory specialization to later perceptual generalization: Dynamic temporal progression in perceiving individual threats.The Journal of Neuroscience, 33
(2), 587–594.Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1997).Motivated attention: Affect, activation and action. In P. J.Lang, R. F. Simons, & M. T. Balaban (Ed.),Attention and orienting: Sensory and motivational processes
(pp. 97–136).Hillsdale, NJ: Erlbaum.Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (2008).International affective picture system (iaps): Affective ratings of pictures and instruction manual
.LeDoux, J. E. (1998).The emotional brain: The mysterious underpinnings of emotional life
. New York: Touchstone.Li, W., Zinbarg, R. E., & Paller, K. A. (2007). Trait anxiety modulates supraliminal and subliminal threat: Brain potential evidence for early and late processing influences.Cognitive,Affective & Behavioral Neuroscience, 7
(1), 25–36.Liddell, B. J., Williams, L. M., Rathjen, J., Shevrin, H., & Gordon,E. (2004). A temporal dissociation of subliminal versus supraliminal fear perception: An event-related potential study.Journal of Cogntive Neurosciece, 16
(3), 479–486.Miller, S. B. (2004).Disgust: The gatekeeper emotion.
Mahwah, NJ: Analytic Press.Olofsson, J. K., Nordin, S., Sequeira, H., & Polich, J. (2008).Affective picture processing: An integrative review of erp findings.Biological Psychology, 77
(3), 247–265.Pessoa, L. (2005). To what extent are emotional visual stimuli processed without attention and awareness?Current Opinion in Neurobiology, 15
(2), 188–196.Pessoa, L., McKenna, M., Gutierrez, E., & Ungerleider, L. G.(2002). Neural processing of emotional faces requires attention.Proceedings of the National Academy of Science of the United States of America, 99
(17), 11458–11463.Phillips, M. L., Young, A. W., Senior, C., Brammer, M.,Andrew, C., Calder, A. J., . . . David, A. S. (1997). A specific neural substrate for perceiving facial expressions of disgust.Nature, 389
(6650), 495–498.Rozin, P., & Fallon, A. E. (1987). A perspective on disgust.Psychological Review, 94
(1), 23–41.Rozin, P., Haidt, J., & McCauley, C. R. (2000). Disgust. In J.M. Haviland-Jones (Ed.),Handbook of emotions
(pp.637–653). New York: Guildford Press.Santos, I. M., Iglesias, J., Olivares, E. I., & Young, A. W.(2008). Differential effects of object-based attention on evoked potentials to fearful and disgusted faces.Neuropsychologia, 46
(5), 1468–1479.Sarlo, M., & Munafò, M. (2010). When faces signal danger:Event-related potentials to emotional facial expressions in animal phobics.Neuropsychobiology, 62
(4), 235–244.Schupp, H. T., Cuthbert, B. N., Bradley, M. M., Hillman, C. H.,Hamm, A. O., & Lang, P. J. (2004). Brain processes in emotional perception: Motivated attention.Cognition &Emotion, 18
(5), 593–611.Schupp, H. T., Flaisch, T., Stockburger, J., & Junghöfer, M.(2006). Emotion and attention: Event-related brain potential studies.Progress in Brain Research, 156,
31–51.Schupp, H. T., Junghofer, M., Weike, A. I., & Hamm, A. O.(2003). Emotional facilitation of sensory processing in the visual cortex.Psychological Science, 14
(1), 7–13.Smith, N. K., Cacioppo, J. T., Larsen, J. T., & Chartrand, T. L.(2003). May i have your attention, please: Electrocortical responses to positive and negative stimuli.Neuropsychologia, 41
(2), 171–183.Sprengelmeyer, R., Rausch, M., Eysel, U. T., & Przuntek, H.(1998). Neural structures associated with recognition of facial expressions of basic emotions.Proceedings of the Royal Society of London. Series B: Biological Sciences,265
(1409), 1927–1931.Stark, R., Walter, B., Schienle, A., & Vaitl, D. (2005).Psychophysiological correlates of disgust and disgust sensitivity.Journal of Psychophysiology, 19
(1), 50–60.Susskind, J. M., Lee, D. H., Cusi, A., Feiman, R., Grabski, W.,& Anderson, A. K. (2008). Expressing fear enhances sensory acquisition.Nature Neuroscience, 11,
843–850.Suzuki, A., Hoshino, T., Shigemasu, K., & Kawamura, M.(2006). Disgust–specific impairment of facial expression recognition in parkinson's disease.Brain, 129
(3), 707–717.Vogt, J., Lozo, L., Koster, E. H., & De Houwer, J. (2011). On the role of goal relevance in emotional attention: Disgust evokes early attention to cleanliness.Cognition & Emotion,25
(3), 466–477.Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J.(2001). Effects of attention and emotion on face processing in the human brain: An event-related fmri study.Neuron,30
(3), 829–841.Wheaton, M. G., Holman, A., Rabinak, C. A., MacNamara, A.,Proudfit, G. H., & Phan, K. L. (2013). Danger and disease:Electrocortical responses to threat- and disgust-eliciting images.International Journal of Psychophysiology, 90
(2),235–239.Wicker, B., Keysers, C., Plailly, J., Royet, J.-P., Gallese, V., &Rizzolatti, G. (2003). Both of us disgusted in my insula:The common neural basis of seeing and feeling disgust.Neuron, 40
(3), 655–664.Woody, S. R., & Teachman, B. A. (2000). Intersection of disgust and fear: Normative and pathological views.Clinical Psychology: Science and Practice, 7
(3), 291–311.猜你喜欢
中华养生保健(2020年7期)2020-11-16 01:14:26
初中生学习指导·中考版(2020年2期)2020-09-10 07:22:44
英语学习(上半月)(2019年9期)2019-10-10 02:17:38
中国医药指南(2017年3期)2017-11-13 02:58:30
家教世界·创新阅读(2016年11期)2016-12-27 18:49:15
天津护理(2016年3期)2016-12-01 05:40:01
故事会(2016年15期)2016-08-23 13:48:41
米娜·女性大世界(2016年8期)2016-08-17 17:01:00
工业设计(2016年11期)2016-04-16 02:44:40
集美大学学报(教育科学版)(2015年1期)2015-02-28 01:11:46
