
利用德氏乳杆菌全酶液水解牛乳酪蛋白
2014-01-18 09:18张咚咚安家彦姜铁民刘继超陈历俊
食品科学 2014年7期
张咚咚,安家彦,姜铁民,刘继超,陈历俊,*
(1.大连工业大学生物工程学院,辽宁 大连 116034;2.北京三元食品股份有限公司,北京 100076)
利用德氏乳杆菌全酶液水解牛乳酪蛋白
张咚咚1,2,安家彦1,姜铁民2,刘继超2,陈历俊1,2,*
(1.大连工业大学生物工程学院,辽宁 大连 116034;2.北京三元食品股份有限公司,北京 100076)
采用响应面法优化超声破碎工艺获取乳酸菌的全酶液,探讨超声破碎功率、超声破碎时间以及溶菌酶的添加量对超声破碎的影响,对获取的全酶液进行水解脱脂乳及酪蛋白的实验。结果表明:随着超声破碎功率的加大,超声破碎时间的增加,所得酶活力值均呈现先增长后下降的趋势,分析所得的回归方程确定最佳工艺修正参数为:超声破碎仪设定功率71%,超声破碎仪设定时间7.5 min,体系中溶菌酶的添加量290 μL/30 mL菌液。在此条件下获取的全酶液水解脱脂乳及酪蛋白显示,超声破碎的过程对酶活力有较大损失,分开处理其胞内和胞外酶液,水解脱脂乳和酪蛋白所获得游离氨基氮的含量分别提高了223.86%和324.11%,高效液相色谱分析结果也证明全酶液水解所得多肽含量及肽段大小多样性均有所提高。
德氏乳杆菌;全酶液;水解;牛乳;酪蛋白
乳酸菌(lactic acid bacteria,LAB)是一大类利用碳水化合物发酵过程中产生乳酸的细菌的总称,广泛存在于人体肠道中,是人体必不可少的益生菌[1],乳酸菌还含有丰富的蛋白水解酶系[2],在应用上的安全性及其所具有复杂的蛋白酶系是其在食品应用上较多的主要原因之一,在乳制品发酵中可产生丰富的风味类物质[3],对水解牛奶中的酪蛋白产生乳清蛋白及多肽[4],促进消化吸收以及潜在的医药应用上,有较大的应用价值。
本实验利用超声破碎菌体获得胞内酶系,并以胞内主要的亮氨酸氨肽酶[4]活力作为响应值做响应面优化工艺研究,而后将胞内和胞外酶系共同作用于12 g/100 mL脱脂乳和1 g/100 mL酪蛋白,检测其蛋白水解能力,并进行高效液相色谱检测。
1 材料与方法
1.1 材料与试剂
1.1.1 菌种
LB2菌株,前期筛选高产蛋白酶乳酸菌所得,保存于北京三元食品股份有限公司技术中心,鉴定为德氏乳杆菌保加利亚亚种(Lactobacillus delbrueckii subsp. bulgaricus)。
1.1.2 试剂与培养基
脱脂乳粉 新西兰西部乳业有限公司;L-亮氨酸-4-硝基苯胺(L-leucine-4-nitroanilide,LNA) 阿法埃莎(天津)化学有限公司;对硝基苯胺 阿拉丁试剂(中国)有限公司;酪蛋白(由牛奶中提取) 北京奥博星生物技术有限责任公司;厌氧盒及厌氧袋 日本三菱瓦斯化学株式会社;溶菌酶 北京索莱宝科技有限公司;其他试剂均为分析纯。
De Man、Rogosa and Sharpe(MRS)培养基和MRS肉汤培养基 北京路桥技术有限责任公司。
1.2 仪器与设备
AC2-4S1二级生物安全柜 美国ESCO公司;Cintra 20双光束紫外-可见分光光度计 澳大利亚GBC公司;3K-15高速冷冻离心机 德国Sigma公司;Polystat K6-1循环式精密恒温水浴锅 德国Huber公司;VCX400超声破碎仪 美国Sonics公司;LC10A vp高效液相色谱日本Shimadzu公司。
1.3 方法
1.3.1 用于超声破碎菌液的制备
将活化好的乳酸菌接入发酵瓶,厌氧37℃培养18~24 h,制成种子液。制备好的种子液按3%的接种量接种于新鲜MRS肉汤培养基中,厌氧37℃培养48 h,考虑到超声破碎本身与培养基的相互影响,所以离心去掉培养基取菌体,用pH 8.0的Tris-HCl缓冲液重悬,洗涤菌体2~3遍[5],重悬于Tris-HCl缓冲溶液中,调整其菌悬液OD600nm为一固定值,4 ℃放置备用[7]。
1.3.2 氨肽酶活力的测定
将超声破碎处理后的菌液稀释数倍,取0.4 mL加入6 mL pH8.0的Tris-HCl缓冲液中,于40 ℃水浴预热5 min,加入0.4 mL 26 mmol/L的LNA乙醇溶液(对照中加超纯水),准确反应30 min后,立即放入冰水浴中,5 min后测定其吸光度A405nm。酶活力定义:在40 ℃、pH 8.0条件下,每分钟水解生成1 μg对硝基苯胺所需要的酶量定义为一个酶活力单位,以U/mL表示[8]。氨肽酶活力按照下式计算。

式中:K为消光系数,本实验中K=0.073 5;6.8为反应的总体积/mL;30为恒温反应时间/min;0.4为酶液体积/mL;D为稀释倍数;ΔA405nm为样品组吸光度与对照组吸光度的差值。
1.3.3 标准曲线的绘制
配制一系列梯度质量浓度的对硝基苯胺标准溶液,以去离子水做对照,分别测定其吸光度A405nm。以吸光度为纵坐标,不同对硝基苯胺质量浓度为横坐标,绘制标准曲线,获得回归方程:y=0.073 5x-0.001 6(R2=0.999 5)。
1.3.4 单因素试验
1.3.4.1 超声功率
取菌液30 mL,设置超声破碎仪的时间为9 min,超声3 s、间隔4 s、超声探头温度4 ℃,分别设定超声功率为50%、60%、70%、80%、90%,由于所用超声破碎仪的总功率为400 W,设定功率分别对应为200、240、280、320、360 W。冰水浴超声。
1.3.4.2 超声时间
取菌液30 mL、超声3 s、间隔4 s、探头温度4 ℃,以1.3.4.1节所得最佳超声功率为设定功率,设定超声破碎仪的超声时间分别为5、7、9、11、13 min,设定超声时间所对应的超声破碎的总时间分别为11.67、16.33、21.00、25.67、30.33 min。冰水浴超声。
1.3.4.3 溶菌酶添加量
取4个50 mL离心管,分别取菌液30 mL,依次加入溶菌酶100、200、300、400 μL,混匀后在37 ℃水浴中反应1 h,反应结束后放入4 ℃冰箱。设定超声3 s、间隔4 s、探头温度4 ℃,以最佳超声功率、最佳超声时间,分别对不同溶菌酶加量的菌液超声破碎,超声过程菌液冰水浴。
1.3.5 响应面优化分析的试验设计
综合单因素试验结果,结合Box-Behnken的中心组合试验设计原理,采用响应面分析法对超声破碎功率、时间和溶菌酶加量做三因素三水平中心点重复5次的试验方案。
1.3.6 利用超声破碎获得全酶液水解脱脂乳和酪蛋白
为避免超声破碎过程中对不需要破碎的发酵液的影响,特别将发酵液与菌体离心分开做对照,一组为直接超声破碎发酵液连带菌体得所需全酶液;另一组将离心所得菌体加入等发酵液体积的pH 7.5磷酸缓冲液中,超声破碎完成后再与发酵液等体积混合,得所需全酶液,在计算水解能力时考虑稀释倍数因素。
取1 mL粗酶液分别加入到2 mL 12 g/100 mL脱脂乳和1 g/100 mL酪蛋白溶液中,37 ℃孵育30 min,加入等体积的24 g/100 mL的三氯乙酸溶液终止反应,室温下静置30 min,3 000×g离心30 min,然后用Whatman 2#滤纸过滤,将滤液保存于-20 ℃冰箱,待用。以先加24 g/100 mL的三氯乙酸和粗酶液37 ℃孵育30 min使酶活力被充分破坏,再加入12 g/100 mL脱脂乳和1 g/100 mL酪蛋白溶液为对照样,作同样条件的处理。定义全酶系水解蛋白的水解程度为100%,以其他各实验组游离氨基氮占全酶系水解组的百分比来表示其蛋白水解能力。
1.3.7 蛋白水解氨基氮的测定
采用邻苯二甲醛法测定水解度[6]。邻苯二甲醛试剂的配制:准确称取0.040 g邻苯二甲醛溶解于1 mL甲醇溶液中,称取0.950 g四硼酸钠(Na2B4O7·10H2O),0.5 g十二烷基磺酸钠,加入100 μL β-巯基乙醇,溶解后定容至50 mL,现配现用。
取100 μL滤液,加入2 mL现配的邻苯二甲醛试剂,室温条件下精确反应2 min,测定340 nm波长处的吸光度。
1.3.8 氨基酸标准曲线的制作
配制1 mg/mL的亮氨酸溶液,依次稀释为50、100、200、300、400、500 μg/mL的标准溶液系列,按照1.3.7节测定吸光度,制作标准曲线,按照式(2)计算样品中的游离氨基氮的含量。

式中:ρ为样品中游离氨基氮的含量/(μg/mL);A为根据滤液的吸光度,通过标准曲线计算出的游离氨基氮的质量浓度/(μg/mL);n为稀释因子。
1.3.9 酶水解液的高效液相色谱检测
对酶水解液过0.22 μm滤膜,按以下条件进行反相色谱分析[7-11]:色谱柱:Agela Venusil ASB C18(250 mm×4.6 mm,孔径150 Å,5 μm粒径);柱温:25 ℃;流速:0.8 mL/min;检测波长:214 nm;进样量:10 μL;梯度混合程序如表1所示。流动相A:0.1%三氟乙酸水溶液,流动相B:0.1%三氟乙酸乙腈溶液。

表1 RP-HPLC 分析的流动相梯度表Table 1 Gradient conditions of the mobile phase for RP-HPLC analysis
2 结果与分析
2.1 单因素试验
2.1.1 超声破碎功率对体系中氨肽酶活力的影响
选取不同超声破碎功率进行破碎,由图1可以看出,当破碎功率设定为70%,即所用功率为280 W时超声破碎效果最好,随着功率的增大,其酶活力急剧下降,这可能与超声过程中产热较快,传导较慢,引起酶结构改变[12],造成酶失活有关,有关文献认为,由于蛋白分子中含有较多的半胱氨酸,超声破碎后产生的自由基会导致蛋白分子间形成二硫键,改变蛋白本身的结构,引起酶结构的改变,从而引起酶活力的下降[13-14]。另外气液界面的剪切效应也是引起酶活力下降的主要因素,Oliva等[15]发现促使蛋白变性的原因主要是蛋白的自身性质和剪切时的气液界面,在超声破碎过程中不可避免的气泡产生,气液面所产生剪切力也是引起蛋白变性及酶失活的原因之一。

图1 超声破碎功率对体系中氨肽酶活力的影响Fig.1 Effect of ultrasonic power on aminopeptidase activity in the fermentation broth
2.1.2 超声破碎时间对体系中氨肽酶活力的影响

图2 超声破碎时间对体系中氨肽酶活力的影响Fig.2 Effect of ultrasonic irradiation time on aminopeptidase activity in the fermentation broth
不同的超声破碎时间下,由图2可以看出,在超声破碎设定时间7 min时获得较高的酶活力,在5 min时可能是超声破碎不完全,而随着超声时间的延长其酶活力在缓慢降低。一般认为在超声破碎过程中,超声波形成的空穴效应及复合伴随形成的瞬时高温高压的变化,都会破坏蛋白结构使酶失活。虽然在实验过程中利用冰水浴使超声破碎样品始终处于较低的温度,但是在空穴气泡破碎的瞬间产生的局部高温高压也可能会引起蛋白结构的破坏和酶的失活[13]。
2.1.3 溶菌酶添加量对体系中氨肽酶活力的影响

图3 溶菌酶添加量对体系中氨肽酶活力的影响Fig.3 Effects of lysozyme dose on aminopeptidase activity in the fermentation broth
溶菌酶是一种专门作用于微生物细胞壁的水解酶,能有效地水解细菌细胞壁的肽聚糖,而不影响细胞的其他组织,且本身无毒无害,是一种天然安全的杀菌剂、防腐剂[14]。
由于本实验采用的德氏乳杆菌保加利亚亚种,有较厚的细胞壁,添加溶菌酶能有效提升细胞的破壁效率。由图3可以看出,在每30 mL菌液中添加量为300 μL时其酶活力较好。
2.2 响应面优化试验
2.2.1 响应面试验设计及其结果分析
根据Box-Behnken的中心组合设计原理,利用Design-Expert.V8.0.6.1软件进行Box-Behnken试验设计,试验方案及结果见表2。

表2 响应面分析试验及结果Table 2 Response surface design matrix and experimental results
2.2.2 拟合模型的建立
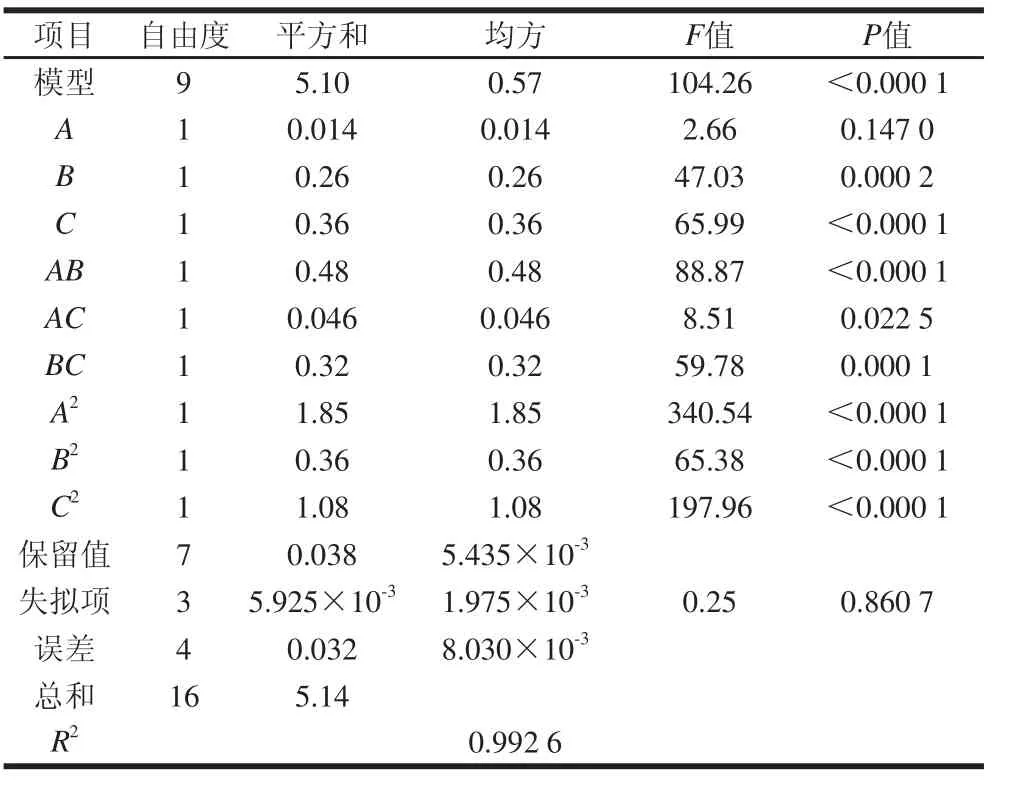
表3 回归方程的方差分析Table 3 Analysis of variance for the regression model
采用Design Expert 8.0.6软件对表2中的实验数据进行多元回归拟合,获得回归方程:R=3.13+0.043A+0.18B-0.21C+0.35AB+0.11AC+0.28BC-0.66A2-0.29B2-0.51C2。
式中采用的是模拟值,R为响应值氨肽酶活力。
从表3可以看出,响应面的回归模型F检验很显著(P<0.05),相关系数R2=0.992 6,表明99%的数据可以依据这个模型来解释,失拟项的F值为0.25,P值为0.860 7>0.05,不显著,说明回归模型失拟不显著,实验误差较小。分析表明可以用回归方程模型代替试验点对实验结果分析。影响酶活力值的因素超声破碎时间(B)和溶菌酶添加量(C)的P值均小于0.01,说明其影响极为显著,超声破碎功率(A)的一次项不显著,说明试验响应值的变化非常复杂,各个试验因素对响应值的影响不是简单的线性关系,而存在二次关系,且三因素间存在明显的交互作用。影响响应值R的因素主次顺序为C>B>A,即是溶菌酶添加量>超声破碎时间>超声破碎功率。
2.2.3 响应面分析
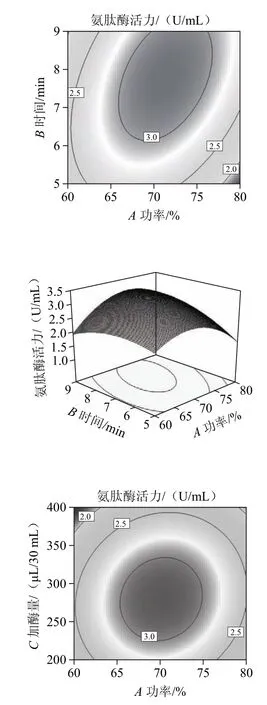

图4 超声破碎功率、时间和溶菌酶添加量对氨肽酶活力影响的响应面和等高线Fig.4 Response surface and contour plots showing the interactive effects of ultrasonic irradiation time, power and lysozyme dose on aminopeptidase activity in the fermentation broth
通过模型方程所得的响应曲面图和等高线图如图4所示。通过Design Expert软件分析所得到的最佳数据组合为:超声破碎仪设定功率71.1%、设定时间7.6 min、溶菌酶添加量289 μL,模型预测的氨肽酶活力值为3.17 U/mL。在实际操作过程中,选取条件为超声破碎仪设定功率为71%、设定时间为7.5 min,溶菌酶添加量为290 μL/30 mL菌液,做3次平行实验所得酶活力值的平均值为3.13 U/mL,与理论预测值基本相符,说明该回归方程较为合理。
2.2.4 酶液水解12 g/100 mL脱脂乳和1 g/100 mL酪蛋白的氨基氮含量
根据亮氨酸溶液制作的氨基酸标准曲线获得回归曲线方程为y=0.002 2x+0.022 9,R2为0.998 9,相关较强。如图5、6所示,在超声破碎所得胞内酶系的共同作用下其水解底物能力的显著增强,脱脂乳和酪蛋白体系的游离氨基氮质量浓度分别由651.41、454.14 μg/mL增加到2 109.70、1 926.05 μg/mL;在发酵液连带菌体的情况下直接进行细胞的超声破碎,其水解底物的能力仅有部分的增强,体系游离氨基氮的含量仅分别增加到858.69 μg/mL和785.56 μg/mL。一方面胞内蛋白水解酶系的释放,会增加总的蛋白水解能力,但是由于超声破碎过程中,对原来释放到发酵液中的胞外蛋白水解酶系的破坏,使其蛋白水解能力有部分抵消,不能显著增强其蛋白水解能力。在将发酵液中释放的胞外蛋白酶系低温离心外部存放后,不会对其造成过多的破坏,然后再将菌体重悬于低温磷酸缓冲液中,进行超声破碎获取胞内的蛋白水解酶系,这样的方式,不但能够最大限度的保留胞外蛋白酶系,也能最大限度地获取胞内的蛋白水解酶系。另一方面,由于乳酸菌的发酵产酸特性,发酵液中的pH值呈酸性,可能会对胞内蛋白酶系产生部分破坏,加入的磷酸缓冲液会减弱此破坏作用,使大部分蛋白水解酶发挥较大的水解能力。在超声破碎过程中,缓冲液的黏度要比发酵液的黏度小,同等超声破碎条件下,可能会有更充分的超声破碎效果[17]。

图5 粗酶液与12 g/100 mL脱脂乳反应体系中游离氨基氮的含量及水解度Fig.5 Free amino nitrogen content and degree of hydrolysis in the reaction system of 12 g/100 mL skim milk and crude enzymes from Lactobacillus delbrueckii subsp. bulgaricus

图6 粗酶液与1 g/100 mL酪蛋白反应体系中游离氨基氮的含量及相对水解度Fig.6 Free amino nitrogen content and degree of hydrolysis in the reaction system of crude enzymes from Lactobacillus delbrueckii subsp. bulgaricus and 1 g/100 mL casein
2.2.5 酶水解液的高效液相色谱检测
经过高效液相色谱后,水解液中的多个多肽组分被较为清晰地分开,在实验中并没有具体确定各多肽的具体成分,仅希望通过分析能较为直观的证明乳酸菌的全酶在水解酪蛋白后产生的多肽更具有多样性,为之后进一步研究其营养价值提供依据[18-19]。

图7 全酶液与胞外酶液水解12 g/100 mL脱脂乳的高效液相色谱图Fig.7 Comparative chromatograms of 12 g/100 mL skim milk hydrolysates by holoenzyme and extracellular enzyme
如图7所示,全酶液水解色谱图中各峰面积较胞外酶液水解组有一定的增加,尤其是27.82 min处峰值增加较为明显,增幅42.63%;41.78 min处峰面积增幅达216.95%,说明此处多肽含量有明显的增加;在42.76 min左右的两个色谱峰变化更为明显,左侧峰的明显加大及右侧峰的明显降低,说明多肽成分及含量的显著变化,说明在不同酶系水解过程中的相似性及差异性。
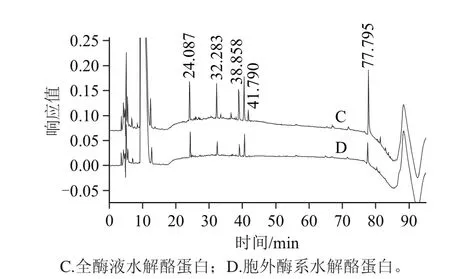
图8 全酶液与胞外酶液水解1 g/100 mL酪蛋白的高效液相色谱图Fig.8 Comparative chromatograms of 1 g/100 mL casein hydrolysates by holoenzyme and extracellular enzyme
图8 是全酶液和胞外酶液水解1 g/100 mL酪蛋白的水解液色谱分析图,全酶液组各色谱峰面积的显著增大,尤其是38.86 min处的峰面积增加了24.32%,77.80 min处的峰面积更是增加了233.83%,表明各多肽含量的增加明显;色谱峰数也有较多的增加,25~29 min出现较多小峰,34、37 min左右、42 min,66~68 min,以及后面洗脱峰的较多出现,都表明有较多新的肽段的出现,表明全酶的水解过程中酶系水解的复杂性及多样性。多肽是目前被研究较多的物质,不但在促进消化吸收上面,而且在其他的一些药用上面有较广泛的价值,全酶的水解应用,不但直接促进蛋白的消化吸收,还有潜在的药用医用价值。
3 结 论
在对乳酸菌全酶系的应用上,本研究采用超声破碎技术,通过响应面优化超声破碎获得胞内酶系。确定体系的最佳超声破碎参数组合为超声破碎仪设定功率为71%、设定时间为7.5 min,溶菌酶添加量为290 μL/30mL菌液,获得最大亮氨酸氨肽酶活力值为3.13U/mL,将此条件下所获得胞内酶系和胞外酶系结合应用到水解脱脂乳及纯酪蛋白上。
在全酶系水解酪蛋白的应用上,尽量排除超声破碎过程中对酶活力的影响,采用菌体菌液分开超声破碎而后再合并的方法,对水解效果有较大的提升,脱脂乳水解过程中体系中水解产生的游离氨基氮含量从仅胞外酶的651.41 μg/mL提升到分开超声破碎时的2 109.70 μg/mL,提升223.86%;牛奶中酪蛋白水解体系中从454.14 μg/mL提升到1 926.05 μg/mL,提升324.11%,水解能力显著增加,这可能和乳酸菌内含有复杂的蛋白水解酶系有关[20]。高效液相色谱分析也表明,全酶系的应用不仅仅是在多肽含量上有增加,而且对多肽大小的多样性上也有较多增加,尤其是在酪蛋白的水解上。这些对促进牛奶中大分子酪蛋白的消化吸收[21],以及对多肽对人体生理方面的影响及其他方面进一步研究都有较大的参考价值。
[1] FULLER R. Probiotics in man and animals[J]. Journal of Applied Bacteriology, 1989, 66(5): 365-378.
[2] LAW B A, KOLSTAD J. Proteolytic systems in lactic acid bacteria[J]. Antonie van Leeuwenhoek, 1983, 49(3): 225-245.
[3] SAVIJOKI K, PALVA A. Purification and molecular characterization of a tripeptidase (PepT) from Lactobacillus helveticus[J]. Applied and Environmental Microbiology, 2000, 66(2): 794-800.
[4] 卞春. 牛乳酪蛋白改性婴儿配方乳的研究[D]. 大庆: 黑龙江八一农垦大学, 2006.
[5] 隋欣, 任发政, 鲁言文, 等. 热激法修饰瑞士乳杆菌的参数优化与特性研究[J]. 农业机械学报, 2009, 40(8): 111-114.
[6] 刘丰, 陈历俊, 隋欣, 等. 契达干酪浆中使用热激瑞士乳杆菌的研究[J].中国食品学报, 2012, 12(8): 87-92.
[7] 李永霞, 秦礼康. 细菌胞壁破碎条件优化及高肽酶菌株的筛选[J].食品与发酵工业, 2012, 38(4): 89-93.
[8] 吴庆勋, 古海先, 赵光鳖. N+注入诱变选育氨肽酶高产菌株及发酵条件初步优化[J]. 食品与发酵工业, 2006, 32(1): 15-18.
[9] HUANG X W, CHEN L J, LUO Y B, et al. Purification, characterization, and milk coagulating properties of ginger proteases[J]. Journal of Dairy Science, 2011, 94(5): 2259-2269.
[10] HAYALOGLU A A, GUVEN M, FOX P F. Influence of starters on chemical, biochemical, and sensory changes in Turkish white-brined cheese during ripening[J]. Journal of Dairy Science, 2005, 88(10): 3460-3474.
[11] BONIZZI I, BUFFONI J N, FELIGINI M. Quantification of bovine casein fractions by direct chromatographic analysis of milk. Approaching the application to a real production context[J]. Journal of Chromatography A, 2009, 1216(1/2): 165-168.
[12] 刘斌, 马海乐, 李树君, 等. 超声破处理对脱脂麦胚分离蛋白结构的变化研究[J]. 光谱学与光谱分析, 2011, 31(8): 2220-2225.
[13] 罗昭锋, 瞿鑫, 沐万孟, 等. 超声和高压处理对牛血清白蛋白结构的影响[J]. 中国生物工程杂志, 2006, 26(1): 46-49.
[14] ARAKAWA T, KITA Y. Protection of bovine serum albumin from aggregation by tween 80[J]. Journal of Pharmaceutical Sciences, 2000, 89(5): 646-651.
[15] OLIVA A, SANTOVEÑA A, FARIÑA J, et al. Effect of high shear rate on stability of proteins: kinetic study[J]. Journal of Pharmaceutical and Biomedical Analysis, 2003, 33(2): 145-155.
[16] 徐敬宜. 溶菌酶及其在食品工业中的应用[J]. 食品与机械, 2006(1): 90-92.
[17] 薛娟琴, 吴川眉. 超声波对溶液性质的影响[J]. 金属世界, 2008(1): 25-28.
[18] KAMINARIDES S E, ANIFANTAKIS E M. Comparative study of the separation of casein from bovine, ovine and caprine milks using HPLC[J]. Journal of Dairy Research, 1993, 60(4): 495-504.
[19] 张源淑, 陈伟华, 邵思湘. 用HPLC测定酪蛋白酶解产物中的β-casomorphin-7[J]. 中国乳品工业, 1999, 27(1): 37-38.
[20] KUNJI R S, MIERAU I, HAGTING A, et al. The proteotytic systems of lactic acid bacteria[J]. Antonie van Leeuwenhoek, 1996, 70(2/4): 187-221.
[21] ARMAFORTE E, CURRAN E, HUPPERTZ T, et al. Proteins and proteolysis in pre-term and term human milk and possible implications for infant formulae[J]. International Dairy Journal, 2010, 20(10): 715-723.
Hydrolysis of Milk Casein with Lactobacillus delbrueckii subsp. bulgaricus Holoenzyme
ZHANG Dong-dong1,2, AN Jia-yan1, JIANG Tie-min2, LIU Ji-chao2, CHEN Li-jun1,2,*
(1. School of Biological Engineering, Dalian Polytechnic University, Dalian 116034, China; 2. Beijing Sanyuan Foods Co. Ltd., Beijing 100076, China)
Response surface methodology (RSM) was employed to optimize ultrasonic power, irradiation time and lysozyme dose for the release of holoenzyme from Lactobacillus delbrueckii subsp. bulgaricus. Experiments were conducted to investigate the hydrolysis of skim milk and casein with the holoenzyme. Results indicated that the enzyme activity rose at first and then declined with increasing ultrasonic power and extended irradiation time. From the proposed regression model, the optimal conditions for holoenzyme release were determined as ultrasonication for 7.5 min at a power of 71% and hydrolysis with lysozyme at a dose of 290 μL/30 mL of cell suspension. The whole fermentation broth of Lactobacillus delbrueckii subsp. bulgaricus was treated under these conditions to obtain holoenzyme 1 or centrifugally separated into cells and supernatant. The cells were individually treated under the same conditions and combined with the supernatant to obtain holoenzyme 2. The content of free amino nitrogen in hydrolyates of skim milk and casein by holoenzyme 2 was increased by 223.86% and 324.11%, respectively, when compared with holoenzyme 1. This may be attributed to losses of extracellular enzyme activities caused by ultasonication. HPLC analysis showed that polypeptides with diverse molecular weight distribution were generated in higher amounts by using the whole enzymes than extracellular enzymes alone.
Lactobacillus delbrueckii subsp. bulgaricus; holoenzyme; hydrolysis; milk; casein
TS252.1
A
1002-6630(2014)07-0107-06
10.7506/spkx1002-6630-201407022
2013-04-19
“十二五”国家科技支撑计划项目(2013BAD18B04)
张咚咚(1988—),男,硕士研究生,研究方向为食品微生物及其应用。E-mail:medong@126.com
*通信作者:陈历俊(1967—),男,高级工程师,博士,研究方向为乳品加工。E-mail:chlj@263.net
猜你喜欢
中国饲料(2022年5期)2022-04-26
成都大学学报(自然科学版)(2021年1期)2021-05-22
乳业科学与技术(2019年6期)2019-12-22
中国乳品工业(2017年9期)2017-10-12
中国酿造(2016年12期)2016-03-01
Asian Agricultural Research(2015年3期)2015-02-02
食品工业科技(2014年3期)2014-03-22
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
中国兽药杂志(2012年4期)2012-11-06
郑州大学学报(理学版)(2012年4期)2012-03-25
