
人际合作与冲突影响博弈决策的结果评价*
2014-02-06 02:27:10白丽英王益文
心理学报 2014年11期
白丽英 袁 博 张 蔚 张 振 兰 姣 王益文
(1福州大学人文学院应用心理学系, 福州 350116) (2宁波大学心理学系暨研究所, 宁波 315211)
(3天津师范大学心理与行为研究院, 天津 300074)
1 引言
人类社会和其他灵长类动物界都存在着合作与冲突行为。在人类社会中, 合作与冲突行为是儿童社会化的一个主要方面, 也是人际互动的两种主要形式。人际合作表现为两个或两个以上的人或群体为达到共同目的, 自觉或不自觉地在行动上相互配合的一种互动方式; 人际冲突则指人与人、群体与群体对于一个共同目标的争夺(Decety, Jackson,Sommerville, Chaminade, & Meltzoff, 2004)。在实验室研究中, 研究者借助于表现人际互动行为的社会博弈任务, 研究和分析人类的合作与冲突行为。这些社会博弈任务对现实中的社会困境进行了简化,并且提供了选择策略的数学化形式, 可以让研究者在实验室情境中为类似于“合作”或者“冲突”这样的概念进行操作性的定义(Camerer, 2003; Rilling et al., 2002)。囚徒困境(Prisoner's Dilemmas Game)是研究人类合作行为的一个经典博弈任务, 在这个任务中, 两个玩家分别选择合作或者背叛, 每个玩家根据两个人的共同选择获得一定数目的收益。对每个玩家而言, 无论对方做出何种选择, 做出背叛选择对自己总是最优选择策略。但当两个玩家都选择背叛, 两个人的总体收益最低; 而两个人都选择合作, 则总体的收益最大(Rilling et al., 2002; Wong &Hong, 2005)。目前, 囚徒困境任务被心理学家和社会学家广泛应用于研究人类的合作行为。与囚徒困境任务类似, Chicken Game任务(有译为:斗鸡博弈任务)也是一个研究合作与冲突行为的博弈任务。Chicken Game创设了一个更为真实的社会情境,在这个任务中, 两个博弈者可以独立地选择和解或者进攻两种策略, 两个人都选择和解双方都会获得一定数目的收益; 如果其中一方选择和解, 另一方选择进攻, 选择和解的一方将损失一定数目的收益,选择进攻的一方将获得一笔较大的收益; 如果两个人都选择进攻, 双方都会损失一笔更大数目的收益。与囚徒困境不同, Chicken Game中没有严格的优势策略, 选择进攻的收益并不一定比选择和解的收益大(Rapoport & Chammah, 1966)。Chicken Game的收益与人类合作与冲突行为所导致的结果非常类似(详见方法部分)。
随着脑成像技术的发展, 研究者已不满足仅仅从表层的行为结果来探讨人际合作与冲突行为。借助于功能性磁共振成像技术(functional-Magnetic Resonance Imaging, fMRI)和事件相关电位技术(event-related potentials, ERPs), 研究者将视点转向了人际合作和冲突行为背后的认知神经机制。Rilling等人(2002)采用fMRI记录了被试与人类同伴进行囚徒困境任务(Iterated Prisoner’s Dilemma Game)时的大脑激活情况, 结果发现, 个体选择相互合作时所激活的脑区主要包括:伏隔核(nucleus accumbens)、腹内侧额叶/眶额皮层(ventromedial frontal/orbitofrontal cortex)以及前扣带回(anterior cingulate cortex)。上述脑区都与奖励加工有关, 研究者认为这些脑区的激活促使被试抵制自私的诱惑, 进而选择互惠利他的合作行为。Fukui等人(2006)采用fMRI技术探讨了Chicken Game任务中与人类同伴进行社会博弈是否会激活心理理论的相关脑区, 并试图阐明心理理论脑区在博弈决策中的作用。研究设置了被试与人类同伴以及与电脑博弈两种实验条件。结果表明, 相对于和电脑进行博弈, 与人类同伴博弈时, 旁扣带回前部(anterior paracingulate cortex)以及后侧颞上沟(posterior superior temporal sulcus)有显著激活。并且研究发现, 当与人类同伴博弈时, 旁扣带回前部在选择冲突时激活更强。这些结果表明后侧颞上沟与旁扣带回前部在心智化过程中有不同的作用。后侧颞上沟可能参与心智化的一般功能, 而旁扣带回前部可能还参与社会风险决策过程。
除了采用fMRI技术探讨个体进行博弈决策选择阶段的脑区激活, 也有研究者采用ERPs技术考察决策之后的结果评价过程。结果评价是决策链中的一个重要环节, 合理的决策需要我们对先前的结果进行编码, 进而做出更好的抉择(Platt, 2002)。人类大脑已经发展出某些机制对结果反馈的各种属性进行快速评价, 已有研究表明, 结果评价涉及到对结果的效价(如输赢)、金额大小以及其他与结果反馈有关的信息加工过程(Leng & Zhou, 2010;Yeung, Holroyd, & Cohen, 2005; Yeung & Sanfey,2004)。研究者采用ERPs技术发现了一个特定的与结果评价有关的ERP成分, 被称为反馈相关负波(feedback-related negativity, FRN)或者内侧额叶负波(medial frontal negativity, MFN)。该成分大约在反馈呈现后250~300 ms内达到峰值, 其源定位在前扣带回(anterior cingulate cortex, ACC)附近, 输钱会比赢钱引发更大的FRN波幅(Gehring & Willoughby,2002; Masaki, Takeuchi, Gehring, Takasawa, &Yamazaki, 2006; Yu, Luo, Ye, & Zhou, 2007)。Holroyd, Hajcak和Larsen (2006)认为FRN反映了个体对当前所得到的结果是否符合预期的一种评价。另一些研究者则认为FRN反映的是对反馈刺激情绪动机意义的评价过程(Gehring & Willoughby, 2002;Masaki et al., 2006; Yu et al., 2007)。最近也有研究者提出, FRN可能反映了大脑的冲突监控过程, 反馈刺激引起的冲突越大, 其诱发的FRN波幅越大。FRN脑内源定位于ACC, 而ACC正是负责冲突监控的脑区(Gehring & Fencsik, 2001; Jia et al., 2007;Veen & Carter, 2002)。与结果评价有关的另一个ERP成分是P300, 其波峰出现在反馈后200~600 ms之间, 研究者认为P300主要是对结果的金额大小的加工(Sato et al., 2005; Yeung & Sanfey, 2004),但近期的研究也发现P300对结果的输赢也敏感(Hajcak, Holroyd, Moser, & Simons, 2005; Wu &Zhou, 2009; Yeung et al., 2005; Zhou, Yu, & Zhou,2010)。Yeung和Sanfey (2004)认为P300可能代表的是一种高水平的情绪动机的评价, 而不是结果效价的直接评价。
近期, 研究者采用FRN和P300成分作为电生理指标, 考察了各种社会情境中的结果评价过程。Hewig等人(2011)采用最后通牒任务研究了社会博弈中公平加工有关的心理生理学机制。其研究结果表明, 相对于公平提议, 不公平提议诱发了更大的FRN, 并且FRN的波幅可以预测被试对不公平提议的拒绝率。这表明, 在最后通牒任务中FRN与理解他人行为公平与否有关, FRN可以作为公平感觉察的一个指标。Long, Jiang和Zhou (2012)研究发现,信任感可以影响结果评价后的FRN和P300波幅, 相对于信任情境, 在不信任情境下, FRN效应会显著减弱。也有研究探讨了责任分散对结果评价的影响, 结果表明, 相对于低责任情境, 在高责任情境下结果反馈会诱发更大的FRN和P300波幅(Li, Han, Lei,Holroyd, & Li, 2011; Li et al., 2010)。这些研究均表明,FRN或者P300可以作为区分不同社会情境的脑电指标。那么, 人际合作与冲突情境中的结果评价过程,如何体现在FRN和P300波幅上?
现实生活中, 很多决策都是发生在复杂的社会情境中, 决策双方的选择是相互影响的。在一个可以产生相互合作或者冲突的社会博弈情境中, 个体如何评价不同博弈策略所导致的结果, 这些评价过程如何影响随后的决策行为; 博弈后的合作与冲突结果评价的时间进程反映在FRN和P300上有何差异?目前尚无研究深入地探讨这一问题。为此, 本研究试图采用ERPs技术和Chicken Game博弈任务探究合作与冲突社会互动情境中的结果评价过程。研究记录了被试与人类同伴进行Chicken Game博弈时的脑电活动, 将结果评价有关的FRN和P300作为电生理指标, 考察被试评价合作或者冲突结果时的心理加工过程。在Chicken Game任务中, 相对于选择冲突, 选择合作时个体会更加期待对方也选择合作; 因此, 选择合作可能会增加个体对博弈结果的预期。根据Holroyd等人(2006)的研究, FRN反映预期与实际结果之间的差异。本研究假设, 选择合作后输钱与赢钱引发的FRN波幅差异更大, 即合作情境下的FRN差异波(dFRN)会大于冲突情境下的FRN差异波(dFRN)。此外, 根据以往的研究,P300是反映注意分配或者高水平动机/情感以及社会性信息评价的成分(Leng & Zhou, 2010; Ma et al.,2011; Nieuwenhuis, Aston-Jones, & Cohen, 2005),而合作共赢的结果除了具有本身的物质奖励还有社会奖励的意义, 相互合作后的赢钱比单纯的赢钱更有意义。因此, 除了预期赢钱会比输钱引发更大的P300, 本研究还预期, 选择合作后的赢钱与输钱引发的P300差异会更大。
2 方法
2.1 方法
22名在校大学生(11男11女, 年龄在18~25岁之间, 平均年龄22.7 ± 2.41)参加了本次实验。2名实验助手充当与被试进行博弈任务的对手, 男女各1名, 年龄分别为24岁和26岁。实验中, 被试和对手之间性别匹配。所有被试都身心健康, 右利手,无神经系统疾病, 视力正常或矫正视力正常。实验前签署知情同意书, 实验后按照被试在实验任务的表现给予一定报酬。
2.2 任务和程序
本实验采用Chicken Game作为实验任务。Chicken Game类似于囚徒困境, 是研究人类合作与冲突行为的一种博弈任务。在这个任务中, 两个参与者都可以选择“和解” (Reconciliation)或者“进攻” (Aggression)这两种策略, 每个参与者得到的报酬由双方的选择共同决定。Chicken Game有四种可能的结果:参与者A和参与者B都选择和解(简称RR); 参与者A选择和解而参与者B选择进攻(简称RA); 参与者A选择进攻而参与者B选择和解(简称AR); 两个参与者都选择进攻(简称AA)。四种结果的收益满足以下条件:AR > RR > RA > AA, 并且AR + AA = RA + RR = 0 (Fukui et al., 2006)。Chicken Game强调的是, 如何在博弈中采用妥协的方式取得利益。如果双方都换位思考, 他们就会尽量选择和解的策略以保证两个人都可以获得一定的收益。但如果只从自己立场出发考虑问题, 就会更多地选择进攻策略, 为自己争取更多利益。Chicken Game可以给博弈者呈现一个比较真实的社会情境, 在这个情境中可以产生相互合作或者相互冲突的社会行为。囚徒困境 (Prisoner’s dilemma Game)任务中虽然也可以产生相互合作或者背叛的社会情景, 但囚徒困境任务中存在明显的优势策略, 即无论对方选择何种策略, 选择背叛的收益总是大于选择合作的收益。Chicken Game并不存在上述的优势策略,因此Chicken Game更类似于人类生活中经常遇到的社会问题。并且由于该任务没有明显的优势策略,更适合于ERP实验范式(ERP的平均叠加需要各种实验条件下的trials达到一定的数目)。
被试(A)来到实验室后, 先向他/她介绍另一名参与实验的被试B (实际为实验助手), 然后采集他们的照片, 作为实验的提示线索。告诉被试他/她将和参与者B (实验助手)通过局域网进行一个博弈任务, 并且依据他们在任务中的表现得到奖励。在调试好EEG电极之后, 被试坐在脑电实验室的房间,参与者B在邻近的另一房间。为了增加任务的真实性, 在实验开始前会带被试去另一个房间观看, 并且让被试看到参与者B就坐在这个房间的电脑前准备与他/她进行博弈任务。正式实验开始时告知被试在任务开始他们有20元的本钱, 这笔钱将会根据他们在随后任务的结果增加或减少, 每一局的博弈结果都和他们最终的报酬有关。被试可以在每一局的博弈中做出对自己最有利的选择。
每个trial开始首先呈现参与者B的照片500 ms, 接着呈现一个由不同选择策略导致不同结果的收益矩阵(水平视角: 6.9°; 垂直视角: 4.6°)。被试在看到这个收益矩阵后2000 ms内做出和解或者进攻的选择, 键盘上‘1’和‘3’分别代表和解和进攻策略, 按键在被试间平衡。要求被试在认真考虑后按键进行选择。在被试做出按键选择之后, 他/她所选择的策略会用红色标出, 并呈现1000 ms。最后呈现被试和搭档的共同选择所导致的结果1500 ms(水平视角: 4.9°; 垂直视角: 3.5°)。为了使结果反馈简单明了, 在呈现的反馈中, 只有被试和搭档的选择所引起的结果的单元被保留, 其他三个单元都被隐去。括弧外面用红色标出的钱数是被试所得到的收益, 括弧里面的钱数是参与者B的收益。被试得到对方“和解”或“进攻” (A或R)的反馈结果按50%的概率伪随机呈现。即当被试选择和解时, 50%的trials给被试呈现对方选择和解(即双方都选择和解,每人均赢得10元, RR)或者50%的trials给被试呈现对方选择进攻(即被试选择和解对方选择进攻, 被试输10元对方赢30元, RA)的结果; 同样当被试选择进攻时, 50%的trials给被试呈现对方选择和解(即被试选择进攻对方选择和解, 被试赢30元对方输10元, AR)或者对方选择进攻(即双方都选择和解,每人均输30元, AA)的结果。结果反馈出现在收益矩阵中的位置在被试间进行平衡。实验刺激之间的间隔以及trials之间间隔均为600~800 ms随机。被试先和搭档进行15个trial的练习实验, 在确保被试真正理解了实验程序之后开始正式实验。正式实验分为2个block, 每个block包括100 trials, 整个实验持续大约30 min。所有被试均表示相信实验中对方的选择是真人做出的。
2.3 脑电记录与分析
采用NeuroScan ERP记录与分析系统(NeuroScan Inc., USA), 按国际10-20系统扩展的64导电极帽记录EEG。以双侧乳突平均值为参考, 具体做法是,在记录中所有电极参考置于右乳突的一只参考电极, 离线分析时再次以置于左乳突的有效电极进行再参考, 即从各导联信号中减去该电极所记录信号的1/2。双眼外侧安置电极记录水平眼电(HEOG),左眼上下安置电极记录垂直眼电(VEOG)。每个电极处的头皮电阻保持在5 kΩ以下。滤波带通为0.05~100 Hz, 采样频率为1000 Hz, 采样增益(Gain)为20000。完成连续记录EEG后离线(off-line)处理数据, 用Scan软件校正VEOG和HEOG (Semlitsch,Anderer, Schuster, & Presslich, 1986), 并充分排除其他伪迹, 波幅大于±75 μV者被视为伪迹自动剔除。
本研究主要分析博弈结果评价阶段的ERP, 根据反馈刺激类型进行分段叠加, 刺激前200 ms (作为基线)开始到刺激呈现后1000 ms, 分析的ERP成分为FRN和P300。根据已有的研究, FRN在反馈刺激后250 ms左右达到峰值(Gehring & Willoughby,2002; Marco-Pallares et al., 2008; Yeung & Sanfey,2004), 我们首先将200~300 ms的平均波幅作为FRN进行重复测量方差分析。考虑到FRN成分可能会受到晚期正成分(P300)的影响, 我们也测量了200~300 ms之间最负峰值作为FRN (Rigoni, Polezzi,Rumiati, Guarino, & Sartori, 2010), 以及选择和解与进攻两种情境下的FRN差异波(dFRN, 输钱引发的波幅减去赢钱引发的波幅), 将两种情境下的dFRN效应定义为200~400 ms时间窗口内差异波的最负的峰值(Chen et al., 2012; Hajcak et al., 2005;Hajcak, Moser, Holroyd, & Simons, 2007)。P300测量的是300~600 ms时间内的平均波幅(Gu, Wu, Jiang,& Luo, 2011; Polezzi, Sartori, Rumiati, Vidotto, &Daum, 2010)。选择中线上的5个电极点进入统计分析:Fz/Fcz/Cz/CPz/Pz, 对FRN和P300进行2 (选择策略:和解, 进攻) × 2 (反馈效价:输, 赢) × 5 (电极:Fz/Fcz/Cz/CPz/Pz)的三因素重复测量方差分析(repeated measure ANOVA), 对FRN差异波(dFRN)进行2 (选择策略:和解, 进攻) × 5 (电极:Fz/Fcz/Cz/CPz/Pz)二因素重复测量方差分析。统计结果非球形性时采用Greenhouse-Gessisser法校正p
值, 多重比较时采用Bonferroni校正。3 结果
3.1 行为结果
在Chicken Game任务中, 和解与进攻的选择率分别为0.42 ± 0.08和0.58 ± 0.08。配对t
检验表明, 选择和解的比率选择小于选择进攻的比率,t
(21) = ‒4.721,p <
0.01。选择和解与选择进攻的平均反应时(RT
)分别为561.83 ± 164.12 ms和570.99 ±177.29 ms。选择两种策略的反应时之间没有显著差异,t
(21) = ‒0.826,p
> 0.05。为了探讨不同反馈类型对被试随后选择策略的影响, 分别计算被试接受4种反馈(RR, RA, AA,AR)后的下一个trials中和解的选择率。对四种反馈后和解的选择率进行单因素方差分析。结果表明,反馈类型的主效应显著,F
(3,63) = 19.89,p
< 0.001,η
= 0.49。事后比较表明, 在接受RR (两人都选择和解)反馈后, 被试选择和解的比率(M
= 0.55,SE
=0.039)大于接受RA与AA反馈后选择和解的比率(M
= 0.37,SE
= 0.041;M
= 0.22,SE
= 0.028); RR反馈后选择和解的比率(M
= 0.55,SE
= 0.039)与AR反馈后选择和解的比率(M
= 0.52,SE
= 0.030)之间没有显著差异(见图4A)。3.2 ERP结果
去除伪迹后进行ERP数据分析时, 四种反馈条件下叠加平均的有效trials数分别为:参与者A和参与者B都选择和解(RR)33个trials; 参与者A选择和解而参与者B选择进攻(RA)37个trials; 参与者A选择进攻而参与者B选择和解(AR)55个trials; 两个参与者都选择进攻(AA)45个trials。
3.2.1 FRN
首先对200~300 ms的平均波幅FRN进行2 (选择策略:和解, 进攻) × 2 (反馈效价:输, 赢) × 5 (电极:Fz/Fcz/Cz/CPz/Pz)的三因素重复测量方差分析。反馈效价的主效应显著,F
(1, 21) = 13.03,p
= 0.002,η
= 0.38, 输钱引发的FRN (M
= 2.99 μV,SE
= 0.86)比赢钱引发的FRN更大(M
= 5.14 μV,SE
= 1.12)。选择策略与反馈效价的交互作用显著,F
(1, 21) =13.99,p
= 0.001,η
= 0.40。简单效应分析表明, 选择和解时, 反馈效价主效应显著(F
(1, 21) = 19.91,p
<0.01), 输钱反馈引发的FRN (M
= 1.95 μV,SE
= 0.89)大于赢钱反馈引发的FRN (M
= 5.79 μV,SE
= 1.21)。选择进攻时, 反馈效价主效应不显著(F
(1, 21) =0.55,p
> 0.05)。此外, 上述FRN的交互作用也可以从另一个角度进行分解, 即在赢钱条件下, 选择策略的主效应不显著(F
(1, 21) = 2.76,p
= 0.11); 在输钱条件下, 选择策略的主效应显著, (F
(1, 21) =19.91,p
< 0.01), 选择和解反馈引发的FRN (M
=1.95 μV,SE
= 0.89)大于选择进攻反馈引发的FRN(M
= 4.03 μV,SE
= 0.96)。对200~300 ms的最负峰值FRN进行2 (选择策略:和解, 进攻) × 2 (反馈效价:输, 赢) × 5 (电极:Fz/Fcz/Cz/CPz/Pz)的三因素重复测量方差分析。选择策略与反馈效价的交互作用显著,F
(1, 21) = 4.91,p
= 0.038,η
= 0.19。但简单效应分析表明, 选择和解与选择进攻条件下, 反馈效价的主效应都没达到显著水平, (F
(1, 21) = 2.36,p
= 0.139;F
(1, 21) =0.76,p
= 0.394)。根据上述结果, 我们只选择了FRN波幅最大的电极点Fz上的峰值进行2 (选择策略:和解, 进攻) × 2 (反馈效价:输, 赢)的两因素重复测量方差分析。结果表明, 在Fz电极点上, 反馈效价的主效应显著,F
(1, 21) = 11.91,p
= 0.002,η
= 0.36,输钱引发的FRN (M
= ‒0.61 μV,SE
= 0.90)比赢钱引发的FRN更大(M
= 1.69 μV,SE
= 1.00)。选择策略与反馈效价的交互作用显著,F
(1, 21) = 16.32,p
=0.001,η
= 0.44。简单效应分析表明, 选择和解时,反馈效价主效应显著(F
(1, 21) = 22.59,p
< 0.01),输钱反馈引发的FRN (M
= ‒1.66 μV,SE
= 1.03)大于赢钱反馈引发的FRN (M
= 2.65 μV,SE
= 1.07)。选择进攻时, 反馈效价主效应不显著(F
(1, 21) = 0.15,p
> 0.05) (见图1与图2)。对FRN差异波进行2 (选择策略:和解, 进攻) × 5(电极:Fz/Fcz/Cz/CPz/Pz)二因素重复测量方差分析。选择策略的主效应显著,F
(1, 21) = 9.23,p
= 0.006,η
= 0.31, 选择和解策略下的dFRN波幅(M
= ‒6.21 μV,SE
= 0.82)显著大于进攻策略下的dFRN波幅(M
=‒3.70 μV,SE
= 0.83)。电极点的主效应显著,F
(4, 84) =5.78,p
= 0.018,η
= 0.22。事后比较表明, Fz以及FCz电极点上的dFRN波幅(M
= ‒6.28 μV,SE
= 0.84;M
=‒5.96 μV,SE
= 0.90)大于CPz电极点上的dFRN波幅(M
= ‒4.65 μV,SE
= 0.68,p
< 0.05) (见图1与图3)。3.2.2 P300
对P300进行2 (选择策略:和解, 进攻) × 2 (反馈效价:输, 赢) × 5 (电极:Fz/Fcz/Cz/CPz/Pz)的三因素重复测量方差分析。结果表明, 反馈效价的主效应显著,F
(1, 21) = 13.60,p
= 0.001,η
= 0.39, 赢钱引发的P300波幅(M
= 9.84 μV,SE
= 1.07)大于输钱引发的P300波幅(M
= 7.67 μV,SE
= 0.79)。选择策略与反馈效价的交互作用显著,F
(1, 21) = 16.43,p
=0.001,η
= 0.44。简单效应分析表明, 选择和解时,反馈效价主效应显著(F
(1, 21) = 19.32,p
< 0.01), 赢钱引发的P300波幅(M
= 10.39 μV,SE
= 1.30)大于输钱引发的P300波幅(M
= 6.57 μV,SE
= 0.73)。选择进攻时,反馈效价主效应不显著(F
(1, 21) = 1.04,p
> 0.05) (见图1与图2)。P300的交互作用也可以从另一个角度进行分解, 即在赢钱条件下, 选择策略的主效应不显著(F
(1, 21) = 1.10,p
= 0.31); 在输钱条件下, 选择策略的主效应显著, (F
(1, 21) = 6.74,p
= 0.017), 进攻反馈引发的P300 (M
= 8.77 μV,SE
= 1.04)大于和解反馈引发的P300 (M
= 6.57 μV,SE
= 0.73)。电极点的主效应显著,F
(4, 84) = 7.20,p
= 0.005,η
= 0.26。事后比较表明, Cz以及CPz电极点上的P300波幅(M
=9.91 μV,SE
= 0.896;M
= 9.63 μV,SE
= 0.96)大于Fz以及FCz电极点上的P300波幅(M
= 6.38 μV,SE
=0.82;M
= 8.23 μV,SE
= 0.90,p
< 0.05)。3.2.2 ERP波幅与和解选择率的相关与回归分析
对4种反馈引起的FRN以及P300波幅与接受4种反馈后和解的选择率进行相关分析。结果表明,RA反馈所引起的FRN波幅(Fz点)与接受RA反馈后下一个trials中和解的选择率呈显著的负相关(r
= ‒0.55,p
= 0.009)。回归分析表明, RA反馈所引起的FRN波幅越大的被试, 在接受RA反馈的下一个trials中选择和解的比率越大, FRN波幅可以解释RA反馈后和解选择率30%的变异量(R
= 0.30,F
(1,20) = 8.41,p
= 0.009)。对RA反馈引起的FRN波幅与总的和解选择率进行相关分析, 结果表明, RA反馈所引起的FRN波幅与总的和解选择率也呈显著的负相关(r
=‒0.59,p
= 0.004)。其他三种反馈(RR, AA, AR)引起的FRN波幅与接受该反馈后和解的选择率(或者总的和解选择率)之间相关不显著。此外, 四种反馈引起的P300波幅与接受四种反馈后和解的选择率之间相关不显著(见图4B)。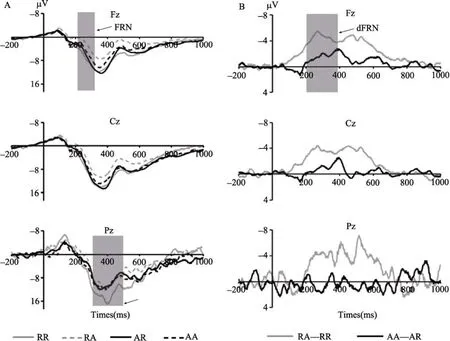
图1 (A)四种反馈结果在Fz, Cz和Pz电极点上总平均的波形图。RR表示被试和对方都选择和解(+10/+10); RA表示被试选择和解对方选择进攻(‒10/+30); AR表示被试选择进攻对方选择和解(+30/‒10); AA表示被试和对方都选择进攻(‒30/‒30)。(B)选择和解与选择进攻条件下的dFRN在Fz, Cz和Pz电极点上总平均的波形图。RA-RR表示和解条件下输钱与赢钱的FRN差异波, AA-AR表示选择进攻条件下输钱与赢钱的FRN差异波。

图2 (A) Fz电极点上和解与进攻条件下输钱与赢钱引发的FRN (峰值)的均值与标准误。(B) Fz /Fcz /Cz /CPz /Pz 电极点上和解与进攻条件下输钱与赢钱引发的P300的均值与标准误。
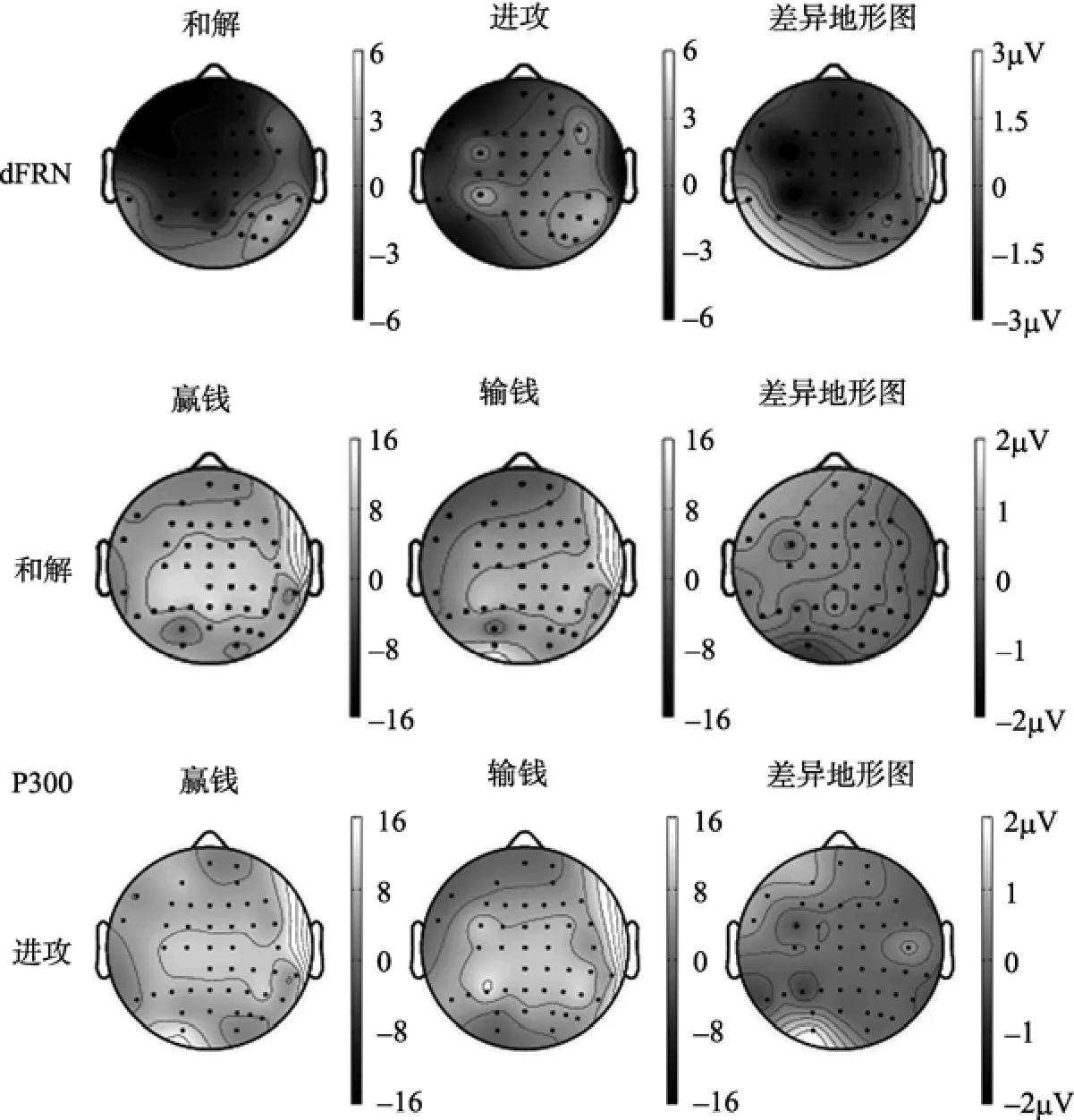
图3 和解与进攻条件下FRN差异波dFRN (250 ms)以及两种条件下赢钱和输钱的P300 (300~600 ms)的脑地形图。最右一列的差异地形图是指前两列中两种条件相减的后的地形图分布。
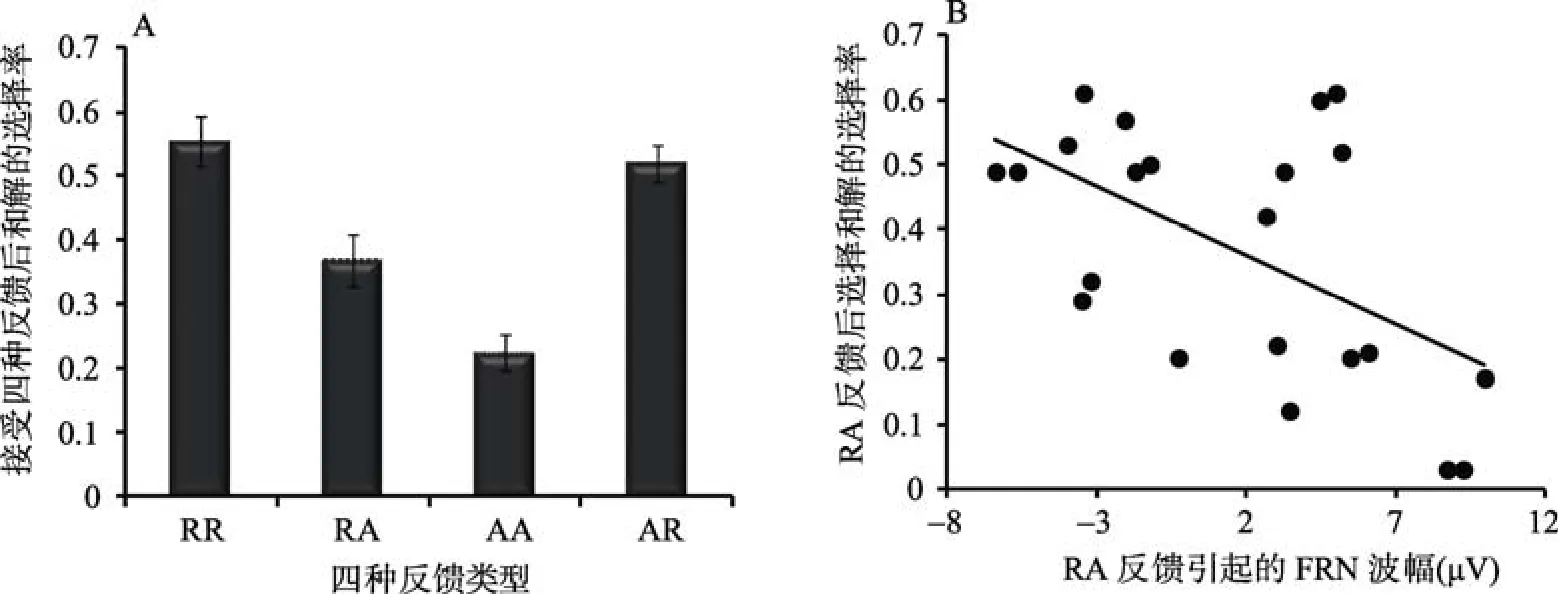
图4 (A) 接受4种反馈之后被试选择和解的比率, RR表示两人都和解, RA被试选择和解对方选择进攻, AA表示两人都选择进攻, AR表示被试选择进攻对方选择和解。(B) RA反馈引起的FRN波幅与接受RA反馈后被试选择和解比率的相关散点图。
4 讨论
目前为止, 采用ERPs技术深入考察人际合作与冲突如何影响个体决策选择之后结果评价过程的研究尚显不足。本研究结合ERPs技术与Chicken Game任务, 采用FRN和P300作为电生理指标, 考察了合作与冲突预期对社会博弈中结果评价的影响。研究进一步揭示了个体对合作与冲突结果的认知评价过程, 为进一步了解社会决策加工过程提供了一定的电生理学证据。下面分别从FRN和P300这两个脑电指标讨论人际合作与冲突对社会博弈中结果评价过程的影响。
4.1 FRN反映主观预期与实际结果之间的冲突强度以及相对损失大小
ERP的结果表明, FRN选择策略的主效应不显著。本研究中选择策略也对应着数量大小维度, 这表明FRN波幅并不直接受到收益或者损失大小的影响, 这与前人的一些研究结果是一致的, 即FRN对反馈大小这个维度不敏感(Gehring & Willoughby,2002; Yeung & Sanfey, 2004)。但本研究发现, FRN的波幅受到选择策略和反馈效价的共同影响, 简单效应分析表明, 只有在选择和解时输钱引发的FRN波幅才显著大于赢钱引发的FRN波幅, 而选择进攻时输赢所引起的FRN波幅差异并没有达到显著。FRN差异波(dFRN)的结果也与上述结果一致, 选择和解条件下的dFRN波幅大于选择进攻条件下的dFRN波幅。根据以往的研究, FRN反映的是大脑ACC区域对负性结果的加工(Gehring & Willoughby,2002; Masaki et al., 2006; Yu et al., 2007)。从另一个角度对FRN的交互作用进行简单效应分析表明,FRN波幅的差异主要是源自于输钱条件下, 选择和解与进攻之间的差异。选择和解输钱的FRN大于选择进攻输钱的FRN, 这表明个体对负性结果的评价并不是由客观的损失大小所决定, 个体的主观预期会显著影响对负性结果的评价过程。在本研究中,被试选择和解, 表明此时被试愿意与对方一起合作共赢, 因此主观上更加期待对方选择和解。当电脑给予的反馈是对方选择进攻时, 这一结果与被试的预期差异很大; 此时被试可能会对该结果感到失望与懊恼, 因为如果对方也选择合作, 两人就会都赢得到10元。因此, 被试选择和解对方选择进攻的结果, 引起的认知或者情绪冲突会更加强烈, 所引发的FRN波幅也更大。
在非社会互动情境中, FRN一般是由负性反馈引发, 并且与反馈大小无关(Hajcak, Moser, Holroyd,& Simons, 2006)。强化学习理论以及冲突监控理论认为, FRN反映的是实际结果与预期之间的差异。Oliveira, McDonald和Goodman (2007)提出的预期偏离假说(expectancy deviation hypothesis)认为无论反馈的效价如何, 只有当实际结果与预期不匹配时才会产生明显的FRN。如果个体对事件结果的预期是负性的, 而实际结果却是正性反馈也会产生明显的FRN。也就是说FRN产生根本原因是实际结果与预期之间的不匹配, 而不是效价上的差异。本研究中选择进攻时输赢所引起的FRN波幅差异并未达到显著。出现这种结果的可能原因是, 选择进攻之后, 虽然也有效价(输赢)上的差异, 但在选择进攻情境下被试对他人选择合作预期可能并不强烈, 或者说此时被试会事先猜测对方可能也选择进攻。因此, 被试选择进攻时, 不管对方选择何种策略, 被试可能已经做好最坏的打算, 此时的输钱反馈与被试主观预期的冲突并不那么强烈, 所以输钱引发的FRN并没有显著大于赢钱所引发的FRN。与本研究结果一致, Long等人(2012)研究发现, 信任感可以调节大脑对结果的反应。在信任条件下输赢之间的FRN差异显著; 而不信任条件下, 输赢之间的FRN差异不显著。研究者认为在信任情境下, 输赢之间的FRN差异反映了对社会预期违反的检测。而不信任情境下之所以出现FRN效应显著减弱, 是因为不信任激活了被试的超然感(a sense of aloofness), 在这种情境下被试根本或者较少地对随后的事件产生预期。此外, 我们推测本研究中选择进攻条件下FRN效应减弱也可能与被试的个体差异有关。我们对每个被试的AVG观测发现, 有些被试选择进攻之后的输赢之间的FRN差异非常明显, 而有些被试择进攻后输赢之间的FRN则没有差异, 这种结果背后的个体差异机制还有待进一步的研究。
已有研究表明, 预期会影响FRN波幅, 与预期不一致的负性结果会产生更大的FRN波幅(Gibson,Krigolson, & Holroyd, 2006; Holroyd & Coles, 2002;Wu & Zhou, 2009), 但也有研究发现预期对FRN波幅没有影响, 而只影响P300的波幅(Cohen & Ranganath,2007; Hajcak et al., 2005; Hajcak, Moser, Holroyd, &Simons, 2007)。根据本研究结果, 我们认为, 预期对FRN的影响取决于主观预期和实际结果之间的冲突强度, 两者之间冲突越大, 所引起的FRN波幅越大。Zhou等人(2010)研究发现, FRN的波幅受到个体的行动与否的影响, 相对于不行动条件, 在行动条件下输钱比赢钱引发更大的FRN波幅。研究者认为行动增加了个体对结果的主观期待, 因此在行动条件下引发的FRN更大。也有研究表明个体所承担的责任大小会影响FRN的波幅, 对于同样的负性结果, 当个体在决策时承担的责任越大, 所引发的FRN的波幅就越大(Li et al., 2010)。这些结果表明,对负性结果的评价受到个体对反馈结果主观认知或者体验的影响。结合上述研究结果, 我们认为在本研究中合作行为增加了个体对博弈结果的主观预期, 因此, 在合作情境下, 损失与收益引起的FRN差异更大。
FRN波幅与和解选择率之间的相关分析表明,RA反馈引起的FRN波幅越大的被试, 在接受RA反馈后下一个trials中选择和解比率以及总体选择和解的比率越大。这表明RA反馈引起的FRN可以在一定程度上预测个体随后的决策行为。Boksem和De Cremer (2010)采用最后通牒任务研究了公平感与FRN之间的关系, 结果表明, 相对于公平提议, 不公平提议引发了更大的FRN, 并且这种效应在那些更在意公平(公平感更强)的被试中更加明显。如前所述, 本研究中RA反馈是博弈对方违背个体的合作意愿而选择进攻的一种结果; 因此, RA反馈引起的FRN波幅反映了个体对他人合作行为的社会预期强度, RA反馈引发的FRN波幅越大, 表明被试对他人合作的预期越强烈, 或者说这些被试更愿意相信对方表现出合作行为。因此, RA反馈引起的FRN波幅越大的被试在随后的博弈中(以及总的博弈过程中)会更多地选择合作行为。
此外, 根据本研究的FRN结果, 我们认为社会博弈中结果评价的FRN成分也可能反映了个体对博弈结果进行社会比较的过程。根据社会比较理论(Social Comparison Theory), 个体会自觉或者不自觉地将自己与他人进行比较(Festinger, 1954)。这种比较会影响到个体对结果的评价, 同样是损失10元,如果此时别人得到30元, 个体对损失的负性体验就会增加; 而如果此时别人也损失同样的钱数, 损失引起的负性体验就会降低。Wu, Zhou, Van Dijk,Leliveld和Zhou (2011)采用最后通牒任务考察了社会比较对结果公平性评价的影响, 研究结果表明,对不公平提议的评价的确受到社会比较的影响。在本研究中, 如果被试和对方都选择进攻, 虽然两人都损失30元, 但从相对损失来看, 他们相对损失差异是零。而当被试选择和解对方选择进攻时, 被试损失10元但对方得到30元, 对被试来说相对损失差异就是40元。被试可能更加难以接受后者, 因此引发的FRN波幅更大。总之, FRN结果表明, 社会博弈情境下的FRN反映的并不是反馈结果本身的性质与量, 而是主观预期与反馈评价的符合程度。反馈信息虽然是客观的, 但只有主观觉知后才能起作用。产生FRN的系统在评价得失时包括着对社会情境因素的认知, 换而言之, 产生FRN的系统不是评价收益或者损失的绝对价值, 而是评价该收益或者损失相对于可能的收益或损失的总体价值。
4.2 P300反映金钱奖励与社会奖励的加工过程
结果表明, 赢钱引发的P300比输钱引发的P300更大, 这重复了先前的一些研究结果(Hajcak et al., 2005; Leng & Zhou, 2010; Wu & Zhou, 2009;Yeung et al., 2005)。很多关于结果评价的ERP研究都已涉及到P300, 这些研究一致认为P300和FRN都是反映结果评价的重要脑电成分。有些研究表明,P300只对反馈的大小敏感, 而对反馈效价不敏感(Yeung & Sanfey, 2004)。但最近的很多研究也发现反馈效价也可以影响P300的波幅, 赢钱会引发更大的P300(Hajcak et al., 2005; Wu & Zhou, 2009;Yeung et al., 2005; Zhou et al., 2010)。对于赢钱引发的P300大于输钱引发的P300的解释, 目前并没有一致的结论。一般认为, P300和决策或结果评价中的注意资源分配(Gray, Ambady, Lowenthal, & Deldin,2004)以及高水平的动机/情感评价(Leng & Zhou,2010; Nieuwenhuis et al., 2005; Yeung & Sanfey, 2004)有关。
除了发现赢钱引发的P300大于输钱引发的P300, 更有意义的是, 本研究发现反馈效价对P300的影响受到人际合作与冲突的调节, 选择和解后,赢钱和输钱的P300差异显著; 选择进攻后, 两者差异没有达到显著。结果评价中P300所代表的意义目前还没有一致的结论, 很多研究者认为结果评价中P300与社会性的注意资源分配或者反馈引起动机和意义有关(Gray et al., 2004; Leng & Zhou, 2010;Linden, 2005; Yeung & Sanfey, 2004)。根据本研究P300的结果, 我们认为社会博弈结果评价中的P300可能反映了金钱奖励与社会奖励的加工过程。在本研究中, 当两人都选择和解时, 双方都赢10元,这是一种互利共赢的博弈结果, 这一结果引起的P300波幅更大, 表明个体认为互利共赢的结果比单纯赢钱更有意义。Rilling等人(2002)采用囚徒困境任务发现, 个体相互合作时激活了伏隔核(nucleus accumbens)、腹内侧额叶/眶额皮层(ventromedial frontal/orbitofrontal cortex)等与奖励加工有关的脑区。也有研究采用fMRI发现, 社会奖励与金钱奖励激活了相同的脑区(Rademacher et al., 2010)。这表明个体把合作共赢的结果作为一种社会奖励, 在一定程度上, 其意义与金钱奖励的意义是等价的。Elliott等人(2006)的研究发现合作也是一种奖励加工过程, 相互合作所得到奖励更强的激活了内侧眶额皮层(medial orbitofronta cortex), 通过合作得到的奖励比单独做得到的奖励更有价值。
本研究从结果评价的角度印证了上述研究结果, 合作条件下赢钱与输钱引发的P300差异大于进攻条件下赢钱与输钱引发的P300差异。这表明社会奖励与金钱有着同样的价值, 甚至更大的价值。合作共赢带来的社会奖励对人类的生存和发展有着重要的意义, 在现实生活中, 很多时候我们需要与他人共同进行一些任务, 人类这种与他人为了同一个目标而共同工作的能力对人类的群居生存具有重要意义(Newman-Norlund, Noordzij, Meulenbroek,& Bekkering, 2007)。基于互惠利他的合作不仅仅在一小部分物种中演化, 也构成了人类社会生活的核心行为准则(Rilling et al., 2002), 人们与他人寻求合作是为了获得共同的利益, 以达到合作共赢的目的, 只有这样才能更好地适应人类社会的发展。因此, 合作共赢的结果不仅具有物质奖励的属性, 还有社会奖励的价值。个体在评价社会博弈结果的过程中, 既加工其本身的物质意义, 也加工其背后隐含的社会价值, 并且这一加工过程可能反映在结果评价晚期的P300成分上。
Boksem, M. A., & De Cremer, D. (2010). Fairness concerns predict medial frontal negativity amplitude in ultimatum bargaining.Social Neuroscience, 5
(1), 118–128.Camerer, C. F. (2003).Behavioral game theory: Experiments in strategic interaction
. Princeton: Princeton University Press.Chen, J., Zhong, J., Zhang, Y., Li, P., Zhang, A., Tan, Q., & Li,H. (2012). Electrophysiological correlates of processing facial attractiveness and its influence on cooperative behavior.Neuroscience Letters, 517
(2), 65–70.Cohen, M. X., & Ranganath, C. (2007). Reinforcement learning signals predict future decisions.Journal of Neuroscience, 27
(2), 371–378.Decety, J., Jackson, P. L., Sommerville, J. A., Chaminade, T.,& Meltzoff, A. N. (2004). The neural bases of cooperation and competition: An fMRI investigation.NeuroImage,23
(2), 744–751.Elliott, R., Völlm, B., Drury, A., McKie, S., Richardson, P., &William Deakin, J. (2006). Co-operation with another player in a financially rewarded guessing game activates regions implicated in theory of mind.Social Neuroscience, 1
(3-4),385–395.Festinger, L. (1954). A theory of social comparison processes.Human Relations, 7
(2), 117–140.Fukui, H., Murai, T., Shinozaki, J., Aso, T., Fukuyama, H.,Hayashi, T., & Hanakawa, T. (2006). The neural basis of social tactics: An fMRI study.NeuroImage, 32
(2), 913–920.Gehring, W. J., & Fencsik, D. E. (2001). Functions of the medial frontal cortex in the processing of conflict and errors.Journal of Neuroscience, 21
(23), 9430–9437.Gehring, W. J., & Willoughby, A. R. (2002). The medial frontal cortex and the rapid processing of monetary gains and losses.Science, 295
(5563), 2279–2282.Gibson, J., Krigolson, O., & Holroyd, C. (2006). Sensitivity of the feedback error-related negativity to reward probability.Psychophysiology, 43
, S41–S42.Gray, H. M., Ambady, N., Lowenthal, W. T., & Deldin, P.(2004). P300 as an index of attention to self-relevant stimuli.Journal of Experimental Social Psychology, 40
(2),216–224.Gu, R., Wu, T., Jiang, Y., & Luo, Y. J. (2011). Woulda, coulda,shoulda: the evaluation and the impact of the alternative outcome.Psychophysiology, 48
(10), 1354–1360.Hajcak, G., Holroyd, C. B., Moser, J. S., & Simons, R. F.(2005). Brain potentials associated with expected and unexpected good and bad outcomes.Psychophysiology,42
(2), 161–170.Hajcak, G., Moser, J. S., Holroyd, C. B., & Simons, R. F.(2006). The feedback-related negativity reflects the binary evaluation of good versus bad outcomes.Biological Psychology, 71
(2), 148–154.Hajcak, G., Moser, J. S., Holroyd, C. B., & Simons, R. F.(2007). It's worse than you thought: The feedback negativity and violations of reward prediction in gambling tasks.Psychophysiology, 44
(6), 905–912.Hewig, J., Kretschmer, N., Trippe, R. H., Hecht, H., Coles, M.G., Holroyd, C. B., & Miltner, W. H. (2011). Why humans deviate from rational choice.Psychophysiology, 48
(4),507–514.Holroyd, C. B., & Coles, M. G. H. (2002). The neural basis of human error processing: reinforcement learning, dopamine,and the error-related negativity.Psychological Review,109
(4), 679–708.Holroyd, C. B., Hajcak, G., & Larsen, J. T. (2006). The good,the bad and the neutral: Electrophysiological responses to feedback stimuli.Brain Research, 1105
(1), 93–101.Jia, S., Li, H., Luo, Y., Chen, A., Wang, B., & Zhou, X. (2007).Detecting perceptual conflict by the feedback-related negativity in brain potentials.Neuroreport, 18
(13), 1385–1388.Leng, Y., & Zhou, X. (2010). Modulation of the brain activity in outcome evaluation by interpersonal relationship: An ERP study.Neuropsychologia, 48
(2), 448–455.Li, P., Han, C., Lei, Y., Holroyd, C. B., & Li, H. (2011). Responsibility modulates neural mechanisms of outcome processing: An ERP study.Psychophysiology, 48
(8), 1129–1133.Li, P., Jia, S., Feng, T., Liu, Q., Suo, T., & Li, H. (2010). The influence of the diffusion of responsibility effect on outcome evaluations: Electrophysiological evidence from an ERP study.NeuroImage, 52
(4), 1727–1733.Linden, D. E. (2005). The P300: Where in the brain is it produced and what does it tell us?The Neuroscientist, 11
(6),563–576.Long, Y., Jiang, X., & Zhou, X. (2012). To believe or not to believe: Trust choice modulates brain responses in outcome evaluation.Neuroscience, 200
, 50–58.Ma, Q., Shen, Q., Xu, Q., Li, D., Shu, L., & Weber, B. (2011).Empathic responses to others' gains and losses: An electrophysiological investigation.Neuroimage, 54
(3),2472–2480.Marco-Pallares, J., Cucurell, D., Cunillera, T., García, R.,Andres-Pueyo, A., Münte, T. F., & Rodríguez-Fornells, A.(2008). Human oscillatory activity associated to reward processing in a gambling task.Neuropsychologia, 46
(1),241–248.Masaki, H., Takeuchi, S., Gehring, W. J., Takasawa, N., &Yamazaki, K. (2006). Affective-motivational influences on feedback-related ERPs in a gambling task.Brain Research,1105
(1), 110–121.Newman-Norlund, R. D., Noordzij, M. L., Meulenbroek, R. G.,& Bekkering, H. (2007). Exploring the brain basis of joint action: Co-ordination of actions, goals and intentions.Social Neuroscience, 2
(1), 48–65.Nieuwenhuis, S., Aston-Jones, G., & Cohen, J. D. (2005).Decision making, the P3, and the locus coeruleus-norepinephrine system.Psychological Bulletin, 131
(4), 510–532.Oliveira, F. T., McDonald, J. J., & Goodman, D. (2007).Performance monitoring in the anterior cingulate is not all error related: Expectancy deviation and the representation of action-outcome associations.Journal of Cognitive Neuroscience, 19
(12), 1994–2004.Platt, M. L. (2002). Neural correlates of decisions.Current Opinion in Neurobiology, 12
(2), 141–148.Polezzi, D., Sartori, G., Rumiati, R., Vidotto, G., & Daum, I.(2010). Brain correlates of risky decision-making.Neuro-Image, 49
(2), 1886–1894.Rademacher, L., Krach, S., Kohls, G., Irmak, A., Grunder, G.,& Spreckelmeyer, K. N. (2010). Dissociation of neural networks for anticipation and consumption of monetary and social rewards.Neuroimage, 49
(4), 3276–3285.Rapoport, A., & Chammah, A. (1966). The game of chicken.American Behavioral Scientist, 10
(3), 10–28.Rigoni, D., Polezzi, D., Rumiati, R., Guarino, R., & Sartori, G.(2010). When people matter more than money: An ERPs study.Brain Research Bulletin, 81
(4), 445–452.Rilling, J., Gutman, D., Zeh, T., Pagnoni, G., Berns, G., &Kilts, C. (2002). A neural basis for social cooperation.Neuron, 35
(2), 395–405.Sato, A., Yasuda, A., Ohira, H., Miyawaki, K., Nishikawa, M.,Kumano, H., & Kuboki, T. (2005). Effects of value and reward magnitude on feedback negativity and P300.Neuroreport, 16
(4), 407–411.Semlitsch, H. V., Anderer, P., Schuster, P., & Presslich, O.(1986). A solution for reliable and valid reduction of ocular artifacts, applied to the P300 ERP.Psychophysiology, 23
(6),695–703.Veen, V., & Carter, C. S. (2002). The timing of action-monitoring processes in the anterior cingulate cortex.Journal of Cognitive Neuroscience, 14
(4), 593–602.Wong, R. Y., & Hong, Y. Y. (2005). Dynamic influences of culture on cooperation in the prisoner's dilemma.Psychological Science, 16
(6), 429–434.Wu, Y., & Zhou, X. (2009). The P300 and reward valence,magnitude, and expectancy in outcome evaluation.Brain Research, 1286
, 114–122.Wu, Y., Zhou, Y., Van Dijk, E., Leliveld, M. C., & Zhou, X.(2011). Social comparison affects brain responses to fairness in asset division: An ERP study with the ultimatum game.Frontiers in Human Neuroscience, 5
, 131.Yeung, N., Holroyd, C. B., & Cohen, J. D. (2005). ERP correlates of feedback and reward processing in the presence and absence of response choice.Cerebral Cortex,15
(5), 535–544.Yeung, N., & Sanfey, A. G. (2004). Independent coding of reward magnitude and valence in the human brain.Journal of Neuroscience, 24
(28), 6258–6264.Yu, R. J., Luo, Y. J., Ye, Z., & Zhou, X. L. (2007). Does the FRN in brain potentials reflect motivational/affective consequence of outcome evaluation?Progress in Natural Science, 17
, 136–143.Zhou, Z., Yu, R., & Zhou, X. (2010). To do or not to do?Action enlarges the FRN and P300 effects in outcome evaluation.Neuropsychologia, 48
(12), 3606–3613.猜你喜欢
心理学探新(2022年1期)2022-06-07 09:16:02
环球时报(2022-04-16)2022-04-16 14:38:15
癫痫与神经电生理学杂志(2022年6期)2022-02-10 13:55:46
井冈教育(2020年6期)2020-12-14 03:04:32
价值工程(2018年25期)2018-09-26 10:14:36
猪业科学(2018年5期)2018-07-17 05:55:18
西南交通大学学报(2016年4期)2016-06-15 20:29:36
华北电力大学学报(自然科学版)(2016年3期)2016-04-25 05:06:56
中国当代医药(2015年8期)2015-03-01 02:01:55
浙江人大(2014年6期)2014-03-20 16:20:40
