
翻堆对蛋鸡粪堆肥甲烷排放的影响
2014-01-21 01:16刘俊专廖新俤吴银宝
家畜生态学报 2014年2期
刘俊专,王 燕,廖新俤,吴银宝,胖 是
(华南农业大学动物科学学院,广东广州510642)
随着全球温室效应的日益加剧,温室气体研究备受各国学者关注。甲烷(CH4)作为重要的温室气体之一,其增暖潜势约是二氧化碳(CO2)的23 倍,对全球变暖的贡献率为19%,仅次于CO2(55%),且甲烷排放每年正以0.9%的速度持续增加[1]。农业生产是CH4的重要排放源,40%的CH4排放源于农业生产[2]。其中畜禽粪便是最为重要的排放源之一,全球约有9%的生物性CH4源于畜禽粪便的管理和应用过程中[3-5]。因此研究畜禽粪便管理利用过程中甲烷排放对全球甲烷减排,减轻全球温室效应具有重要意义。
堆肥是实现畜禽粪便无害化、减量化和资源化的主要途径,同样也是减少畜禽粪便温室气体排放的重要方式[6-7]。在堆肥过程中甲烷排放约占初始总有机碳的0.01%~8.0%[4]。翻堆可以促进堆体升温、物料降解及腐熟,但也可能影响CH4 排放。近年研究表明,翻堆可显著减少CH4的排放[8-9];而也有学者研究发现,翻堆会增加CH4的排放[6,10]。这可能是因为不同研究采用的翻堆频率不同,从而导致翻堆对CH4排放产生不同影响。但目前鲜见专门研究翻堆对CH4排放影响的相关报道。因此,本试验通过设定不同的翻堆频率,研究其对CH4排放所产生的影响,并对影响CH4生成的微生物菌群进行分析,为堆肥过程中合理翻堆及CH4减排提供依据。
1 材料与方法
1.1 堆肥原料与试验分组
新鲜的蛋鸡粪便采自中山市白石鸡场产蛋鸡舍,传输带收集粪便、打包,第2天运输至试验现场。锯末购自河源市东源县某木材加工厂。堆肥原料主要成分见表1。将鸡粪和锯末按鲜重湿重比为7.64︰1,控制堆体物料初始重约150kg,初始C/N 为10左右,含水率68%。
按照4 次/d、2 次/d、1 次/d、1 次/3d、1 次/6d和不翻堆6个不同翻堆频率,分为A、B、C、D、E 和F(对照组)6个处理。A 组每天09︰00、15︰00、21︰00和03︰00翻堆,B 组每天09︰00和21︰00翻堆,其他各组在翻堆当天的09︰00翻堆。试验时间为40d。
1.2 堆肥设备
采用自行设计的动态箱式堆肥试验装置,内部配有搅拌装置以及温度探头,对堆肥过程中的温度实时监测。堆肥箱长×宽×高为60cm×60cm×80cm,体积为288L。箱体结构见图1。

表1 堆肥原料的主要成分Table 1 Main component of composting material
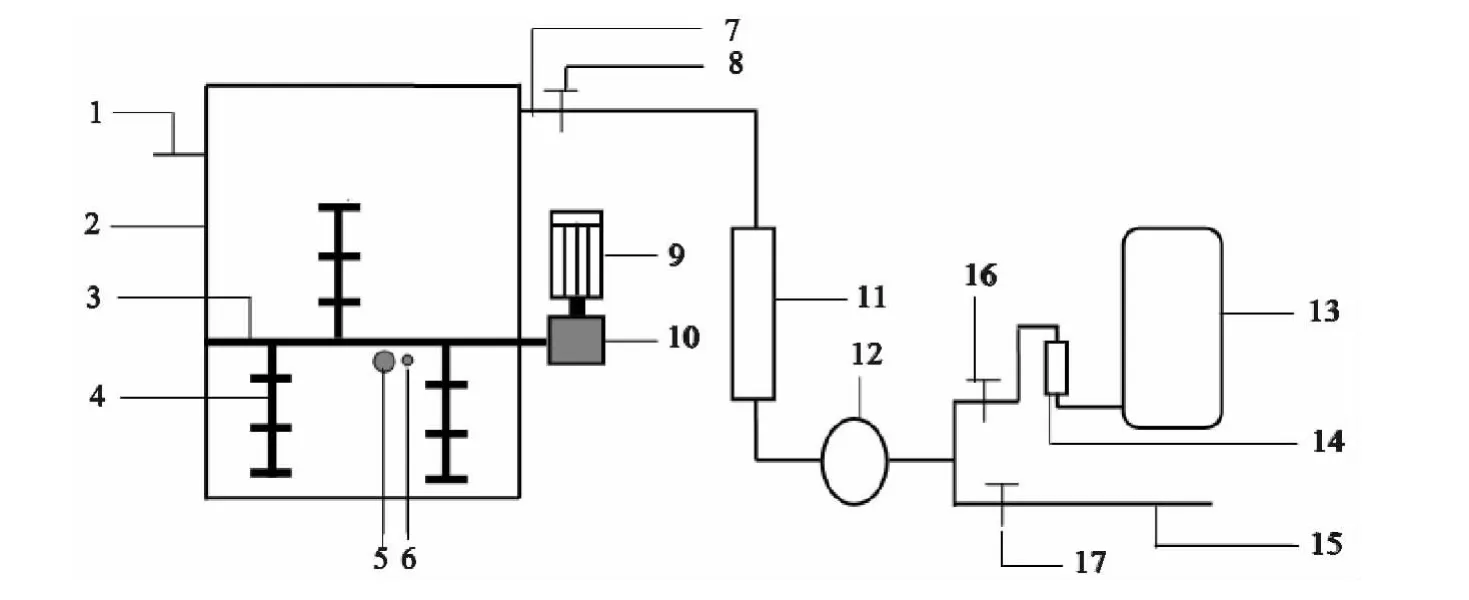
图1 动态箱式堆肥试验装置结构图Fig.1 Structure diagram of dynamic box composting test device
1.3 测定指标及方法
1.3.1 气样采集及CH4浓度测定 堆肥前15d,每天分别在09︰00~10︰00、14︰00~15︰00和20︰00~21︰00 3个时间段,收集1h内由箱体出气口排出的所有气体,混匀后用1L 铝箔袋采集气样,用于测定CH4浓度。
采用安捷伦气相色谱仪(7890A)FID 检测器测定CH4浓度,色谱柱采用Agilent19091P-Q04。CH4排放量的计算公式如下:
F=(C1-C2)V
式中:F 为CH4排放量(mL),C1为出气口CH4浓度(μL/L),C2为进气口CH4浓度(μL/L),V 为出气口总流量(m2)。
1.3.2 总DNA 提取 堆肥样品微生物总DNA 采用OMEGA 公司E.Z.N.A.TM Soil DNA Kit试剂盒提取。
1.3.3 产甲烷菌普通PCR 扩增 以总DNA 为模板,采用巢式PCR 法进行扩增,试验所用引物均由北京奥科鼎盛生物科技有限公司合成。首先采用Met86F和Met1340R 为引物(表2)[11],扩增产甲烷菌16SrRNA 基因片段,目的片段大小为1 254bp,反应体系采用25μL 的扩增体系,其中Premix Ex TaqTM Version2.0 12.5 μL,引物Met86F(10 pmol/μL)0.5μL,引物Met1340R(1 0pmol/μL)0.5μL,DNA 模板0.5μL,加无菌ddH2O 至体积25μL。PCR 条件为95 ℃预变性3min,95 ℃变性30s,54℃复性30s,72℃延伸80s。34个循环,最后72 ℃延伸10min,然后4 ℃保温,反应结束后取5μL PCR 产物用2%琼脂糖凝胶于130V 电泳23 min,然后通过凝胶成像系统观察PCR 产物,并拍照记录,确认PCR 扩增出目的产物后用于下一步PCR 的模板;再采用产甲烷菌的特异性引物对GC-Arc344F 和519R(表2)[12],扩增产甲烷菌16S rRNA 基因上的V3可变区,用于DGGE 分析。反应体系采用50μL 的扩增体系,其中Premix Ex TaqTM Version2.0 25.0μL,引物GC-Arc344F(10 pmol/μL)1.0μL,引物Arc519R(10pmol/μL)1.0 μL,DNA 模板1.0μL,加无菌ddH2O 补充体积到50μL。PCR条件为95℃预变性3min,95℃变性15 s,56℃复性30s,72℃延伸30s。34个循环,72℃延伸7min,然后4℃保温,反应结束后取5μL PCR产物用2%琼脂糖凝胶于130V 电泳23min,然后通过凝胶成像系统观察PCR产物,并拍照记录。
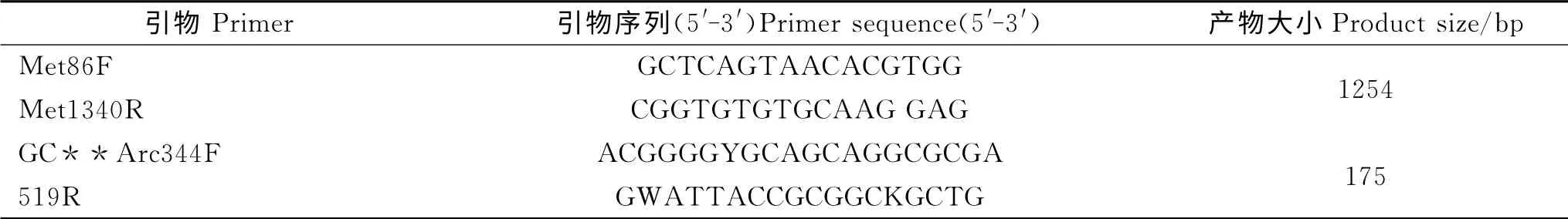
表2 产甲烷菌的引物序列Table 2 Primer sequence of methanogens
1.3.4 实时荧光定量PCR 将已知浓度的产甲烷菌质粒DNA 用双蒸灭菌水按1∶10 倍稀释8个梯度,每个梯度3个重复,同时作3个阴性对照,进行实时荧光定量PCR 反应。采用引物GC-Arc344F 和519R(表2)[12],采用20μL反应体系,其中10pmol/μL的引物各0.5μL,2×SYBR Green qPCR Supermix 10μL,超纯水8μL。95 ℃预变性1min,变性15s,52℃退火30s,72℃延伸25s,循环40次。反应结束后,由溶解曲线判定PCR反应的特异性,根据荧光曲线的Ct值以及标准曲线技术定量结果[13]。
堆肥样品中产甲烷菌拷贝数计算公式如下:

1.4 数据分析
试验数据采用SPSS17.0软件进行统计分析,多重比较采用邓肯极差检验法,显著水平P值设为0.05。
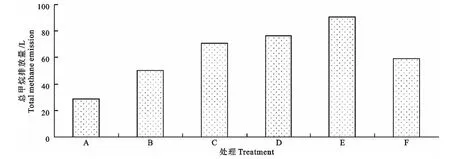
图2 各处理甲烷排放总量Fig.2 Total methane emission of different treatment
2 结果与分析
2.1 CH4 排放总量
由图2可见,翻堆频率为4次/d(处理A)、2次/d(处理B)、1次/d(处理C)、1次/3d(处理D)、1次/6d(处理E)和不翻堆(处理F)时,甲烷排放总量分别为28.60L、50.07L、70.85L、76.48L、90.74 L和59.33L。本试验中,甲烷排放量最高的翻堆频率为1 次/6d(E 组),最低的翻堆频率为4 次/d(A 组)。与不翻堆相比,在高翻堆频率下(4 次/d和2次/d),甲烷的排放量减少,且随翻堆频率的增加而甲烷排放量降低。
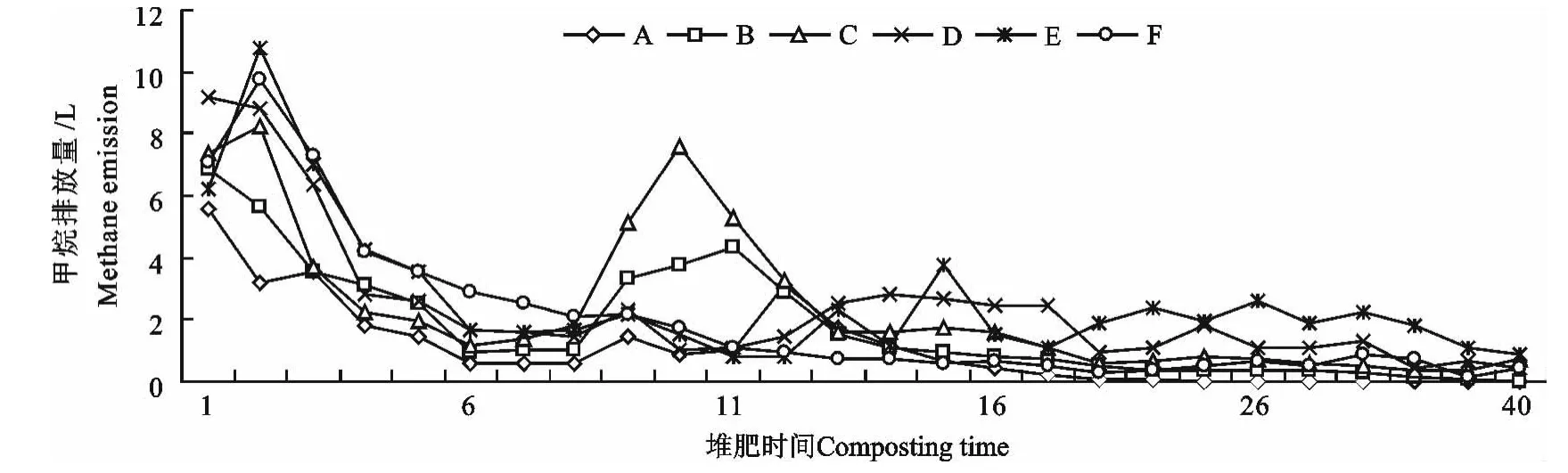
图3 甲烷排放量随堆肥时间的变化Fig.3 Methane emissions in different composting time
2.2 CH4 排放量随堆肥时间变化
如图3所示,各处理在堆肥初期甲烷排放量最大,这与很多学者的研究结果一致[6-7,14-15]。随堆肥时间的延长,甲烷排放量基本呈逐渐下降的趋势,但B、C组在9至11d出现一个甲烷排放小高峰,可能与B、C组此时温度上升较快有关。
堆肥前12d,各组甲烷排放量大小顺序为A<B<D<E<F<C,方差分析表明:堆肥前12d各组甲烷排放量差异不显著(P>0.05)。但从堆肥第13 d至结束各组甲烷排放量大小顺序为A<B<F<C<D<E,A 组显著低于C、D、E 组(P<0.05),B 组显著低于D、E组(P<0.05),F组显著低于D、E 组(P<0.05),C 组显著低于D、E 组(P<0.05)。上述分析表明,翻堆对堆肥升温期甲烷的排放没有显著影响,但在堆肥高温期和降温期,高频翻堆会显著降低甲烷的排放,翻堆频率越低甲烷排放量越大。
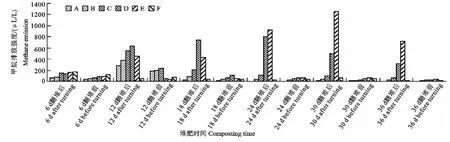
图4 翻堆后和翻堆前1h甲烷排放强度对比Fig.4 Comparison of methane emission before and after turning within 1hduring composting
2.3 翻堆前后CH4 排放强度差异
图4是不同天数翻堆前后1h内收集的气体中甲烷浓度的比较。由图4知,各处理翻堆后甲烷浓度均高于翻堆前,且翻堆频率较低(1 次/3d 和1次/6d)时,翻堆前后浓度差别最大。说明翻堆会促进甲烷的挥发,翻堆后甲烷排放强度会增大,在翻堆频率较低时,翻堆后甲烷排放强度增加越多。
2.4 产甲烷菌多样性
堆肥过程中各组产甲烷菌多样性指数H′的变化见表3。由表3可知,各组产甲烷菌多样性指数H′的变化趋势基本一致,在堆肥开始时各组产甲烷菌多样性指数H′为2.38~2.67之间,在堆肥中期由于堆体温度较高,产甲烷菌活性受到抑制,各组产甲烷菌多样性指数H′明显下降,到堆肥后期,由于堆体温度的下降,各组产甲烷菌多样性指数H′又开始升高。
在整个堆肥过程中,各组产甲烷菌多样性指数H′大小顺序为C<A<B<D<F<E。方差分析表明:各组产甲烷菌多样性指数H′差异显著(P<0.05);E组显著高于A、B、C、D 组(P<0.05),F 组显著高于A、B、C组(P<0.05),其它各组之间差异均不显著(P>0.05)。上述分析表明,高翻堆频率会降低产甲烷菌的多样性,这也可能是高翻堆频率降低甲烷排放总量的主要原因。
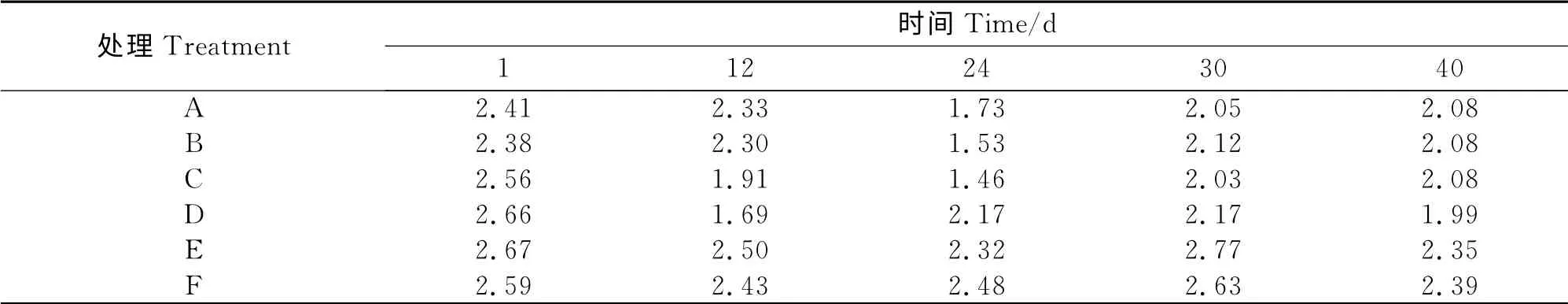
表3 各组产甲烷菌多样性指数H′的变化Table 3 Changes of methanogens diversity index H′
2.5 产甲烷菌总量
各组产甲烷菌拷贝数介于1.77×104~7.02×107copies/g DM 之间(表6)。且各组产甲烷菌拷贝数随堆肥时间的变化规律基本一致,均表现为堆肥初始产甲烷菌含量最高,然后快速下降,在堆肥的6~12d 后,开始缓慢上升。这可能是因为堆肥初始,堆体内的产甲烷菌尚未适应堆体环境,产甲烷菌数量迅速下降,当适应之后,又受堆体温度的升高及堆体可利用碳源和氮源的限制,而缓慢增加。整个堆肥过程中,各组产甲烷菌拷贝数对数值A 组<B组<F组<C 组<D 组<E 组,方差分析表明:差异显著(P<0.05)。A 组显著低于其他各组(P<0.05),B组显著低于C、D、E 组(P<0.05),F 组显著低于E组(P<0.05),其他各组差异不显著(P>0.05)。说明在高翻堆频率下,翻堆可显著降低产甲烷菌的数量;而在低翻堆频率下,翻堆会增加产甲烷菌的数量。这与高翻堆频率会减少CH4的排放,而低翻堆频率会增加CH4的排放的结果相一致。
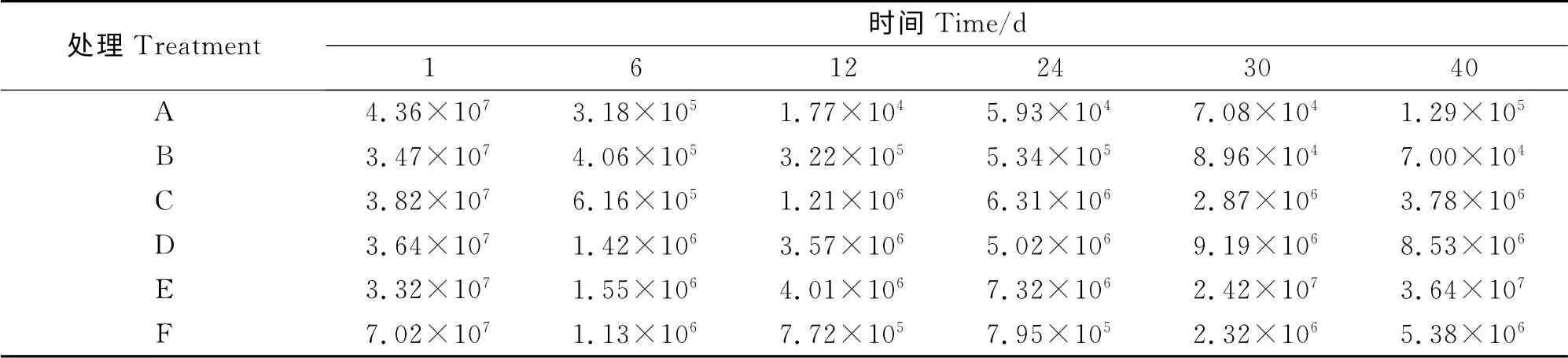
表4 各组产甲烷菌总量Table 4 Total methanogens among different treatment copies/g DM
3 讨论
本研究中,与不翻堆相比,高翻堆频率能减少甲烷的排放量,且随翻堆频率的增加而甲烷排放量降低,低翻堆频率增加了甲烷的排放。这可能有以下两个原因:(1)在高翻堆频率下,堆体内氧气浓度上升较快,堆体内大部分区域氧气浓度较高,抑制了甲烷的生成[5-7]。低翻堆频率下堆体氧气浓度较低,堆体大部分区域处于厌氧状态,适合产甲烷菌活动产生甲烷,而翻堆促进了甲烷的挥发,同时堆体内的甲烷能被甲烷氧化细菌氧化,而翻堆促进了甲烷的挥发,使被氧化的甲烷减少,从而促进甲烷的排放[4]。(2)通过影响产甲烷菌多样性及总量来影响甲烷排放。荧光定量PCR 研究表明,产甲烷菌数量的研究表明高翻堆频率可显著降低产甲烷菌的数量;而低翻堆频率会增加产甲烷菌的数量。这与高翻堆频率会减少CH4的排放,而低翻堆频率会增加CH4的排放的结果相一致。可能是因为翻堆频率越高,前期堆体温越快,越有利于堆体物料降解,且翻堆频率越高高温持续时间越长。而高温期持续时间越长,对产甲烷菌活性的抑制作用也越强,导致高翻堆频率显著降低产甲烷菌的多样性指数和产甲烷菌的总量,从而降低了甲烷的排放。同时高翻堆频率还促进了蛋鸡粪堆肥含水率的降低,而高含水率会降低堆体的通气性能,降低了堆体的烟囱效应,进一步导致了堆体内部氧气缺乏,从而导致低翻堆频率甲烷排放增加。
不同天数翻堆前后1h内收集的气体中,各处理翻堆后甲烷浓度均高于翻堆前,且翻堆频率较低(1次/3d和1次/6d)时,翻堆前后浓度差别最大。说明翻堆会促进甲烷的挥发,翻堆后甲烷排放强度会增大,在翻堆频率较低时,翻堆后甲烷排放强度增加越多。这主要是因为翻堆促进了堆体内部气体的交换,使堆体内的甲烷大量挥发,导致翻堆后甲烷排放强度的短暂增大,而低翻堆频率时,堆体处于厌氧状态区域更大,堆体内蓄积的甲烷更多,因此翻堆后排放强度大幅增加。
4 结论
在蛋鸡粪和锯末堆肥过程中,堆肥初期CH4排放量最大,翻堆对堆肥升温期CH4的排放没有显著影响,但在堆肥高温期和降温期,与不翻堆相比,高翻堆频率(4次/d和2次/d)会减少CH4的排放,而低翻堆频率(1次/3d和1次/6d)会增加CH4的排放。同时,高翻堆频率会显著降低产甲烷菌的多样性指数和产甲烷菌的总量,而低翻堆频率显著增加了产甲烷菌的总量,这可能是导致在不同翻堆频率下,CH4排放量变化的重要原因。
综合不同时期翻堆对CH4排放量、堆肥进程以及成本的影响,2次/d可以降低甲烷的排放,为较合适的翻堆频率。
[1]林而达,李玉娥.全球气候变化和温室气体清单编制方法[M].北京:气象出版社,1998.
[2]Thompson A G,Wagner-Riddle C,Fleming R.Emissions of N2O and CH4during the composting of liquid swine manure[J].Environmental Monitoring and Assessment,2004,91(1/3):87-104.
[3]Tao J,Schuchardt F,Guoxue L.Effect of turning and covering on greenhouse gas and ammonia emissions during the winter composting[J].Transactions of the CSAE,2011,27(10):212-217.
[4]Mosier A,Kroeze C,Nevison C,et al.Closing the global N2O budget:Nitrous oxide emissions through the agricultural nitrogen cycle:OEDC/IPCC/IEA phase II development of IPCC guideline for national greenhouse gas methodology[J].Nutrient Cycling in Agroecosystems,1998,52(2-3):225-248.
[5]Steed J,Hashimoto A G.Methane emissions from typical manure management systems[J].Bioresource Technology,1994,50(2):123-130.
[6]Ahn H K,Mulbry W,White J W,et al.Pile mixing increases greenhouse gas emissions during composting of dairy manure[J].Bioresource Technology,2011,102(3):2 904-2 909.
[7]Park K H,Jeon J H,Jeon K H,et al.Low greenhouse gas emissions during composting of solid swine manure[C]∥Animal Feed Science and Technology Special Issue:Greenhouse Gases in Animal Agriculture-Finding a Balance between Food and Emissions,2011,166-167:550-556.
[8]江 滔,Schuchardt F,李国学.冬季堆肥中翻堆和覆盖对温室气体和氨气排放的影响[J].农业工程学报,2011,27(10):212-217.
[9]Szanto G L,Hamelers H M,Rulkens W H,et al.NH3,N2O and CH4emissions during passively aerated composting of straw-rich pig manure[J].Bioresource Technology,2007,98(14):2 659-2 670.
[10]Hao X,Chang C,Larney F J,et al.Greenhouse gas emis-sions during cattle feedlot manure composting[J].Journal of Environmental Quality,2001,30(2):376-386.
[11]Wright A,Pimm C.Improved strategy for presumptive identification of methanogens using 16Sriboprinting[J].Journal of Microbiological Methods,2003,55(2):337-349.
[12]Zhou M,Hernandez-Sanabria E,Guan L L.Characterization of variation in rumen methanogenic communities under different dietary and host feed efficiency conditions,as determined by PCR-denaturing gradient gel electrophoresis analysis[J].Applied and Environmental Microbiology,2010,76(12):3 776-3 786.
[13]Whelan J A,Russell N B,Whelan M A.A method for the absolute quantification of cDNA using real-time PCR[J].Journal of Immunological Methods,2003,278(1-2):261-269.
[14]Jiang T,Schuchardt F,Li G,et al.Effect of C/N ratio,aeration rate and moisture content on ammonia and greenhouse gas emission during the composting[J].Journal of Environmental Sciences-China,2011,23(10):1 754-1 760.
[15]Shen Y,Ren L,Li G,et al.Influence of aeration on CH4,N2O and NH3emissions during aerobic composting of a chicken manure and high C/N waste mixture[J].Waste Management,2011,31(1):33-38.
猜你喜欢
南京农业大学学报(2023年1期)2023-02-02
安徽农业科学(2022年22期)2022-12-20
中国农业大学学报(2022年3期)2022-05-19
军民两用技术与产品(2021年10期)2021-03-16
水上消防(2020年1期)2020-07-24
智富时代(2019年5期)2019-07-05
智富时代(2019年5期)2019-07-05
疯狂英语·新读写(2018年3期)2018-11-29
黑龙江八一农垦大学学报(2017年5期)2017-10-31
湖南农业科学(2017年7期)2017-03-04
