
黄土高原半干旱区天然草地群落结构与土壤水分关系
2014-01-02 08:12张志南武高林王冬邓蕾郝红敏杨政上官周平
草业学报 2014年6期
张志南,武高林,王冬,邓蕾,郝红敏,杨政,上官周平,
(1.西北农林科技大学农学院,陕西 杨凌712100;2.西北农林科技大学水土保持研究所,陕西 杨凌712100;3.中国科学院水利部水土保持研究所 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌712100)
*植物群落的结构格局是环境中各种物理、化学和生物因素综合影响以及组成物种对上述因素综合反映的结果[1-3]。水分是影响生态系统和植物群落的重要因素[4],尤其是在干旱、半干旱地区水分是制约植物生存和生长发育的关键因素,对植被状况和生态恢复有很大的限制作用[5-7]。近年来,气候变化对生态系统的影响已成为生态学、植物学和地理学的研究热点问题[8-10]。气候变暖导致蒸发加剧,尤其是对于干旱半干旱区,在降水增加不明显的条件下,加速了土壤干燥化程度。草原是自然环境严苛、生态系统稳定性相对脆弱的典型区域之一[11-12],预测水分波动对草地群落的影响对维持生态系统的稳定具有重要意义。目前很多学者对于土壤水分与植被群落生产力、多样性及二者的关系进行了大量研究,包括羊草(Leymuschinensis)草地植物功能群组成及多样性特征对水淹干扰的响应[13]、土壤水分与生产力的关系[14-16]、水分梯度上放牧对群落多样性与生产力的影响[17]、土壤水分对根系垂直分布的影响[18-20]等,但这些研究往往集中于干扰(水淹干扰、放牧干扰)程度对群落多样性及生产力的影响,或者是土壤水分的垂直变化对植被生产力的影响,而对于黄土高原区域土壤水分空间异质性对群落演替的影响尚未见报道。
黄土高原是世界上水土流失最严重和生态环境最脆弱的地区之一,因此植被恢复是黄土高原遏制水土流失,促进生态系统恢复的关键因素和有效途径[12]。然而,土壤水分是制约黄土高原地区植被恢复与重建的主要限制因子[5,21-22]。因此,本研究以黄土高原半干旱区天然草地为对象,研究天然草地群落在土壤水分梯度条件下的变化特点,探讨草地植被随土壤水分变化的演替规律,对于黄土高原草地生态系统的适应性管理具有重要意义。
1 材料与方法
1.1 研究区域概况
研究区位于甘肃省中北部黄土丘陵沟壑草原地段,地理位置为103°24′11.94″~104°54′13.92″E,35°15′9.12″~37°22′9.60″N,海拔1452~2523m,由于受季风的影响,雨季、旱季差异明显,年降雨量大部分为185~350mm。雨季集中在7-9月,此期降水量多达全年降水量的50%~80%,而从10月至来年6月为旱季。年平均气温在7~10℃之间。土壤以棕钙土、栗钙土为主。由于自然因素制约和人为活动的影响,天然草地受到不同程度的破坏。植被类型主要为长芒草(Stipabungeana)、冰草(Agropyroncristatum)、糙隐子草(Cleistongenessquarrosa)、骆驼蓬(Peganummultisectumz)、猪毛菜(Salsolacollina)、油蒿(Artemisiaordosica)、铁杆蒿(Artemisia gmelinii)、冷蒿(Artemisiafrigida)、茭蒿(Artemisiagiraldii)、百里香(Potentillabifurca)、红砂(Reaumuria soongorica)等。
1.2 研究方法
1.2.1 调查测定方法 野外调查工作于2011年8月草地群落地上生物量高峰期进行。为了揭示气候、土壤等自然因素与草地生物量的关系,尽量选择围栏封育或远离公路受人为干扰较轻的天然草地群落。采用随机取样的方法,对研究区域内的地带性植被群落进行随机调查。每个典型群落设置1块样地,样地面积不少于1hm2;在各选定的样地中心布设1个10m×100m的样带,然后沿直线方向每隔10m布设1个1m×1m观测样方,每个样地共调查10个样方,并按顺序编号,调查样方内所有草本植物的总盖度和群落高度。在偶数样方内,分物种调查植物平均高度、株数和地上部活体生物量;在奇数样方内,调查群落的地上、地下生物量、凋落物质量和土壤水分。
1.2.2 样品处理与室内分析 地上生物量调查方法:将样方内植物地面以上的所有绿色部分用剪刀齐地面剪下,不分物种按样方分别装进信封袋,做好标记后带回室内,在65℃下烘干后称量干重。数据记录时保留小数点后2位。样品量较多时,先称量总鲜重,然后取部分鲜样品,称量鲜重后进行烘干、测定,所得值除以其取样比率,即可获得整体干重值。
凋落物生物量调查方法:将地表当年的凋落物和立枯收集,按样方分别装入信封内做好标记后带回室内,在65℃下烘干后称量干重。数据记录时保留小数点后2位。样品量较多时,方法同地上活体生物量的调查方法。
地下生物量调查方法:因为草地根系主要分布在0~1m土层内,1m以下土层内根系极少,基本无根系分布,所以本研究把0~1m土层内的根系生物量作为草地总地下生物量。取样前将样方土壤表面的残留物和杂质清理干净,然后在5个取过地上生物量的样方内,用直径为9cm的根钻采集地下0~1m土层内的根系生物量,按照0~20~40~60~80~100cm进行分层,每个样方沿对角线方向取3~5钻,混合后,用干筛法获取根系。取好的样品,按样方分装在尼龙袋纱袋中,并用塑料标签写好样方号和各样方的根钻数,置于孔径为0.4mm纱袋内带回室内漂洗,然后装进信封并做好标记,放进65℃烘箱烘至恒重后称量干重。
土壤水分的调查方法:在挑选根系后的土壤中用铝盒取土壤样品,土层与根系层对应。分层取样,每层5个重复,做好标记后带回实验室,称取土壤鲜重后放入105℃烘箱烘干称重。
1.2.3 植物功能群的划分及计算 根据草地群落内植物所处的科的不同将植物划分成3类,禾本科:即禾本科植物;豆科:即所有豆科植物;杂类草:除去禾本科和豆科以外的其他科的植物统称为杂类草。禾本科所占比例为禾本科质量占地上生物量的百分比;杂类草所占比例为杂类草质量占地上生物量的百分比;豆科所占比例为豆科质量占地上生物量的百分比。
1.2.4 群落多样性特征分析 物种多样性采用Shannon-Winner指数来计算:
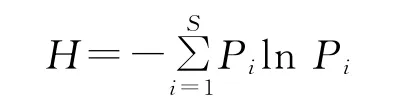
式中,H为物种多样性指数;Pi为属于种i的个体在全部个体中的比例;S为物种数目。物种丰富度的计算:
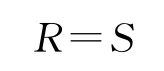
式中,S为物种数目。
1.3 数据分析
地上生物量为样地调查时1m2样方内的所有地上生物量,对于地下生物量,把各样方的根系生物量换算成单位面积的地下生物量,在此基础上取平均值,得到每个样地的地下生物量;凋落物为烘干后质量;总生物量=地上生物量+地下生物量+凋落物质量。土壤鲜重与土壤干重的差与土壤干重的比值乘以100%,为土壤含水量[15]。
土壤含水量与生物量、功能群比例和多样性关系以及多样性与生物量的关系进行拟合分析。在SPSS 17.0软件上进行相关统计处理,采用Sigma Plot 10.0进行图形处理。
2 结果与分析
2.1 研究区群落和土壤概况
方差分析结果表明,各样地土壤含水量差异显著(F=33.59,P<0.0001)。在生产力水平上,草地的地上生物量(F=13.03,P<0.0001)、地下生物量(F=27.56,P<0.0001)、总生物量(F=27.58,P<0.0001)和凋落物质量(F=23.47,P<0.0001)存在显著的差异(表1)。在物种多样性水平上,各样地间群落的物种多样性(F=20.67,P<0.0001)和丰富度(F=45.23,P<0.0001)有显著差异。在群落的功能群组成上,各样地群落的禾本科(F=16.62,P<0.0001)、杂类草(F=17.89,P<0.0001)、豆科(F=3.68,P<0.0001)差异显著(表1)。
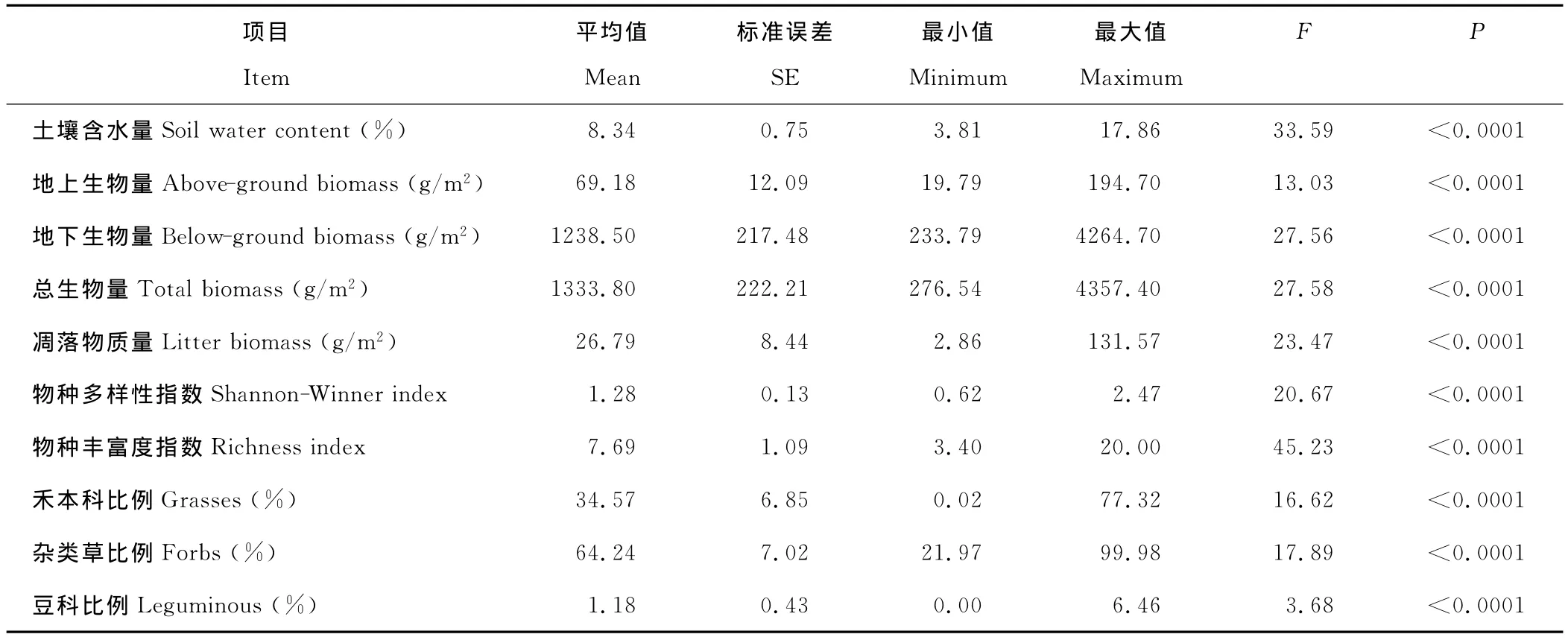
表1 天然草地群落随土壤含水量的变化特点(n=18)Table 1 Change of plant community characteristics along with soil water in the nature grasslands(n=18)
2.2 群落生物量随土壤含水量的变化
从整体趋势来看,随土壤含水量的增加地上生物量显著增加(相关系数R=0.64,P=0.0044)(图1);地下生物量与土壤含水量呈线性增加趋势,但是不显著(相关系数R=0.33,P=0.40),总生物量的变化趋势与地下生物量的变化趋势一致,随着土壤含水量的增加表现先增加后减小的趋势(相关系数R=0.36,P=0.35);凋落物质量随土壤含水量的增加而增加,但不显著(R=0.24,P=0.34)。
2.3 群落物种多样性和功能群组成随土壤含水量的变化
由图2看出,物种多样性和丰富度在水分梯度上的变化格局极为一致,都表现为随土壤水分增加而增加的趋势,且物种丰富度与土壤水分的相关性显著(R=0.72)。
本试验结果显示,土壤含水量在9.27%处,禾本科所占群落比例最大。土壤含水量在7.72%处,杂类草所占比例最大。豆科植物相对于禾本科和杂类草在群落中所占的比例较小,随土壤水分含量增加豆科植物在群落中比例从0到6.46%(图3)。禾本科在群落中所占比例随土壤含水量的增加呈显著增加趋势(R=0.49,P=0.04);与此相反的是杂类草随土壤含水量的增加呈显著减少的趋势(R=-0.51,P=0.029);豆科植物随着土壤含水量的增加呈显著的线性增加趋势(R=0.62,P=0.0059)。

图1 天然草地土壤含水量与生物量的关系Fig.1 The relationships between soil water content and biomass in the nature grassland

图2 天然草地土壤含水量与群落物种多样性的关系Fig.2 The relationships between soil water content and community diversities in the nature grassland
3 讨论
水是限制植物生长发育的主要因子之一,特别是在气候干旱半干旱、生态环境脆弱的黄土高原地区,植物群落特征与分布对水分需要十分敏感[21,23]。本研究结果表明,在土壤水分梯度条件下,地上生物量与土壤水分呈正相关关系,此结果与高寒草地群落的研究结果一致[4]。而且在牧压梯度上对群落多样性的研究与本结果也一致[24]。相对于地上生物量随土壤水分变化规律而言,地下生物量和总生物量与地上生物量表现出不一致性。地下生物量随着土壤含水量的增加而增加但不显著,由于地下生物量占总生物量的90%以上,所以总生物量在水分梯度上也表现同样趋势。地下生物量与土壤含水量相关不显著的原因是,土壤水分减少,群落对资源的竞争加强,尤其是在半干旱区植物对水资源的竞争更加激烈,当水分减小到植物本身的生理耐受性时,物种多样性减少(图2),抗旱能力强的物种存活下来,为获取更多水资源根系向深层土壤延伸[20]。因此,植物根系随土壤含水量的减少不明显。由于前人的研究也只是针对地上植被随水分梯度的变化[4,17],或者是垂直水分梯度上地下生物量的变化[15,25],而对于水平梯度上水分与地下生物量的关系鲜有报道。因此此研究结果为以后研究半干旱区土壤水分空间异质性与地下生物量关系提供参考。
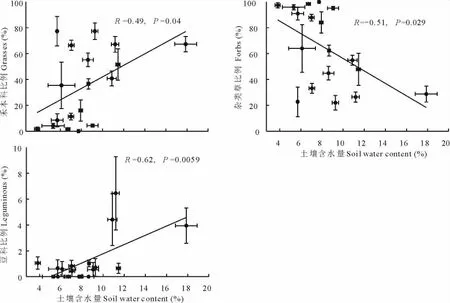
图3 天然草地土壤水分与群落中各功能的关系Fig.3 The relationships between soil water content and functional groups in the nature grassland
水分梯度上群落的物种多样性决定于次生优势种和优势种间的竞争和土壤水分状况[4]。在一定的土壤含水量范围,土壤含水量较高时,优势物种占主导地位,抑制了大多数植物的生长发育。随着土壤含水量的减少改变了群落种间关系,消弱了优势种对种群的作用,群落具较高的生物多样性。王正文等[26]的研究结果表明,群落的物种丰富度、多样性在经历中等强度水淹干扰的样带上较高,而在未受干扰的对照样带和受干扰较为严重的样带都较低,符合“中度干扰”假说,这与物种受水淹干扰后的生理耐受性和物种间的互作有关。本研究结果表明,物种多样性和丰富度与土壤含水量呈正相关关系。以上2个结果与本研究结果存在矛盾,其原因可能是:1)研究地域不同,导致水热条件不同,植物的生态类型也不尽相同。高寒草甸以湿生、湿中生植物占主要优势,当面对环境干扰后,其优势度降低给外来物种增加入侵机会。而本研究的地区是黄土高原干旱半干旱区,旱生、旱中生植物占主要优势,当土壤含水量减少时,物种间的竞争及植物自身抵御干旱的能力不同,优胜劣汰使竞争力强的物种存活下来。2)自然因素和人为控制实验之间存在差异,自然条件下存在不可控因素。而本研究正是自然条件下,水分状况对群落多样性的影响。王正文等[26]研究的是人为控制的水分梯度对群落多样性的影响,因此与自然条件下半干旱区水分条件对群落多样性的影响不具可比性。
本研究显示,在土壤水分含量较多时,禾本科和豆科植物占主要优势,尤其是豆科植物与土壤含水量的相关性最大(图3),在土壤水分含量较低时,杂类草则占主要优势。因此,土壤水分不仅在垂直水平上影响植物生物量及其分布[15,27],而且在水平梯度上影响植物群落的组成(图3)。在黄土高原区,土壤水分不仅影响植物的空间分布格局,而且植物的功能群组成作为指使物种来反映土壤水分状况。
[1]Mitchell J E,West N E,Miller R W.Soil physical properties in relation to plant community patterns in the shadscale zone of northwestern Utah[J].Ecology,1966,47(4):627-630.
[2]黄德青,于兰,张耀生,等.祁连山北坡天然草地地下生物量及其与环境因子的关系[J].草业学报,2011,20(5):1-10.
[3]王长庭,龙瑞军,王根绪,等.高寒草甸群落地表植被特征与土壤理化性状,土壤微生物之间的相关性研究[J].草业学报,2010,19(6):25-34.
[4]Wu G L,Ren G H,Wang D,etal.Above-and below-ground response to soil water change in an alpine wetland ecosystem on the Qinghai-Tibetan Plateau,China[J].Journal of Hydrology,2012,476:120-127.
[5]苏嫄,焦菊英,王巧利.黄土丘陵沟壑区不同侵蚀环境下幼苗库及其与地上植被的关系[J].草业学报,2013,22(5):154-164.
[6]杨阳,韩国栋,李元恒,等.内蒙古不同草原类型土壤呼吸对放牧强度及水热因子的响应[J].草业学报,2012,21(6):8-14.
[7]张娜,梁一民.黄土丘陵区天然草地地下/地上生物量的研究[J].草业学报,2002,11(2):72-78.
[8]Kardol P,Cregger M A,Campany C E,etal.Soil ecosystem functioning under climate change:plant species and community effects[J].Ecology,2010,91(3):767-781.
[9]刘国华,傅伯杰.全球气候变化对森林生态系统的影响[J].自然资源学报,2001,16(1):71-78.
[10]任继周,梁天刚,林慧龙,等.草地对全球气候变化的响应及其碳汇潜势研究[J].草业学报,2011,20(2):1-22.
[11]李愈哲,樊江文,张良侠,等.不同土地利用方式对典型温性草原群落物种组成和多样性以及生产力的影响[J].草业学报,2013,22(1):1-9.
[12]郑粉莉.子午岭林区植被破坏与恢复对土壤演变的影响[J].水土保持通报,1996,16(5):41-44.
[13]白永飞,李凌浩,王其兵.锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J].植物生态学报,2000,24(6):667-673.
[14]黄德青,于兰,张耀生,等.祁连山北坡天然草地地上生物量及其与土壤水分关系的比较研究[J].草业学报,2011,20(3):20-27.
[15]苏嫄,焦菊英,马祥华.黄土丘陵沟壑区主要群落地上生物量季节变化及其与土壤水分的关系[J].水土保持研究,2012,19(6):7-12.
[16]张雷明,上官周平.黄土高原土壤水分与植被生产力的关系[J].干旱区研究,2002,19(4):59-63.
[17]王国杰,汪诗平,郝彦宾,等.水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响[J].生态学报,2005,25(7):1649-1656.
[18]白春利,阿拉塔,陈海军,等.氮素和水分添加对短花针茅荒漠草原植物群落特征的影响[J].中国草地学报,2013,35(2):69-75.
[19]董世魁,丁路明,徐敏云,等.放牧强度对高寒地区多年生混播禾草叶片特征及草地初级生产力的影响[J].中国农业科学,2004,37(1):136-142.
[20]王文,蒋文兰,谢忠奎,等.黄土丘陵地区唐古特白刺根际土壤水分与根系分布研究[J].草业学报,2013,22(1):20-28.
[21]傅伯杰,王军,马克明.黄土丘陵区土地利用对土壤水分的影响[J].中国科学基金,1999,4:225-227.
[22]胡良军,邵明安.黄土高原植被恢复的水分生态环境研究[J].应用生态学报,2002,13(8):1045-1048.
[23]武春华,陈云明,王国梁.黄土丘陵区典型群落特征及其与环境因子的关系[J].水土保持学报,2008,22(3):64-69.
[24]臧岳铭,朱志红,李英年,等.高寒矮嵩草草甸物种多样性与功能多样性对初级生产力的影响[J].生态学杂志,2009,28(6):999-1005.
[25]Zhou Z C,Shangguan Z P.Vertical distribution of fine roots in relation to soil factors inPinustabulaeformisCarr.forest of the Loess Plateau of China[J].Plant and Soil,2007,291:119-129.
[26]王正文,邢福,祝廷成,等.松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应[J].植物生态学报,2002,26(6):708-716.
[27]王俊,刘文兆,钟良平,等.长期连续种植苜蓿草地地上部分生物量与土壤水分的空间差异性[J].草业学报,2009,18(4):41-46.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
今日农业(2020年19期)2020-12-14
林业调查规划(2020年3期)2020-06-03
高原山地气象研究(2016年2期)2016-11-10
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
