
低温胁迫下玉米苗期根系保护酶活性及内源激素变化
2014-01-16 00:44魏湜罗宁李晶顾万荣陈光寿李丽杰谢腾龙朱恒光
东北农业大学学报 2014年9期
魏湜,罗宁,李晶,顾万荣,陈光寿,李丽杰,谢腾龙,朱恒光
(1.东北农业大学农学院,哈尔滨 150030;2.农业部东北地区作物栽培科学观测实验站,哈尔滨 150030;3.越南农业部栽培局,越南河内 100803)
低温胁迫下玉米苗期根系保护酶活性及内源激素变化
魏湜1,2,罗宁1,2,李晶1,2,顾万荣1,2,陈光寿1,3,李丽杰1,谢腾龙1,朱恒光1
(1.东北农业大学农学院,哈尔滨 150030;2.农业部东北地区作物栽培科学观测实验站,哈尔滨 150030;3.越南农业部栽培局,越南河内 100803)
为揭示不同抗冷性玉米品种幼苗根系保护酶活性及内源激素与品种抗寒性的关系,以耐低温型(郑单958),中间型(先玉335),低温敏感型(丰禾1)玉米品种为试验材料,分析幼苗根系在14℃/5℃、16℃/7℃、18℃/9℃、20℃/11℃(CK)4个温度梯度,处理2、4、6、8、10 d时根系保护酶SOD、POD活性、MDA含量及内源激素ABA、IAA、GA变化。结果表明,随着胁迫时间延长,不同温度处理下SOD、POD活性及MDA含量相对对照均呈上升趋势,SOD、POD活性表现为郑单958>先玉335>丰禾1,MDA含量变化为丰禾1>先玉335>郑单958。低温胁迫下ABA显著高于对照,IAA变化不显著,GA减少。表明品种间对低温存在抗(耐)性差异,幼苗在低温胁迫下根系生长受到抑制,但耐低温型品种自身调节能力较强,通过内源激素调节可保持根系正常生长,其中ABA在缓解低温胁迫伤害中起关键作用。
玉米;低温胁迫;幼苗根系;保护酶;内源激素
玉米是黑龙江省第一大粮食作物,种植范围遍及全省,生长期内易发生延迟型低温冷害,且以苗期常见。生产上土壤表层温度稳定在10℃以上,发芽温度为6~7℃,5℃时根系几乎停止生长[1]。苗期根系生长状况与活跃度直接影响玉米生长发育与产量构成。根系是激素类物质合成场所,是其生长活跃的代谢器官,低温土壤环境影响根系对水分与矿物质的吸收,导致生长受阻[2]。
逆境胁迫下植物代谢产生具有较强氧化能力自由基,破坏生物功能分子。Avinash研究指出在逆境胁迫下,细胞膜保护酶系统对清除活性氧,维持细胞膜的稳定起重要作用[3]。随着胁迫增强,抗氧化酶活性也显著增强[4]。对逆境胁迫下水稻、辣椒等作物叶片研究较多[5-6],但对玉米根系保护酶活性却鲜有报道。
植物体内内源激素在植物根系内已被发现,并可作为一些信号分子参与低温胁迫的调控[7]。前人就玉米种子萌发[8],不同生长期叶片[9-13]内源激素的研究较多,但对低温胁迫下玉米幼苗根系内植物内源激素却鲜有报道。植物内源激素虽含量甚微,但在植物生长代谢与作物产量形成和品质保持中起重要作用。
基于对黑龙江省常规种植60个品种低温胁迫下芽期及苗期反应试验及品种低温反应聚类分析结果[14]。选择生产上大面积种植的温度敏感度不同的三个类型品种为试材,于三叶期进行低温胁迫处理,测定根系保护酶活性及根系激素浓度变化,探讨分析不同冷敏感型品种在低温下保护酶系统与激素水平上差异表现,明确低温胁迫对玉米根系生理伤害机理,以及为生产上筛选抗寒品种、提高品种抗冷水平提供理论依据。
1 材料与方法
1.1 供试材料
供试玉米材料为郑单958(耐低温型)、先玉335(中间型)、丰禾1号(低温敏感型)。
供试土壤为黑土,来自东北农业大学香坊农场实验实习基地。基本理化性质为:有机质为31.7 mg·kg-1,碱解氮(N)217.8 mg·kg-1,速效磷(P2O5)49.8 mg·kg-1,速效钾(K2O)231.1 mg·kg-1。
1.2 处理方法
1.2.1 幼苗培养
试验种子于75%乙醇中浸泡15 s,放入2%次氯酸钠溶液消毒10 min,经蒸馏水冲洗3遍,于培养钵中播种育苗,培养钵直径20 cm,高17 cm,装土3 kg,3次重复,保留长势一致的植株,每个培养钵保苗定植6株。
1.2.2 低温胁迫处理
玉米于室外正常环境下长至三叶一心时,将幼苗移入人工智能培养箱(HPG-280HX)中进行低温处理,光照时长为12 h/12 h(昼/夜),相对湿度为65%。设(昼/夜)14℃/5℃、16℃/7℃、18℃/ 9℃、20℃/11℃(CK)4个温度梯度,分别处理2、4、6、8、10 d。培养期间严格控制温度及含水量,采用称重法补充土壤水分。
1.3 取样与测量
取样时,轻轻敲打培养钵里幼苗的浮土,避免损坏根系。放入500 mL大烧杯的水中,用毛笔慢慢扫掉根系上的土粒和杂质。每个指标的测定对于样品部位具有严格的一致性,确保根系受到的损伤最小。然后用蒸馏水润洗根系,滤纸吸干。取样后的根系于液氮中冷冻后迅速保存于-80℃超低温冰箱中,以供保护酶活性与内源激素含量的测定。
1.3.1 POD、SOD、MDA测定
SOD活性测定采用Gisnnopolitis等[15]与郝再彬[16]NBT光照化学还原法,POD活性测定采用愈创木酚法[16],MDA含量测定采用硫代巴比妥酸显色法[17]。
1.3.2 内源激素测定
每个样品准确称取0.5 g,用酶联免疫吸附法(ELISA)测定样品中各激素含量,3次重复。酶联免疫试剂盒由中国农业大学农学与生物技术学院提供。
1.4 数据分析
各处理指标数据分析采用DPS 7.05进行方差差异分析和Excel 2010进行作图统计。
2 结果与分析
2.1 低温胁迫下幼苗根系保护酶变化
2.1.1 幼苗根系SOD活性变化
SOD以清除植物体内积累过多的O2-,减轻逆境对其的伤害。由图1可知,经低温胁迫后各处理SOD活性上升。耐低温型品种SOD上升幅度大于敏感型品种,且耐低温能力越强,上升幅度越大。在20℃/11℃(CK)温度下,供试品种SOD均处于较低水平,活性变化不大。随温度降低,SOD活性增加趋势缓慢。郑单958、先玉335、丰禾1于14℃/5℃低温处理下10 d相对低温处理2 d分别提高47.8%、28.1%、21.5%。耐低温型增加幅度高于敏感型品种,利于植株抵御低温伤害。低温胁迫初期,郑单958在14℃/5℃下相对对照分别提高6.8、5.4、4.4倍,而丰禾1在14℃/5℃下相对对照分别提高4.5、4.3、3.9倍。18℃/9℃低温处理下郑单958、丰禾1 SOD差异不明显。这可能是18℃/9℃的低温尚不能引起幼苗根系细胞内活性氧代谢平衡的紊乱与启动膜的过氧化所致。
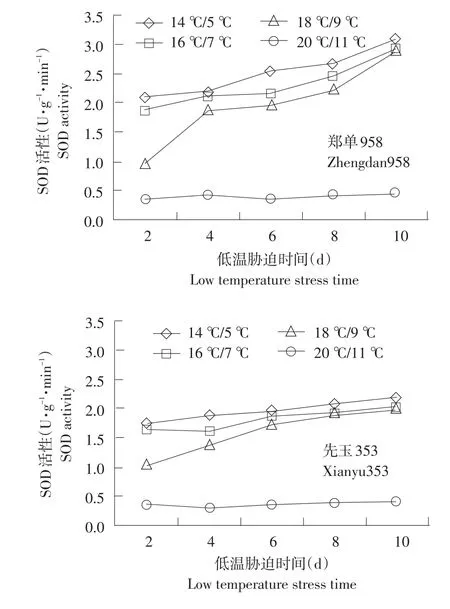
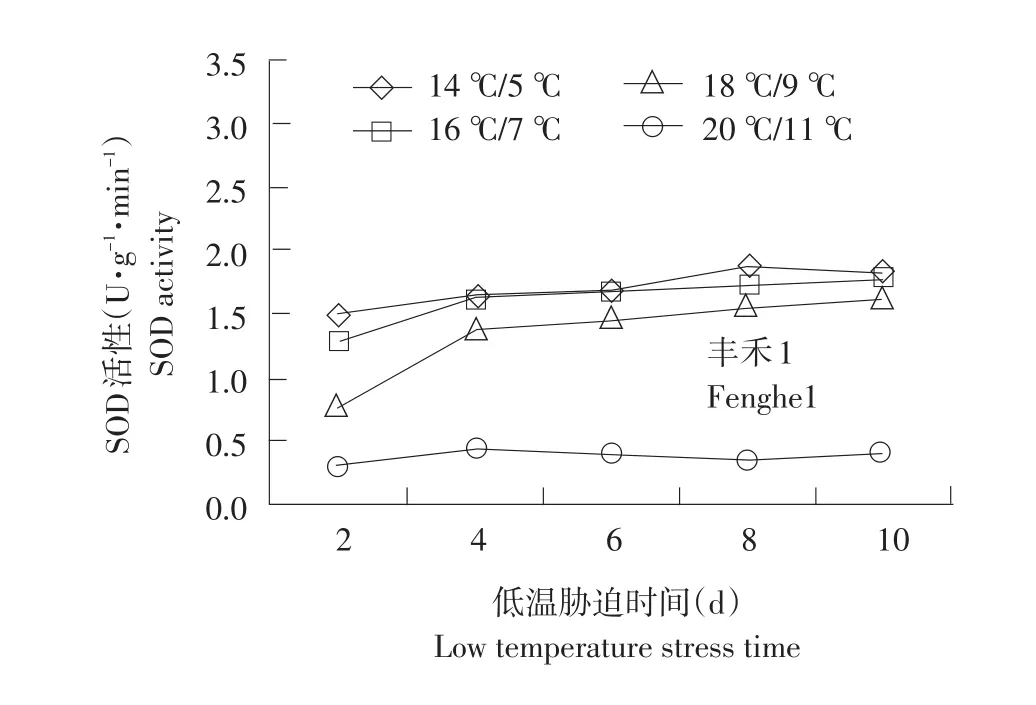
图1 不同温度下不同品种幼苗根系SOD活性变化Fig.1 Changes of SOD activity under different temperature of different seedling roots
2.1.2 幼苗根系POD活性变化
不同温度下不同品种幼苗根系POD活性变化见图2。由图2可知,经低温处理后,郑单958、先玉335根系POD含量与对照相比,呈上升趋势,而丰禾1幼苗根部POD活性经低温处理,第2天先下降,后上升。整个低温处理期间,3个供试品种幼苗根部POD活性(除丰禾1低温处理10 d)均高于对照。其中郑单958、先玉335在14℃/5℃、16℃/ 7℃、18℃/9℃时显著高于对照,丰禾1在14℃/ 5℃时显著高于对照,在18℃/9℃时差异不显著。随胁迫时间延长,不同品种POD活性增幅表现出差异,郑单958与先玉335 14℃/5℃低温处理10 d相对2 d,POD活性增幅分别为69.3%和45.9%。丰禾1 14℃/5℃时低温处理6 d相对2 d时,增幅为9.2%。可能由于低温胁迫下,低温敏感型细胞膜受到自由基的损坏,代谢紊乱。同时表明,POD活性可以反映玉米幼苗的抗寒能力,耐低温型比低温敏感型玉米POD活性强,上升幅度大。
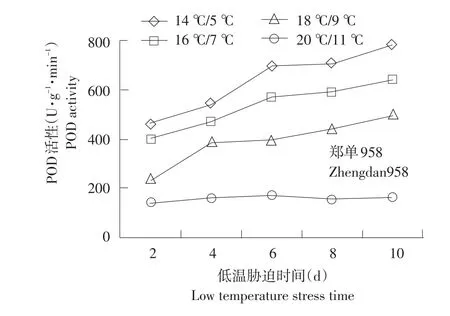

图2 不同温度下不同品种幼苗根系POD活性变化Fig.2 Changes of POD activity under different temperature of different seedling roots
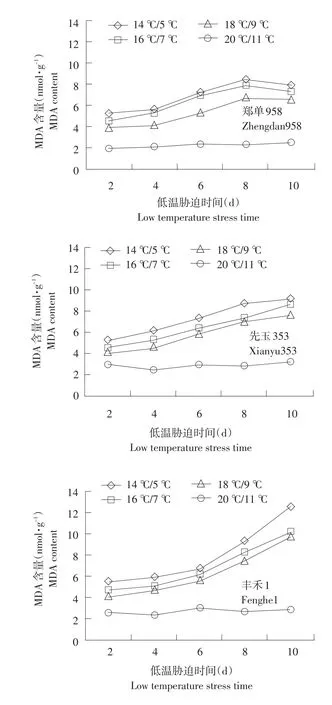
图3 不同温度下不同品种MDA含量变化Fig.3 Changes of MDA content under different temperature of different seedling roots
2.1.3 幼苗根系MDA活性变化
不同温度下不同品种MDA含量变化见图3。
由图3可知,供试品种经低温处理后,幼苗根系MDA发生不同变化,郑单958低温处理2 d时14℃/5℃、16℃/7℃、18℃/9℃3个温度胁迫下,差异不显著。随着胁迫时间延长,呈缓慢递增趋势,呈单峰曲线变化,峰值出现在第8天,第8天时较低温处理2 d MDA含量分别增加71.4%、61.3%、50%。先玉335、丰禾1中MDA变化趋势与郑单958趋势基本一致,但未出现峰值。丰禾1在14℃/5℃、16℃/7℃、18℃/9℃温度下与对照相比差异显著。表明,随着低温胁迫程度加深,根系细胞脂膜过氧化程度加深。不同胁迫温度下,MDA均出现增长,且增长率为丰禾1>先玉335>郑单958。表明耐低温型品种通过自身调节可以减少植物细胞MDA生成,增加其抗寒性。
图3 不同温度下不同品种MDA含量变化Fig.3 Changes of MDA content under different temperature of different seedling roots
2.2 低温胁迫下幼苗根系内源激素含量变化
2.2.1 幼苗根系ABA含量变化
不同温度下不同品种幼苗根系ABA含量变化见图4。低温胁迫下,根系幼苗ABA含量变化随处理温度的降低与时间延长呈升高趋势。耐低温型品种幼苗根系中ABA含量显著(P<0.05,n=3)大于低温敏感型。

图4 不同温度下不同品种幼苗根系ABA含量变化Fig.4 Changes of ABA content under different temperature of different seedling roots
由图4可知,郑单958在14℃/5℃下,ABA含量相对16℃/7℃、18℃/7℃、20℃/11℃(CK)较高,在第2天时比其他温度分别提高1.8、2.98、3.11倍。随胁迫时间延长,第4、6、8、10天相对第2天增长率为2.0%、9.2%、11.6%、55.4%。丰禾1体内ABA含量在14℃/5℃低温胁迫2 d有所增加,但增加不明显,相对16℃/7℃、18℃/7℃、20℃/ 11℃(CK)分别提高3.0%、12.9%、28.5%。随着胁迫时间延长,相对4、6、8、10 d增长率为2.8%、8.1%、12.3%、23.9%。先玉335增长率介于其他两品种之间。随胁迫时间延长与胁迫程度加深,幼苗根系ABA含量表现为郑单958>先玉335>丰禾1。耐低温型品种对低温胁迫反应较明显,其含量在短时间内迅速上升,抵御外界不良环境,这可能是其耐低温的原因之一。
2.2.2 幼苗根系IAA含量变化
不同温度下不同品种幼苗根系IAA变化见图5。
由图5可知,整个处理期间IAA含量基本呈上升趋势,幅度变化较小。耐低温品种在不同温度处理下差异显著(P<0.05,n=3)。各品种变化有所不同,郑单958随胁迫时间延长处于缓慢增长趋势,先玉335在低温处理第6天有明显上升,之后保持平稳,甚至有所下降。丰禾1在第8天有明显上升趋势,之后保持平稳。可能是由于低温胁迫可降低植物体内IAA氧化酶含量。在一般情况下,IAA氧化酶活性越低,IAA含量越高,对植物生长抑制越小。
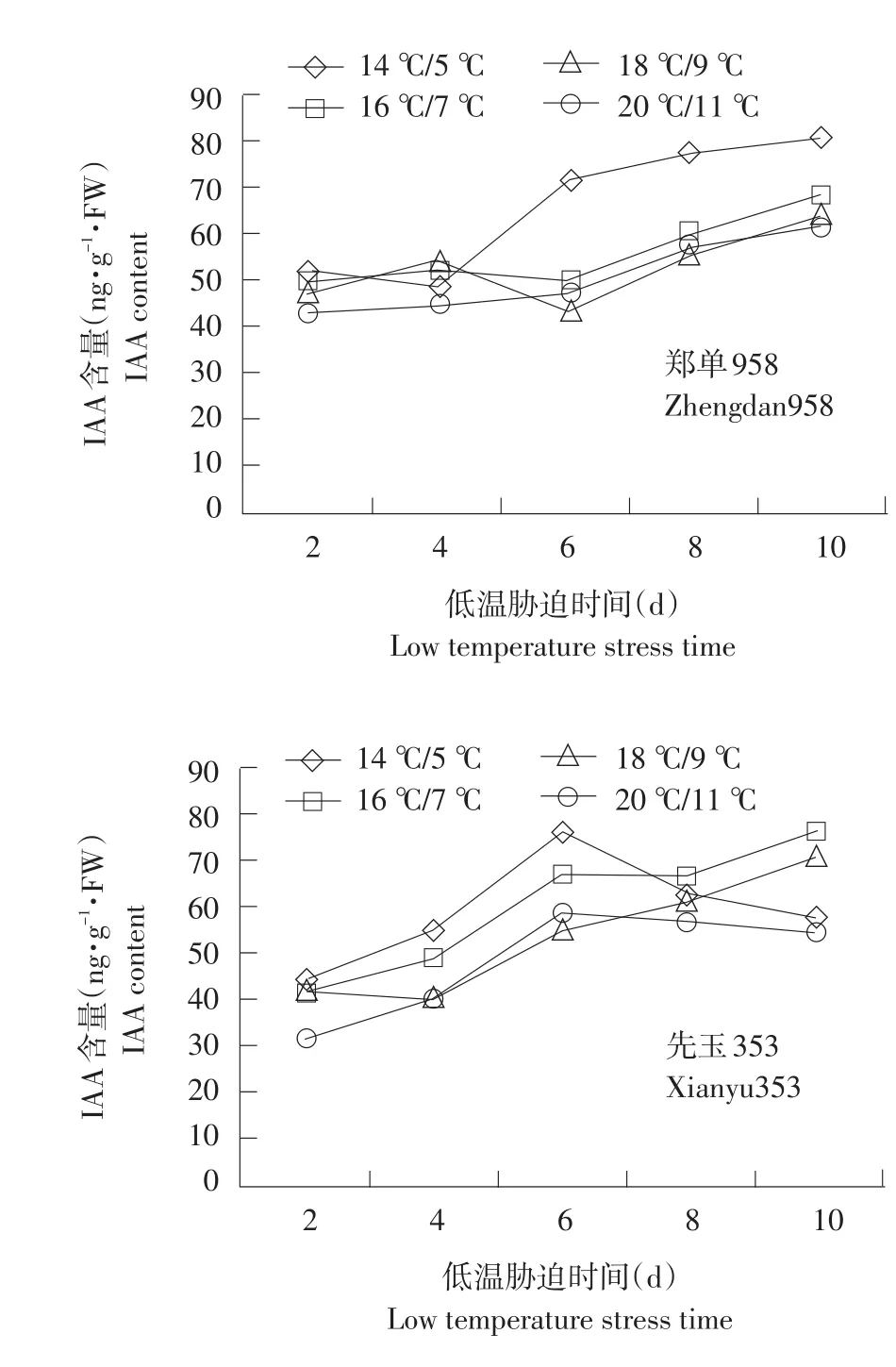
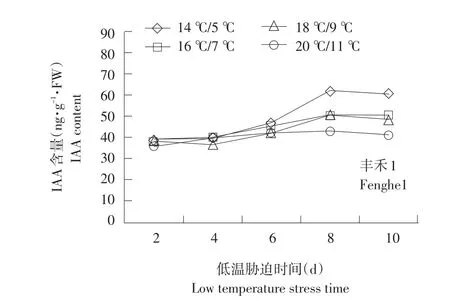
图5 不同温度下不同品种幼苗根系IAA变化Fig.5 Changes of IAA content under different temperature of different seedling roots
2.2.3 低温胁迫下根系幼苗GA含量变化
不同温度下不同品种根系幼苗GA含量变化见图6。GA含量变化较为复杂,耐低温型郑单958根系GA含量随着胁迫时间延长相比对照含量降低。GA敏感度强,在胁迫初期立即下降。
由图6可知,胁迫第2天郑单958根系GA含量14℃/5℃相对16℃/7℃、18℃/9℃、20℃/11℃分别降低52.1%、56.3%、62.3%。先玉335与丰禾1在18℃/9℃、20℃/11℃温度处理下差异不显著。不同温度处理下丰禾1 GA含量较高。并随着胁迫温度降低,保持下降趋势,但是下降保持平缓。幼苗增长随胁迫时间延长程度加深,含量下降程度为郑单958>先玉335>丰禾1。可能是由于耐低温品种GA维持在较低水平,可迅速解除低温胁迫对根系造成的伤害,以恢复正常代谢。
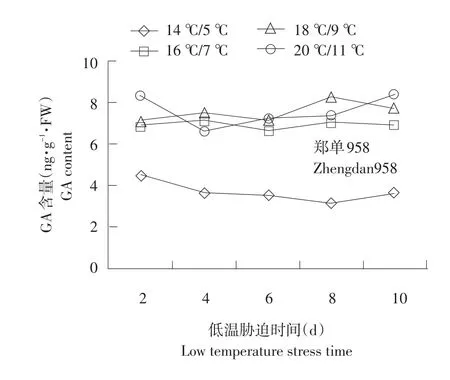
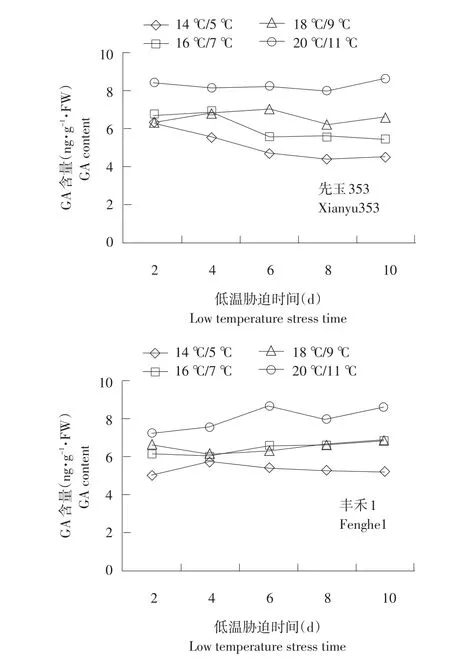
图6 不同温度下不同品种根系幼苗GA含量变化Fig.6 Changes of GA content under different temperature of different seedling roots
3 讨论
植物在遭受逆境胁迫时,不仅影响植株生长,体内也发生生理生化变化。活性氧平衡失调将导致体内自由基大量积累,引发脂膜过氧化作用,造成细胞膜系统损伤。苗期玉米根系的生长,决定土壤中水分与营养物质的吸收与对逆境抵抗能力[18]。因此,苗期根系MDA含量、SOD、POD活性、ABA、IAA、GA内源激素含量与植物抗寒性有密切关系。
3.1 低温胁迫对玉米根系膜系统的影响
植株在逆境胁迫下,自由基增多,SOD、POD等植物体内的酶清除系统被激发,其活性上升,以清除过多自由基[19]。在一定范围内,保护酶系统的酶活性与自由基保持一定平衡,保证植株正常生长。李春燕等研究表明,低温胁迫初期小麦叶片中SOD、POD活性均呈上升趋势[20]。孙富等研究表明,在低温条件下,甘蔗幼苗叶片透性增大,叶绿体中MDA含量增加,叶片中SOD、POD活性增大[21]。本试验研究结果与以上研究规律一致,随着胁迫温度降低,供试品种SOD、POD活性均上升。不同冷敏感品种表现不同,郑单958 SOD、POD活性均显著高于丰禾1,可能是由于耐低温植物一般具有较高的膜脂不饱和度,在较低温度下保持流动性,维持正常生理功能,因而其膜脂相变温度低于不抗冷植物[22]。
低温胁迫下,细胞膜系统遭到破坏,质膜过氧化程度加深,MDA作为过氧化重要产物之一,可以衡量低温胁迫下质膜的过氧化程度。本试验研究表明,随着胁迫时间延长与程度加深,3个品种MDA含量均呈上升趋势,证明低温造成质膜过氧化。玉米根系保护酶活性上升趋势均表现为郑单958>先玉335>丰禾1,MDA含量丰禾1>先玉335>郑单958。说明耐冷性玉米品种可以清除活性氧,抗氧化酶的活性高,清除能力强,对脂膜系统的伤害较少,可以保证各类代谢物质的适应性变化,因此抗寒性较强。
3.2 低温胁迫对幼苗根系内源激素的影响
植物激素在植物体内代谢产生,并能运输到各个部位进行信息交流,微量浓度就有明显调节生长发育效应[23]。目前广泛研究与应用的有ABA、GA、IAA,在根系中都有发现[24],逆境胁迫下其含量影响植物代谢与发育。本试验研究表明,低温胁迫下玉米幼苗根系内源激素水平发生显著变化,随胁迫时间延长,ABA含量升高,郑单958显著高于先玉335与丰禾1。IAA含量略微升高,随胁迫时间延长其含量平缓上升。GA含量下降。这与周碧燕[25],王兴等[26]研究结果基本一致。三种内源激素均对玉米根系低温胁迫作出应答反应,从反应时间与含量变化分析,ABA变化较大,各处理与对照相比均达显著差异,且耐低温品种间均为显著差异。这可能是由于ABA促进气孔关闭,水分散失减少,保证体内水分平衡,减少逆境伤害[27],是ABA促进抗寒基因表达[28]。
沈漫等有研究表明高含量ABA可提高SOD、POD保护酶活性,减少超氧自由基含量,缓解逆境伤害[22]。GA含量下降,保持根系较慢的根系细胞分裂速度与生长速度,减轻低温胁迫对植物危害,缓解生理活动压力,对抗不利环境,使植物在逆境下免受更大伤害。在低温胁迫下,耐低温型郑单958 ABA含量高于敏感型丰禾1,而GA含量前者低于后者,表现出相反趋势。是由于ABA/ GA可反映出抗冷性的关系,随着ABA/GA降低,抗冷性减弱[28]。植物抗寒机制同样也是几种激素互作效应。ABA在胁迫初期含量升高,变化幅度大,因此在抗寒机制中起关键作用。综上所述,低温胁迫下,抗寒性不同的三个品种变化幅度不同。植物内源激素ABA通过激发抗寒基因,诱导保护酶活性增加,ABA、GA、IAA含量变化缓解逆境对植物伤害。
4 结论
玉米品种在耐低温特性上存在明显差异,从幼苗根系的保护酶系统及内源激素上,耐低温品种都具有明显优势,温度越低,优势越明显。低温胁迫使幼苗根系保护酶系统,内源激素含量发生变化。表明根系细胞产生过多自由基,破坏细胞膜透性。通过ABA含量增多诱导抗寒基因,ABA、IAA、GA互作效应保持植物代谢平衡。本试验20℃/11℃(CK)、18℃/9℃、16℃/7℃、14℃/5℃温度处理间差异显著。综合来看,耐低温型品种郑单958保护酶活性与内源激素含量均表现较强优势,低温下表现较稳定的质膜系统,有效缓解逆境伤害的激素含量,为提高低温下品种抗冷能力提供保障。
[1]魏湜.黑龙江玉米生态生理与栽培[M].北京:中国农业出版社,2013.
[2]Andreas H,Yvan F.Cold tolerance of maize seedlings as deter⁃mined by root morphology and photosynthetic traits[J].Europ J Agronomy,2008,28:178-185.
[3]Avinash C R,Major S,Kavita S.Effect of water withdrawal on for⁃mation of free radical,proline accumulation and activities of anti⁃oxidant enzymes in ZAT12-transformed transgenic tomato plants [J].Plant Physiology and Biochemistry,2012,61:108-114.
[4]郝福顺,崔香环,赵世领,等.渗透胁迫对黑麦幼苗活性氧和抗氧化酶活性的影响[J].植物学通报,2007(5):603-608.
[5]Shvaleva A L,Silva F C,Breia E,et al.Metabolic responses to wa⁃ter deficit in two Eucalyptus globulus clones with contrasting drought sensitivity[J].Tree Physiology,2006(2):239-248.
[6]Kavita S,Sareeta N.Heat exposure alters the expression of SOD, POD,APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars[J].Plant Physi⁃ology and Biochemistry,2012,57:106-113.
[7]Kefeli V I,Kalevitch M V.Natural growth inhibitors and phytohor⁃mones in plants and environment[M].London:Kluwer Academic Publishers,2003:7-9.
[8]谢阳姣,吕凤连,戴罗杰,等.种子包衣对微胚乳玉米种子发芽过程中生理生化变化的影响[J].玉米科学,2009(6):53-55,59.
[9]顾万荣,李召虎,张明才,等.DCPTA和DTA-6对大豆和玉米苗期叶片内源激素与氧自由基代谢的影响[J].植物遗传资源学报,2009(2):300-305.
[10]王如芳,张吉旺,赵斌,等.不同类型玉米品种分蘖发生过程中内源激素的作用[J].中国农业科学,2012(5):840-847.
[11]王艳芳,崔震海,张立军,等.不同类型春玉米灌浆期间籽粒中内源激素IAA、GA、ZR、ABA含量的变化[J].植物生理学通讯,2006(2):225-228.
[12]李青苗,杨文钰,关华,等.烯效唑浸种对玉米幼苗生长和内源激素含量的影响[J].植物生理学通讯,2005(6):752-754.
[13]汤继华,赫忠友,李永亮,等.玉米温敏核雄性不育育性转换与内源激素的关系[J].作物学报,2003(3):336-338.
[14]杨猛.低温胁迫对玉米幼苗的生理生化响应及差异蛋白研究[D].哈尔滨:东北农业大学,2011.
[15]Gisnnopolitis C N,Nries S K.Superoxide dismutases occurrence in higher plants[J].Plant Physiol,1977,59:309-314.
[16]郝再彬.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社, 2004:115-116.
[17]李合生.植物生理生化试验指导[M].北京:高等教育出版社, 2000:164-167.
[18]曹宁,符力,张玉斌,等.低温对玉米苗期根系生长及磷养分吸收的影响[J].玉米科学,2008,16(4):58-60.
[19]Yoshiaki U,Naoko U,Haruto S,et al.Impacts of acute ozone stress on superoxide dismutase(SOD)expression and reactive oxy⁃gen species(ROS)formation in rice leaves[J].Plant Physiology and Biochemistry,2013,70:396-402.
[20]李春燕,陈思思,封超年,等.苗期低温胁迫对扬麦16叶片抗氧化酶和渗透调节物质的影响[J].作物学报,2011,37(12): 2293-2298.
[21]孙富,杨丽涛,谢晓娜,等.低温胁迫对不同抗寒性甘蔗品种幼苗叶绿体生理代谢的影响[J].作物学报,2012(4):732-739.
[22]沈漫,王明庥,黄敏仁.植物抗寒机理研究进展[J].植物学通报,1997(2):1-8.
[23]符继红,孙晓红,王吉德,等.植物激素定量分析方法研究进展[J].科学通报,2010(33):3163-3176.
[24]段留声,田晓莉.作物化学控制原理与技术[M].北京:中国农业大学出版社,2011:130-132.
[25]周碧燕,李宇彬,胡志群,等.低温胁迫和喷施ABA对荔枝内源激素和成花的影响[J].园艺学报,2002,29(6):577-578.
[26]王兴,苍晶,李卓夫,等.低温条件下不同抗寒性冬小麦内源激素的变化[J].麦类作物学报,2009,29(5):827-831.
[27]李静.拟南芥IAM路径合成的IAA及其与逆境胁迫响应[D].哈尔滨:哈尔滨师范大学,2012.
[28]李霞,戴传超,程睿,等.不同生育期水稻耐冷性的鉴定及耐冷性差异的生理机制[J].作物学报,2006,32(1):76-83.
Change of the root protective enzyme activities and endogenous hormones of maize seedling under low-temperature stress
WEI Shi1,2,LUO Ning1,2,LI Jing1,2,GU Wanrong1,2,CHEN Guangshou1,3,LI Lijie1,XIE Tenglong1,ZHU Hengguang1(1. School of Agriculture,Northeast Agricultural University,Harbin 150030,China;2.Observation Experiment Station of the Ministry of Agriculture for Crop Cultivation Science in Northeast Area,Harbin 150030,China;3.Bureau ofAgriculture Cultivation,Hanoi 100803,Vietnam)
In order to reveal the protective enzyme activity,endogenous hormones in the seedling root and their relationships to low temperature stress,Zhengdan958(cold resistance type),Xianyu335(middle type),Fenghe1(sensitivity type)were investigated at diurnal temperature of 14℃/5℃,16℃/7℃,18℃/ 9℃,20℃/11℃(CK)for 2,4,6,8,10 d,which analyzed the SOD and POD activities,MDA content and the change of ABA,IAA,GA.The results showed that SOD and POD activities were Zhengdan958>Xianyu335>Fenghe1,MDA content showed Fenghe1>Xianyu335>Zhengdan958.The results showed that with a prolonged stress under different temperature treatment,the activity of SOD,POD and the content of MDA in relative comparison were on the rise.The content of IAA changed insignificantly,the content of GA was decreased,which indicated that cold resistance type had higher ability to remove free radical and resulting toreduce the damage of membrane and keep plant metabolic.As low temperature inhibited the growth of root system,endogenous hormone adjustment could keep it grow normally,which showed that resistant varieties had strong resist-ability to low temperature and ABA played a key role in relieving stress damage.
maize;low-temperature stress;seedling root;protective enzyme;endogenous hormones
S513
A
1005-9369(2014)09-0001-08
2014-05-16
国家科技支撑计划(2011BAD32B03);国家自然科学基金(31401320);寒地作物品种改良与生理生态重点开放实验室项目(HDZW-008)
魏湜(1956-),男,教授,博士,博士生导师,研究方向为作物高产栽培生理生态、宏观农业。E-mail:weishi5608@163.com
时间2014-9-18 10:43:32[URL]http://www.cnki.net/kcms/detail/23.1391.S.20140918.1043.006.html
魏湜,罗宁,李晶,等.低温胁迫下玉米苗期根系保护酶活性及内源激素变化[J].东北农业大学学报,2014,45(9):1-8.
Wei Shi,Luo Ning,Li Jing,et al.Change of the root protective enzyme activities and endogenous hormones of maize seedling under low-temperature stress[J].Journal of Northeast Agricultural University,2014,45(9):1-8.(in Chinese with English abstract)
猜你喜欢
现代园艺(2022年7期)2022-11-19
四川劳动保障(2021年3期)2021-06-09
江苏农业科学(2019年2期)2019-08-13
现代园艺(2017年13期)2018-01-19
动物营养学报(2016年7期)2016-08-03
中国农业文摘-农业工程(2016年5期)2016-04-12
电线电缆(2016年5期)2016-02-27
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
橡胶工业(2015年5期)2015-08-29
橡胶工业(2015年10期)2015-08-01
