
Kiss-1/GPR54系统在不同发育阶段五指山猪下丘脑中的表达
2014-01-21 01:16荀文娟侯冠彧周汉林施力光
家畜生态学报 2014年2期
荀文娟,侯冠彧,周汉林,施力光,曹 婷
(中国热带农业科学院热带作物品种资源研究所,海南儋州571737)
Kiss-1基因于1996年被发现,并被鉴定为转移抑制基因,在具有不同转移能力的黑色素瘤细胞中表达存在差异[1]。随后该基因编码的产物kisspeptin被发现,G 蛋白偶联受体54(G protein-coupled receptor 54,GPR54)是kisspeptin 的受体。在人、牛、羊、鼠等哺乳动物研究发现,Kiss-1 基因与GPR54基因一起组成Kiss-1/GPR54系统,参与动物下丘脑-垂体-性腺轴功能的调节[2]。Lents等[3]研究发现,脑室内注射Kisspeptin后备母猪初情日龄提前。Tomikawa等[4]也在初情期母猪下丘脑中检测到Kiss-1基因高水平表达,表明下丘脑Kiss1/GPR54可能是母猪初情启动的关键调节基因。鉴于Kiss-1基因在初情期启动调控中的重要性,本研究推测Kiss-1基因可能与五指山猪高繁殖力和性早熟有关。一直以来,中国地方品种猪的高繁殖能力始终是中外学者的研究热点。五指山猪产于海南省,是我国体型最小、体重最轻的珍稀品种,具有繁殖率高、性成熟早等特性[5]。
本试验以五指山猪作为研究对象,检测Kiss-1/GPR54系统在不同发育阶段五指山猪下丘脑中的表达,从而为阐明动物生殖机理和早熟品种选种选育提供理论基础。
1 材料与方法
1.1 试验动物
选取日龄相近、健康、无生殖缺陷五指山母猪16头,分别于30、60、90(初情期)和120日龄屠宰,采集血清和下丘脑组织样品,血清样品于-20 ℃冰箱保存,组织样品液氮速冻,-70 ℃冰箱保存备用。
1.2 试剂与仪器
RNAiso Plus 总RNA 提取试剂、反转录试剂盒、荧光定量PCR 试剂盒(SYBR R Premix Ex TaqTM)购自Takara;荧光定量PCR 仪购自eppendorf。
1.3 总RNA 的提取及cDNA 第一链合成
采用TaKaRa公司RNAiso Plus试剂盒提取总RNA。用变性琼脂糖凝胶电泳鉴定RNA 的品质,并用核酸蛋白测定仪测定其浓度及纯度。采用SYBR®PrimeScriptTM RT-PCR reagent Kit中的反转录反应试剂进行cDNA 第一条链的合成。
1.4 引物设计
从NCBI下载已登录的猪持家基因序列β-actin(DQ178122)、猪Kiss-1(AB466320)和GPR54(DQ459345)的mRNA 序列,利用Primer Premier 5.0 软件设计合成两对特异性引物,分析Kiss-1和GPR54在不同发育阶段五指山猪下丘脑中的表达水平。引物信息见表1。

表1 荧光定量PCR 所用引物序列Table 1 Primer sequences of quantitative real-time PCR
1.5 Real-time PCR
根据SYBR®Premix Ex TaqTM II试剂盒建议的反应体系及优化确定的反应条件,将设计并合成好的目的基因和看家基因的引物,分别加入反应体系:SYBR Premix Ex Taq II:12.5μL,PCR Forward Primer:1μL,PCR Reverse Primer:1μL,cDNA 溶液2μL,ddH2O:8.5μL,扩增条件:95 ℃预热30s,PCR 循环参数为:变性95.0 ℃5s;退火60.5 ℃/58.5 ℃20s;延伸72 ℃10s,循环40 次。
1.6 血清FSH、LH、E2、P4 的测定
血清FSH、LH、E2、P4用化学发光法测定。
1.7 数据统计
目的基因的相对表达量采用-ΔΔCT 方法获得,ΔCT 就是Kiss-1 和GPR54 相对内参β-actin的表达量,ΔΔCT 为每个试验组的ΔCT 与对照组的相对数值,目的基因的相对表达水平用2-ΔΔCT 表示。所有数据用Excel进行计算,结果用均值±标准差(Means±SD)表示,用SPSS 13.0软件进行方差分析和差异显著性检验,以P<0.05为差异显著水平,P<0.01为差异极显著水平。
2 结果与分析
2.1 Kiss-1mRNA 在不同发育阶段五指山猪下丘脑中的表达
由图1可知,初情期前Kiss-1mRNA 表达量随着五指山猪年龄的增长表达量逐渐升高,初情期(90日龄)时表达量达到最高(P<0.05),到120日龄(初情期后)时表达量逐渐降低(P<0.05)。
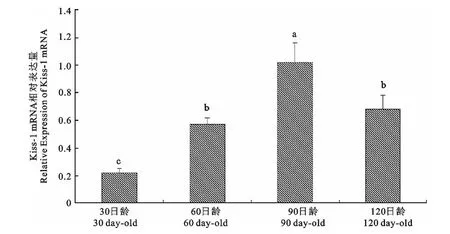
图1 Kiss-1mRNA 在不同发育阶段五指山猪下丘脑中的表达Fig.1 Expression of Kiss-1mRNA in hypothalamus in different age of Wuzhishan pig
2.2 GPR54 mRNA 在不同发育阶段五指山猪下丘脑中的表达
由图2 可知,GPR54 mRNA 表达量随着五指山猪年龄的增长有升高的趋势,但是差异不显著(P>0.05)。

图2 GPR54mRNA 在不同发育阶段五指山猪下丘脑中的表达Fig.2 Expression of GPR54mRNA in hypothalamus in different age of Wuzhishan pig
2.3 五指山猪30~120dFSH、LH、E2、P4 浓度变化
由表2 可知,五指山猪血清FSH、LH、E2、P4浓度变化的趋势大体一致。初情期前,各激素水平逐渐升高,差异不显著(P>0.05),至初情期(90日龄)时显著升高;血清FSH、LH、E2浓度在90日龄和120日龄时差异不显著(P>0.05),P4浓度在90日龄时显著高于120日龄(P<0.05)。

表2 五指山猪30-120日龄之间FSH、LH、E2、P4 浓度变化Table 2 Concentration changes of FSH,LH,E2,and P4of Wuzhishan pig from 30-day-old to 120-day-old
3 讨论
Yasuko等[6]近年来研究发现,Kiss-1 基因参与调控下丘脑-垂体-性腺轴系生殖功能。在青春期,下丘脑中的Kiss-1 mRNA 表达增加,其编码产物Kisspeptin与其受体GPR54结合,通过GnRH 刺激LH 和FSH 的释放[7]。刘小辉等[8]用达那唑皮下注射,以诱导大鼠真性性早熟,表明处于青春期的大鼠下丘脑Kiss-1mRNA 表达量显著高于青春前期。葛伟等[9]在雌性大鼠上也得出了相似结论。可见,Kiss-1mRNA 的表达与生长发育的时期密切相关,提示Kiss-1可能与性早熟的发生有关。本研究中,在初情期前,Kiss-1表达量随着五指山猪年龄的增长呈现不断升高的趋势,初情期(90日龄)时表达量达到最高(P<0.05),随后表达量显著降低(P<0.05),表明Kiss-1对五指山母猪初情期的启动起重要的作用。刘萍[10]对Kiss-1 mRNA 在小梅山猪和苏姜猪下丘脑的表达情况进行了研究,发现小梅山猪和苏姜猪从初生到初情期表达量逐渐上升,初情期后呈下降趋势,与本试验结果基本一致。而Shahab等[11]研究发现,Kiss-1和GPR54mRNA 在雌性猴子下丘脑的表达量随着青春期发育过程逐渐升高,但在去势雄性猴子中则只观察到Kiss-1 mRNA 表达量增加,GPR54mRNA 表达量并未受影响。本研究中,下丘脑GPR54基因的变化不显著,这一结果与去势雄性猴和其他学者关于猪的研究结果相一致[11-12]。GPR54基因变化与其他物种不同可能与其配体对受体上调幅度有关[13]。可见,Kiss-1/GPR54在不同物种及性别个体青春期发育过程中的作用途径还有待深入研究。
自研究者发现Kiss-1/GPR54 系统在生殖功能调控中的重要作用以来,Kisspeptin及其受体对生殖的调控迅速成为研究的热点。性激素对下丘脑Kiss-1表达具有反馈抑制作用。Gottsch等[14]研究发现,给成年鼠注射极少量的Kisspeptin就可以引起其血清中LH 显著增加。而对于幼年小鼠,只有高剂量的Kisspeptin 才能引起LH 含量变化。Brailoiu等[15]研究发现,Kisspeptin神经纤维投射到弓状核、视前区、视交叉后区等,这些区域是调控动物生殖GnRH 分泌的主要神经核团所在区域,由此提示Kisspeptin参与GnRH 分泌的激素调控。刘萍[10]研究发现,Kiss-1mRNA 在各日龄小梅山猪的表达丰度与血浆LH、FSH、E2和P4水平相似并呈显著相关。本试验也得出了类似的结果,五指山猪血清FSH、LH、E2、P4水平在初情期前逐渐升高,至初情期时显著升高,且达到一个峰值,与Kiss-1mRNA 表达丰度基本一致。说明Kiss-1mRNA 水平与初情期前后性激素分泌水平具有相似周期变化规律,证实Kiss-1基因对五指山母猪初情期的启动具有重要的调控作用。
[1]Lee J H,Miele M E,Hicks D J,et al.Kiss-1,a novel human malignant melanoma metastasis-suppressor gene[J].J Natl Cancer Inst,1996,88(23):1 731-1 737.
[2]Navarro V M,CastellanoJ M,Fernandez-Fernandez R,et al.Developmental and hormonally regulated messenger ribonucleic acid expression ofKiss-1and its putative receptor,GPR54,in rat hypothalamus and potent luteinizing hormone-releasing activity ofKiss-1peptide[J].Endocrinology,2004,145(10):4 565-4 574.
[3]Lents C,Heidorn N,Barb C,et al.Central and peripheral administration of kisspeptin activates gonadotropin but not somatotropin secretion in prepubertal gilts[J].Biol Reprod,2008,135(6):879-887.
[4]Tomikawa J,Homma T,Tajima S,et al.Molecular characterization and estrogen regulation of hypothalamic KISS1gene in the pig[J].Biol Reprod,2009,82(2):313-319.
[5]王 峰,姜维思,郑心力,等.五指山猪资源保护现状与建议[J].海南农业科学,2006(2):89.
[6]Yasuko T,Satoshi K,Yoshihiro T,et al.Expression ofKiss-1,a metastasis suppressor gene,in trophoblast giant cells of the rat placenta[J].Biochim Biophys Acta,2004,1678(2-3):102-110.
[7]Navarro V M,Fernandez-Femandez R,Castellano J M,et al.Advanced vaginal opening and precocious activation of the reproductive axis byKiss-1peptide,the endogenous ligand of GPR54[J].J Physiol,2004,561(2):379-386.
[8]刘小辉,刘 方,夏 治,等.Kiss-1在雌性性早熟大鼠下丘脑中mRNA 的表达[J].中国当代儿科杂志,2007,9(1):59-62.
[9]葛 伟,王海莲,薛 江,等.Kiss-1 和GPR54 基因在雌性性早熟大鼠下丘脑中的表达[J].山东大学学报:医学版,2012,50(4):71-75.
[10]刘 萍.Kiss-1基因在母猪初情期下丘脑-垂体-卵巢轴中的定位和发育性变化[D].江苏扬州:扬州大学,2008.
[11]Shahab M,Mastronardi C,Seminara S B,et al.Increased hypothalamic GPR54signaling:apotential mechanism for initiation of puberty in primates[J].Proc Natl Acad Sci USA,2005,102(6):2 129-2 134.
[12]Li S,Ren J,Yang G,et al.Characterization of the porcine Kisspeptins receptor gene and evaluation as candidate for timing of puberty in sows[J].J Anim Breed Genet,2008,125(4):219-227.
[13]朱碧泉,李卓昭,吴 德,等.梅山与长大母猪下丘脑-垂体-卵巢轴Kiss1和GPR54基因的表达差异[J].中国畜牧杂志,2011,47(13):21-25.
[14]Gottsch M L,Cunningham M J,Smith J T,et al.A role for kisspeptins in the regulation of gonadotropin secretion in the mouse[J].Endocrinology,2004,145(9):4 073-4 077.
[15]Brailoiu G C,Dun S L,Ohsawa M,et al.Kiss-1expression and metastin-like immunoreactivity in the rat brain[J].J Comp Neurol,2005,481(3):314-329.
猜你喜欢
作物学报(2022年7期)2022-05-11
猪业科学(2021年3期)2021-05-21
国外畜牧学·猪与禽(2021年6期)2021-01-10
阅读与作文(初中版)(2018年3期)2018-06-08
科学与财富(2017年26期)2017-09-23
国际木业(2016年2期)2016-12-01
兽医导刊(2016年12期)2016-05-17
江苏农业科学(2014年9期)2014-11-15
实验动物与比较医学(2014年4期)2014-02-28
果树学报(2013年2期)2013-05-07
